
Abstract
The present study evaluated the effects of curcumin on epithelial cell apoptosis, the immunoreactivity of the phospho–c-Jun N-terminal kinase (JNK) and phospho-p38 mitogen-activated protein kinases (MAPKs) in inflamed colon mucosa, and oxidative stress in a rat model of ulcerative colitis induced by acetic acid. Rats were randomly divided into three groups: control, acetic acid, and acetic acid+curcumin. Curcumin (100 mg/kg per day, intragastrically) was administered 10 days before the induction of colitis and was continued for two additional days. Acetic acid–induced colitis caused a significant increase in the macroscopic and microscopic tissue ranking scores as well as an elevation in colonic myeloperoxidase (MPO) activity, malondialdehyde (MDA) levels, and the number of apoptotic epithelial cells in colon tissue compared to controls. In the rat colon, immunoreactivity of phospho–p38 MAPK was increased, whereas the phospho-JNK activity was decreased following the induction of colitis. Curcumin treatment was associated with amelioration of macroscopic and microscopic colitis sores, decreased MPO activity, and decreased MDA levels in acetic acid-induced colitis. Furthermore, oral curcumin supplementation clearly prevented programmed cell death and restored immunreactivity of MAPKs in the colons of colitic rats. The results of this study suggest that oral curcumin treatment decreases colon injury and is associated with decreased inflammatory reactions, lipid peroxidation, apoptotic cell death, and modulating p38- and JNK-MAPK pathways.
Introduction
I
A growing body of evidence indicates that abnormal production of reactive oxygen species (ROS) and altered antioxidant defenses participate in the intestinal damage observed in IBDs. 2 During inflammation, ROS formation and the activation of nuclear factor kappa-beta (NF-κB) have been shown to result in adaptive modifications of damaged cells, such as the expression of proinflammatory mediator genes and apoptosis-mediated cellular death. 3 In ulcerative colitis, the apoptotic rate is significantly increased and seems to be the main factor responsible for the loss of epithelial cells. 4 –6 Recent studies have demonstrated that an increase in the number of apoptotic epithelial cells might play a role in the disruption of epithelial defenses and in the acceleration of mucosal inflammation. 7 Mitogen-activated protein kinases (MAPKs), such as c-Jun N-terminal kinase (JNK) and p38, modulate the transcription of many genes involved in the inflammatory process. 8 However, the literature regarding the exact role of MAPKs in intestinal inflammation remains unclear.
Curcumin, a major component of turmeric, is known to have anti-inflammatory and antioxidant properties, 9 and can prevent and ameliorate the severity of the inflammatory damage in dextran sulfate sodium (DSS)–, dinitrobenzene sulfonic acid (DNB)–, and trinitrobenzene sulfonic acid (TNBS)–induced colitis by inhibiting NF-κB. 10 –12 However, there has only been one study in the literature supporting the antiapoptotic effect of curcumin in ulcerative colitis. 13 Therefore, we performed the present study to further characterize the roles of epithelial cell apoptosis, immunoreactivity of phospho-p38 and phospho-JNK in the inflammation of colonic mucosa and evaluate the possible protective effects of curcumin in a rat model of acetic acid–induced ulcerative colitis.
Materials and Methods
Animals
Twenty-one male Wistar albino rats (4 months old, weighing 300–350 g) were used in this study. The animals were bred and maintained under clean barrier conditions in the Experimental Animal Center of Trakya University. The rats were housed in wire-mesh cages with a maximum of four rats per cage. All animals were daily fed tap water and pellet foods, including 21% pure protein under optimum laboratory conditions (temperature, 22°C±2°C; humidity, 50%–55%; light/dark period: 12 h/12 h). All animals received humane care according to the criteria outlined in the “Guide for the Care and Use of Laboratory Animals” prepared by the National Academy of Sciences and published by the National Institutes of Health. The study was approved by the Ethics Committee of Trakya University (TÜHDYEK 2009/043).
Reagents
Curcumin was purchased from Sigma Chemical Co. Dimethylsulfoxide (DMSO) and acetic acid were obtained from Merck. All other chemicals and reagents used were of analytical grade.
Experimental design
Animals were randomly divided into three groups: the sham-control group (n=7) received a saline solution instead of acetic acid and without therapy; the acetic acid colitis group (n=7) received acetic acid enema without any therapy; and the acetic acid+curcumin group (n=7) received acetic acid enema and daily 100 mg/kg curcumin (in 1 mL of DMSO) using an intragastric tube. Curcumin was administered for 10 days before the induction of colitis and was continued for two additional days. In the present study, the dose and administration schedule of oral curcumin pretreatment was chosen on the basis of dose–response effects in a preclinical animal study. 14 The rats in the sham-control and acetic acid colitis groups were treated with 1 mL/kg DMSO (vehicle of curcumin) in the same way.
