
Abstract
Antrodia camphorata (AC) has been used as a traditional medicine to treat food and drug intoxication, diarrhea, abdominal pain, hypertension, pruritis (skin itch), and liver cancer in East Asia. In this study, we investigated anticancer activities of AC grown on germinated brown rice (CBR) in HT-29 human colon cancer cells. We found that the inhibitory efficacy of CBR 80% ethanol (EtOH) extract on HT-29 and CT-26 cell proliferation was more effective than ordinary AC EtOH 80% extract. Next, 80% EtOH extract of CBR was further separated into four fractions; hexane, ethyl acetate (EtOAc), butanol (BuOH), and water. Among them, CBR EtOAc fraction showed the strongest inhibitory activity against HT-29 cell proliferation. Therefore, CBR EtOAc fraction was chosen for further studies. Annexin V-fluorescein isothiocyanate staining data indicated that CBR EtOAc fraction induced apoptosis. Induction of G0/G1 cell cycle arrest on human colon carcinoma cell was observed in CBR EtOAc fraction-treated cells. We found that CBR decreased the level of proteins involved in G0/G1 cell cycle arrest and apoptosis. CBR EtOAc fraction inhibited the β-catenin signaling pathway, supporting its suppressive activity on the level of cyclin D1. High performance liquid chromatography analysis data indicated that CBR EtOAc fraction contained adenosine. This is the first investigation that CBR has a greater potential as a novel chemopreventive agent than AC against colon cancer. These data suggest that CBR might be useful as a chemopreventive agent against colorectal cancer.
Introduction
C
Many studies have reported that cell cycle inhibitors and apoptosis inducers are effective in reducing the risk for colon cancer. Thus, a strategy in the search of anticancer drugs is to screen agents that are capable of targeting apoptosis related molecules, including caspases and Bcl-2 family proteins, and cell cycle arrest markers. 4
It is well accepted that Wnt signaling is aberrant in >90% of human colorectal cancers. 5 β-Catenin is a key molecule that leads to colon tumor development. Many anticancer agents can reduce the nuclear accumulation of β-catenin, which results in the transcription of Wnt/β-catenin target genes, such as myc, cyclin D1, vascular endothelial growth factor, and matrix metalloproteinases (MMPs). 6 For instance, Wnt signaling activation can occur through phosphorylation of the negative regulatory elements glycogen synthase kinase 3 beta (GSK-3β), which in turn causes β-catenin protein stabilization. 7 Recent evidence demonstrated that mutations of components in the Wnt/β-catenin pathway generally occur in the early stage of colon cancer progression. Accumulation of β-catenin in the nucleus is related with late stages of tumor progression and the metastasis development. 8,9 Hence, several molecules in the Wnt/β-catenin pathway have been identified as novel targets for cancer therapy.
Recently, a number of medicinal mushrooms and herbal medicines have recently been spotlighted as chemopreventive agents due to their potential chemopreventive and therapeutic activities at non-toxic levels. 10 Antrodia camphorata (AC), also named Antrodia cinnamomea, is a parasitic fungus that only grows inside the heartwood wall of Cinnamomum kanehirai Hay (Lauraceae). AC has been used as a traditional medicine to treat food and drug intoxication, diarrhea, abdominal pain, hypertension, pruritis (skin itch), and liver cancer in East Asia. 7,11 –30 Recently, it was reported that AC induces apoptosis in ovarian cancer cells. 7 Another group