Colitis induction and sample collection
All animals were fasted overnight, with access to water ad libitum, before induction of colitis as previously described by Mustafa et al. 15 and were anesthetized with xylazine/ketamine (10/90 mg/kg, intraperitoneally). A soft 6F pediatric catheter inserted through the rectum into the colon to a distance of 8 cm was used to inject 2 mL (3%, v/v) acetic acid in 0.9% saline. The rats were then maintained in a supine Trendelenburg position for 30 sec to prevent early leakage of the intracolonic instillate. Control rats received 2 mL/rat saline solution enema (vehicle of acetic acid). All animals were sacrificed by cervical dislocation under xylazine/ketamine anesthesia 48 h after colitis induction. The abdomen was opened by a midline incision and colonic biopsies were taken for macroscopic scoring, histopathological examination, and biochemical studies. At the same time, epithelial cell apoptosis was detected by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay.
Clinical course assessment
The rats were observed on a daily basis and checked for diarrhea, loose stools, and rectal bloody stools. These observations were represented as a numerical score (Table 1). Body weights of all animals were recorded before and after the induction of colitis.
Macroscopic assessment
At postmortem laparotomy, 6 cm of colon extending proximally for 2 cm above the anal orifice was excised, freed from surrounding tissues, opened along the antimesenteric border, rinsed with a cold saline solution, and weighed. The inflamed colon was assessed according to the modified criteria for colonic changes, as reported in Table 1. 16 Each colon was scored by an independent observer according to a scale ranging from 0 (normal) to 15 (maximal activity of colitis) indicating ulcerations and severe inflammation of the colon.
Microscopic assessment
The segments of colon were collected from all animals, and then the distal colon segments were cut into small pieces of 0.5 cm. In each animal, two samples were selected randomly, fixed in 10% formalin solution, and embedded in paraffin. Serial 5 μm sections were cut and stained with hematoxylin–eosin, Masson's trichrome, and Periodic Acid Schiff using standard methods. The colonic pathological changes were graded semiquantitatively from 0 to 11 according to the described criteria: 17
• loss of mucosal architecture: 0, none; 1, mild; 2, moderate; 3, severe
• cellular infiltration: 0, none; 1, mild; 2, moderate; 3, severe
• muscle thickening: 0, none; 1, mild; 2, moderate; 3, severe
• crypt abscess formation: 0, negative; 1, positive
• goblet cell depletion: 0, negative; 1, positive
In situ TUNEL assay
Apoptotic cells in the colonic epithelial layer were detected with the TUNEL assay (ApopTaq Peroxidase In Situ Apoptosis Detection Kit; S7101-KIT) by following the manufacturer's instructions. The apoptotic cells were counted as follows: five high-power lens visual fields (each 200 cells) and a total of 1000 epithelial cells were counted. The positive cells were scored for apoptosis. Data were expressed as numbers of apoptotic cells per total number of cells counted. The areas of necrosis (e.g., in an ulcer bed) were excluded from the evaluation in the colitis group. For this reason, apoptotic assays were evaluated for four animals in this group.
Staining specificity controls
Normal female rodent mammary gland tissue sections were used as positive control. For negative control, distilled water was used instead of the Tdt enzyme.
Immunohistochemistry
Sections were incubated at 56°C overnight, then deparaffinized in xylene and rehydrated in a graded series of ethanol. They were boiled in citrate buffer (10 mM; pH 6.0) for 15 min for antigen retrieval. Following washing with Tris-buffered saline (TBS), the sections were immersed in 3% H2O2 (in 50% methanol/50% distilled water) for 5 min to inhibit endogenous peroxidase activity. The sections were washed in TBS 3×5 min, and then were incubated with 5% normal goat serum (Vector Laboratories) in TBS for 30 min at room temperature to block nonspecific binding of antibodies. After that, sections were incubated overnight at 4°C with the primary antibodies monoclonal rabbit phospho-JNK (Cell Signaling Technology), diluted 1:100, and monoclonal rabbit phospho-p38 (Cell Signaling Technology), diluted 1:300. The sections were rinsed with TBS, and then the biotinylated goat anti-rabbit antibody (Vector Laboratories) was added at a 1:400 dilution for 30 min at room temperature. Negative controls included stained in tissue sections without the primary antibody. After washes in TBS, the antigen–antibody complex was detected by using a streptavidin–biotin peroxidase kit (Vector Laboratories). Following washing with TBS, they were incubated with 3,3-diaminobenzidine (Vector Laboratories) to visualize immunolabeling, and finally counterstained with Mayer's hematoxylin. The intensity for phospho-JNK and phospho-p38 immunoreactivity was semiquantitatively evaluated using the following intensity categories: (−), no staining; (+), weak, but detectable staining; (++ ), moderate or distinct staining; (+++), intense staining. In each slide, five randomly selected areas were evaluated under a light microscope (× 40 magnification) at different times by two investigators blinded to the experimental condition. Their average score was used.
Myeloperoxidase activity assay
Myeloperoxidase (MPO) is a biochemical marker of neutrophil infiltration into intestinal tissues 16 and was used to confirm colitis. Tissue samples were homogenized in 10 volumes of ice-cold potassium phosphate buffer (50 mM K2HPO4, pH 6.0) containing hexadecyltrimethylammonium bromide (0.5%, g/mL). The MPO activity was assessed by measuring the H2O2-dependent oxidation of o-dianisidine.2HCl. One unit of the enzyme activity was defined as the amount of MPO present per gram of tissue weight that cause a change in absorbance of 10 min at 460 nm (37°C). 18
Lipid peroxidation determination
Malondialdehyde (MDA) is used as a marker of lipid peroxidation. MDA was estimated to indicate peroxide damage to membrane caused by inflammation. Lipid peroxidation in tissues was evaluated by the thiobarbituric acid method. The colon tissue samples were homogenized in 10% KCl (1:10 g/mL) in a glass–glass homogeneizer. Homogenate was added to the reaction mixture, which included 8.1% sodium dodecyl sulfate, 0.82% thiobarbituric acid, and acetate buffer (3 M, pH 3.5), and the reaction was monitorized at 532 nm by spectrophotometery. 19 Results were expressed as μmol/L using a standard curve.
Assessment of antioxidant status
The colon tissue samples (0.5 g) were homogenized with five volumes per weight, the 0.05 M phosphate buffer pH 7.0 containing 1% (g/mL) Triton X-100 in a glass–glass homogeneizer. The extract was centrifuged for 20 min at 10,000 g (+4°C). The supernatant was used in the assay for glutathione peroxidase (GPx), catalase (CAT), and superoxide dismutase (SOD). SOD and GPx activities were measured, respectively, by the Ransod kit (Randox Cat. No: SD 125) and Ransel kit (Randox Cat. No: RS 505) by spectrophotometry. One unit SOD activity was defined as the amount of enzyme that causes a 50% inhibition of the rate of reduction of 2-(4-iodophenyl)-3-(nitrophenol)-5-phenyltetrazolium chloride under the conditions of assay (at 37°C, pH 7.0). One unit GPx activity was defined as the amount of enzyme that oxidized 1 μmol NADPH to NADP in 1 min at 37°C. One unit CAT was defined as the amount of enzyme that decomposed 1 μmol H2O2 per minute at 30°C and pH 7.0. 20 The amount of total protein of colon was determined by using the method of Lowry et al. 21 Enzyme activities were expressed as units (U)/mg protein.
Statistical analysis
The Kolmogorov–Smirnov test was used to assess the normality of numeric variables. For the numeric variables that were normally distributed, comparison between three groups was made by analysis of variance. For the scores and non-normally distributed variables, comparison between two and three groups was made by the Mann–Whitney U and Kruskal–Wallis test. The appropriate post hoc test was selected for multiple (pairwise) comparisons. The results are expressed as mean±standard deviation and median (25–75 percentiles). Significance was defined as P<.05.
Results
Clinical findings
The clinical results of colitis induction are shown in Table 2. Colonic infusion of acetic acid leads to diarrhea and body weight loss in rats. Intracolonic administration of acetic acid caused a significant (P<.01) body weight loss in all groups. There was a marked weight loss during the study in the colitic rats compared to the control animals, which tended to gain weight (+7.1±3.6). Weight loss was the greatest in the acetic acid group (−16±9.8). However, curcumin-treated animals lost less weight (−3±3.9) compared with the untreated colitic rats (P<.05).
Values are expressed as mean±SD and median (25–75 percentiles), n=7.
P<.05, ** P<.01, *** P<.001 vs. control.
P<.05, †† P<.01, ††† P<.001 vs. colitis.
MPO, myeloperoxidase; ND, nondamage; SD, standard deviation.
Macroscopic findings
The acetic acid-induced colitis was characterized by a thick and stiff colon wall due to the presence of edema. The colonic mucosa morphologically appeared hyperemic and ulcerated. The dilated colon was markedly inflamed. Moreover, multiple adjacent organs adhesions were observed in some rats. Rectal acetic acid administration caused severe macroscopic inflammation in the colon when assessed by a macroscopic damage score (Table 2). In the acetic acid-induced group, the macroscopic score was significantly higher compared to the control group (10 [7–12] out of 15). On the other hand, oral curcumin treatment significantly prevented macroscopic damage except slight hyperemia (3 [2–3] out of 15, P<.01 vs. colitis).
Histopathological findings
In the control group, histological structures were observed to be normal without inflammatory changes in any layer (Fig. 1A–C). In the colitic groups, the acetic acid induced severe inflammation in the colon 48 h after rectal inoculation. Histological examination demonstrated the presence of focal mucosal ulceration extending through the muscularis mucosa. Moreover, there was evidence of mucosal desquamation with the loss of epithelial and goblet cells. The lamina propria mucosa showed edema, hemorrhages, fiber hyperplasia, and the infiltration of inflammatory cells. There was marked edema with hemorrhage in the submucosa, accompanied by inflammatory cell infiltration, all of which result in the thickening of the tunica submucosa. In addition, the tunica muscularis was found to be thicker compared with the control group, and the colon walls had became smooth (Fig. 1D–F). We also observed that oral curcumin treatment clearly prevented these abnormal histological findings of the acetic acid–induced colonic inflammation (Fig. 1G–I). As shown in Table 2, the microscopic score (9 [6–9]) of the colons in the acetic acid colitis group was significantly higher compared with the control group (P<.01 vs. control). Curcumin treatment markedly attenuated the extent and severity of colitis. As a result, the histopathologic score (3 [2–3]) in the curcumin-treated group was significantly decreased.
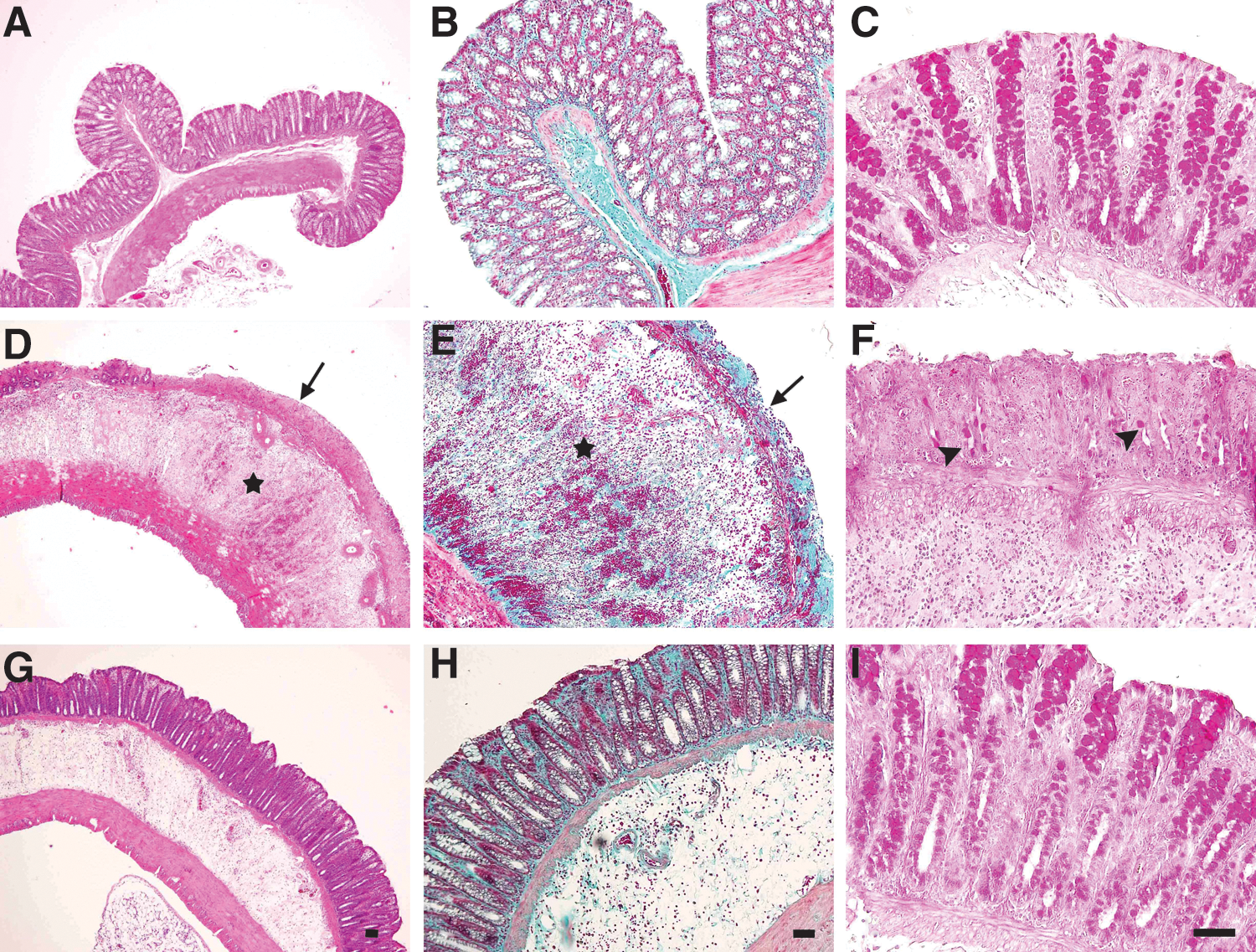
Representative micrographs of rat colon sections stained with hematoxylin–eosin
Evaluation of epithelial cell apoptosis
TUNEL-positive epithelial cells for each group are depicted in Figure 2. In the control group, the number of apoptotic epithelial cells was negligible (Fig. 2A). In the colitic groups, acetic acid administration led to a marked induction of cell apoptosis in the colons 48 h after rectal inoculation (Fig. 2B). In comparison with the control group (1 [1–2]), the number of apoptotic cells in the acetic acid-induced colitis group (5 [2.8–7.3]) was significantly increased (P<.01). Oral curcumin treatment clearly prevented programmed cell death in the colons of colitic rats (Fig. 2C), which lead to a significant decrease in the number of apoptotic epithelial cells (2 [1–3]), P<.05) in this group.
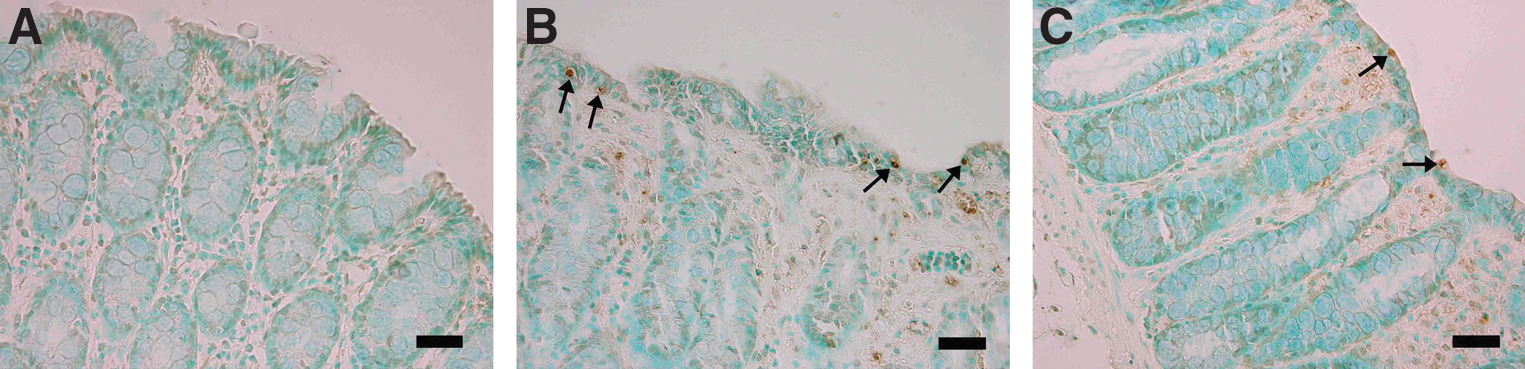
Detection of apoptotic cells in terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL)-stained sections of rat colon.
Immunohistochemical findings
The intensity of the phospho-JNK and the phospho-p38 immunolabeling in the epithelial cells of colon is summarized in Table 3. The colonic sections of the acetic acid colitis group had more phospho-p38 immunopositive staining compared to the control group (Fig. 3). However, the immunoreactivity for phospho-JNK was decreased in the colitic group (Fig. 4). Curcumin treatment ameliorated the activity of the above-mentioned MAPKs in colitic rats and in the same group, a moderate immunoreactivity for phospho-p38 and phospho-JNK was detected (Figs. 3 and 4).
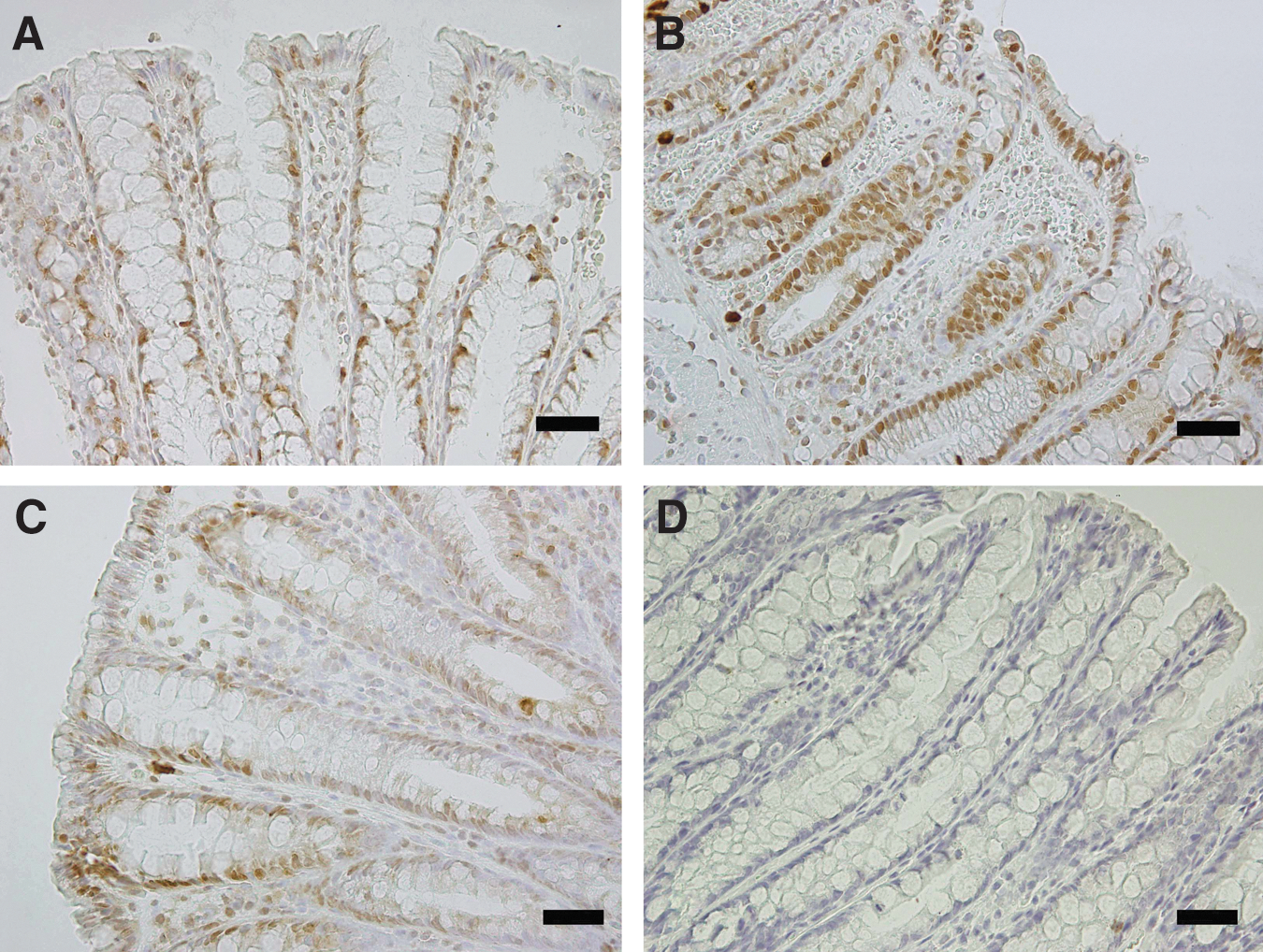
Effect of curcumin pretreatment on the phospho-p38 expression after rectal acetic acid administration as evaluated by immunohistochemistry. Photomicrographs of colonic sections depicting
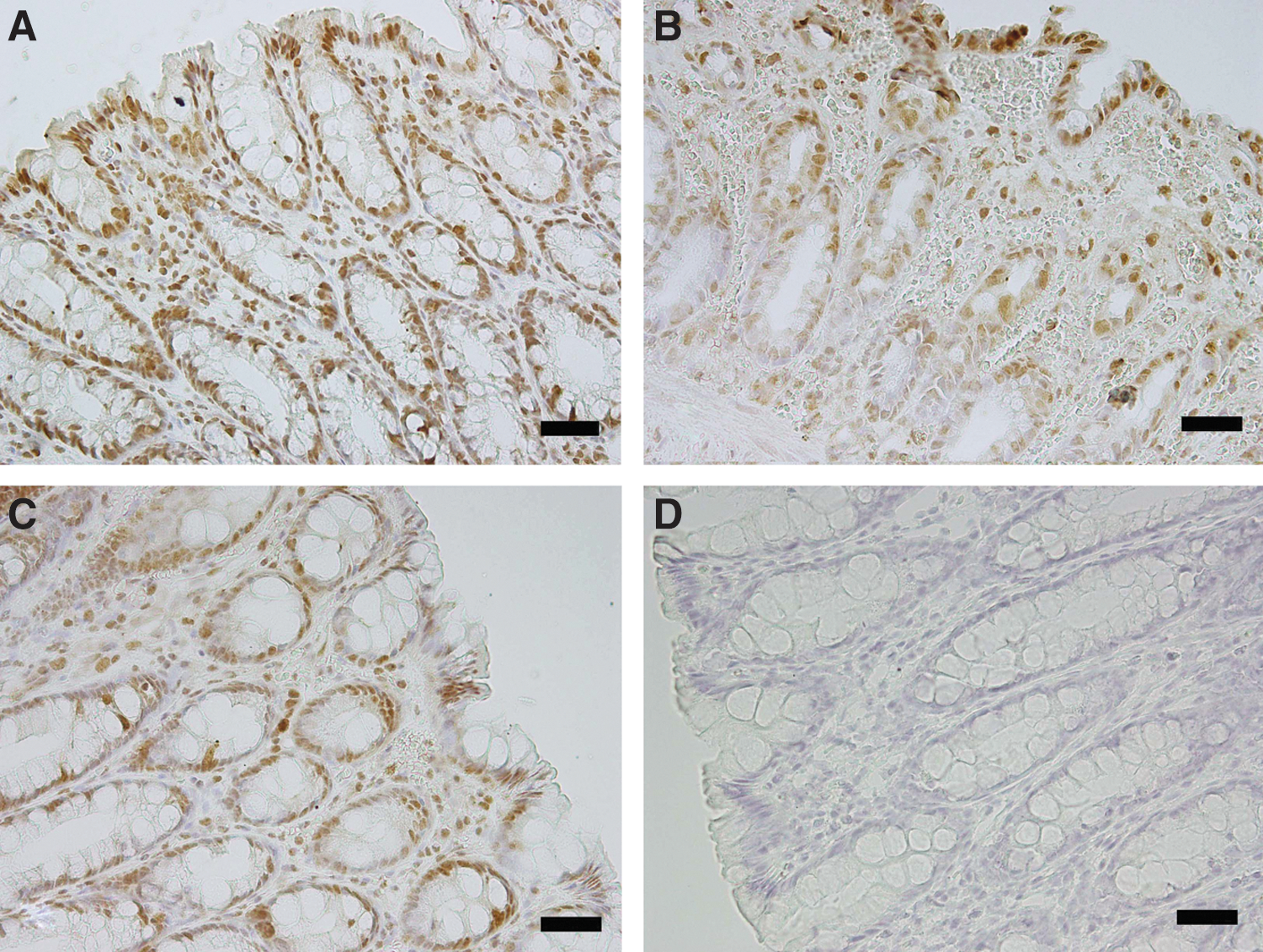
Effect of curcumin pretreatment on the phospho-c-Jun N-terminal kinase (JNK) expression after rectal acetic acid administration as evaluated by immunohistochemistry. Photomicrographs of colonic sections depicting
JNK, c-Jun N-terminal kinase.
The inflammatory response
Intracolonic acetic acid injection resulted in a colonic inflammatory reaction. The inflammatory status of the colon was characterized biochemically and morphologically by the colon wet weight and the MPO activity (Table 2). The colon wet weight was determined per 6 cm of colon length. The colon weight, a rough measure of edema and inflammation, was significantly increased in the colitic groups compared to the control group (P<.01), while oral curcumin supplementation prevented the increase of colon weights. The MPO activity in the colonic mucosa was also significantly elevated in the acetic acid-induced colitis group compared to the control group (P<.001), which was in aggrement with the histological findings. Nevertheless, the MPO levels were significantly reduced (P<.001 vs. colitis) in the curcumin-treated colitis rats.
Biochemical assessment
Table 4 presents the levels of lipid peroxidation and antioxidant status in the experimental groups. To examine oxidative stress, the MDA levels in the colon tissue were determined. The MDA levels in the acetic acid-treated rats were significantly higher compared with control animals. In contrast, oral curcumin treatment administered to the colitis rats significantly reduced the MDA levels compared to the untreated colitis group. Acetic acid administration also caused a significant increase in the colonic enzymatic (SOD and CAT) defense systems (P<.001). However, no difference was observed in the GPx activity in any of the groups. Curcumin treatment prevented the elevation of the CAT activity in colitic rats (P<.001), but the change of the SOD activity was not statistically significant compared to the untreated colitis group.
Values are expressed as mean±SD, n=7.
P<0.05, *** P<0.001 vs. control.
P<0.05, ††† P<0.001 vs. colitis.
MDA, malondialdehyde (μmol/L); SOD, superoxide dismutase (U/mg protein); CAT, catalase (U/mg protein); GPx, glutathione peroxidase (U/mg protein).
Discussion
In pathophysiological situations, the balance between cell proliferation and cell death is often altered, resulting in the loss of tissue homeostasis. Insufficient or excessive apoptosis may be a causative factor for the onset of various colon diseases, including ulcerative colitis, Crohn's disease, and cancer. 22,23 The findings of this study show that oxidative damage and apoptosis contribute to the development of colon injury in the acute model of ulcerative colitis (UC) induced by the colonic infusion with acetic acid, a reproducible and simple model that shares many features with human colitis. Furthermore, we documented that curcumin may counteract acetic acid-induced UC by decreasing the inflammatory responses and the level of oxidative stress and by utilizing other possible mechanisms such as the modulation of the epithelial cell apoptosis, the downregulation of p38 MAPK, and the upregulation of JNK pathways.
UC is a chronic IBD. Despite the fact that its etiology is still unknown, many factors have been implicated in UC pathogenesis, such as neutrophil infiltration and the overproduction of proinflammatory mediators, including cytokines, arachidonate metabolites, and reactive oxygen metabolites. These proinflammatory mediators ultimately lead to the disruption and the ulceration of the mucosa. 24,25 Morphological injury and histological changes are usually applied to evaluate the severity of the inflammatory injuries. MPO, a good indicator of the acute inflammatory response, reflects the activity of neutrophils. 16 In our study, the MPO activity was elevated in rats treated with acetic acid, which verifies the development of inflammation. This elevation in MPO activity was supported by the histological findings, which showed goblet cell depletion, hemorrhagic necrosis, mucosal ulcerations, and infiltration of inflammatory cells in the colons of the colitic rats, similar to the findings of a recent study. 26 We also observed that treatment with curcumin remarkably reduced the histological scores and the MPO activity, confirming the anti-inflammatory effect of curcumin. Our findings are well-matched with previous reports from other experimental models of ulcerative colitis. 11,14
Increased ROS production and impaired antioxidant defense mechanisms are considered to be causative factors in inflammatory diseases such as IBDs. 2 MDA, an end product of the lipid peroxidation process, is commonly known as a marker of oxidative stress. In our study, acetic acid instillation induced ROS formation, as verified by the elevated MDA levels, and increased the SOD and CAT activity, but it had no effect on the GPx activity in the rat colons. It has already been shown that the colonic MDA contents increased in both human 27 and experimental animal studies 26 of ulcerative colitis, in aggrement with the current findings. However, the results of studies evaluating the levels and the activity of antioxidant enzymes, for example, SOD and CAT, in UC are controversial. In contrast to our findings, El-Abhar et al. 26 reported a significant decrease in the SOD and CAT levels in colon tissues in acetic acid-induced colitis. Conversely, Beno et al. 28 also reported a clear elevation in the SOD and CAT activities in IBDs, similar to our study. Furthermore, we observed that treatment with curcumin significantly decreased the elevation of MDA and CAT levels in the colons of colitic rats. This protective effect may be attributed to the ability of curcumin to reduce neutrophil infiltration (as indicated not only by the macroscopic and the histological data, but also the MPO activity) in the inflamed colonic tissue. Curcumin has been demonstrated to counteract in DSS-, DNB-, and TNBS-induced colitis by activating inflammatory modulators in a similar manner to that shown by the results of this study. 11,12,14
Under normal conditions, epithelial cell shedding does not mediate an alteration of the intestinal barrier. 29 However, an increase in the apoptotic rate of the epithelial cell in the colon mucosa may generate leaks contributing to a decrease in the barrier function and thus cause gross lesions such as erosions or ulcers. It has been shown that the loss of ions and water in the intestinal lumen causes leak flux diarrhea due to the epithelial apoptosis. 30 Recent studies reported an increased number of apoptotic epithelial cells in the experimental models of UC induced by DSS, DNB, TNBS, and tumor necrosis factor (TNF)-α. 6,7,13,31 To investigate whether an acute model of UC induced by acetic acid is associated with apoptotic cell death, we performed a TUNEL assay on the inflammed colon tissues. Forty-eight hours after the rectal administration of acetic acid, colon tissues exhibited a marked appearance consistent with apoptotic epithelial cells. In addition, we observed diarrhea and body weight loss in the colitic animals. To date, there has been no evidence indicating an increase in epithelial cell apoptosis in the acetic acid–induced colitis model. In the present study, we show for the first time that epithelial cell apoptosis increased in this model of experimental colitis.
Curcumin has a wide range of biological activities such as anti-inflammatory, antitumorigenic, and antioxidant activities. It is also under preclinical evaluation as an anti-inflammatory and anticancer drug. 32 In addition, it was recently shown that curcumin acts as an important antiapoptotic mediator in several cellular events. 33 –35 It is well-established that cellular signaling pathways involving the Bcl-2/Bax family of proto-oncogenes, the transcription factor NF-κB, TNF-α, and interleukin-1β are able to induce apoptosis. 34,36 Molecular studies have shown that the anti-inflammatory effects of curcumin in the treatment of colitis result from the inhibition of the NF-κB pathways and its already-known antioxidant properties. 11,12,24 However, only one study has shown the relationship between curcumin and epithelial cell apoptosis in ulcerative colitis. The same study reported that curcumin attenuated ROS-related cytotoxicity, the levels of oxidants, and colon epithelial apoptosis by inhibiting the priming of neutrophils and macrophages in an immune-mediated model of TNF-α-induced colitis. 13 Similarly, our results showed that curcumin markedly reduced epithelial cell apoptosis and the severity of diarrhea in an acetic acid-induced colitis model.
It is known that ROS can activate a variety of downstream signaling pathways such as MAPKs, which can activate transcription factors modulating a number of different steps in the inflammatory cascade. It has been recently reported that the JNK and p38 MAPK activities are elevated in IBD patients. 37 –39 However, results of the experimental studies regarding the exact role of MAPKs in UC are highly controversial. Malamut et al. 40 reported that the p38 and JNK expression or activity did not increase following TNBS-induced colitis. Conversely, Hommes et al. 41 showed an enhancement of the phospho-p38 and the phospho-JNK expression in patients with active Crohn's disease. Furthermore, it has been reported that the JNK signaling pathway is activated in the DSS- and TNBS-induced colitis models and is modified by specific inhibitors such as SP600125, XG-102. 38,42 It has also been demonstrated that the colonic activity of p38 MAPK increases in experimental models of DSS-, TNBS-, and DNB-induced colitis. 43 –45 Similarly, with these studies, we also observed an increase in the phospho-p38 immunoreactivity in the colon epithelial cells of rats with colitis. Interestingly, our results showed a decrease in phospho-JNK immunoreactivity in the rat colon epithelial cells following acetic acid–induced colitis. Moreover, oxidative stress activates phosphorylation of members of the MAPK family such as the extracellular signal-regulated kinase (ERK), JNK, and p38. 46 In general, ERK delivers a survival signal, while JNK and p38 are associated with the induction of apoptosis under stressful conditions. 47,48 However, the activation of JNK has also been associated with the proliferation and the survival of cells. 49 Several studies have reported a prosurvival role for JNK. El-Najjar et al. 50 demonstrated a survival role for JNK and likely ERK activation in thymoquinone-induced apoptosis in colon cancer. Likewise, Dougherty et al. 51 demonstrated that JNK has a prosurvival role in neonatal cardiac myocytes when exposed to a hypoxic injury. Taken together, we suppose that increasing the phospho-p38 activity and decreasing the phospho-JNK activity may be associated with an excess number of apoptotic epithelial cells in the colons of rats with colitis. Furthermore, in our study, treatment with oral curcumin reduced the phospho-p38 immunoreactivity and enhanced the phospho-JNK immunoreactivity, confirming the antiapoptotic prosurvival effect of curcumin. Similar to our results, Salh et al. 44 showed that curcumin is able to attenuate DNB-induced colitis by inhibiting the activation of NF-κB and by decreasing the activity of p38 MAPK. In addition, Camacho-Barquero et al. 43 observed that curcumin treatment decreased the activation of p38 MAPK without affecting JNK activation, following TNBS instillation and suggesting that inhibition of the p38 MAPK signaling by curcumin could explain the decreased immunosignals of cyclo-oxygenase-2 and inducible nitric oxide synthase as well as the attenuated nitrite production in the colon mucosa. Furthermore, it was recently reported that curcumin suppresses p38 MAPK activation in IBD patients. 52
In conclusion, we demonstrated that oral administration of curcumin produced beneficial effects in acetic acid–induced colitis by decreasing the inflammatory responses and levels of oxidative stress. In addition, curcumin can act by other mechanisms such as the modulation of the epithelial cell apoptosis, the downregulation of p38 MAPK, and the upregulation of JNK pathways. Further preclinical studies may be helpful to characterize the antiapoptotic effect of curcumin in ulcerative colitis.
Footnotes
Acknowledgment
The authors are grateful to the Trakya University Research Center for financial support of this study (Project no: 2009/154).
Author Disclosure Statement
No competing financial interests exist.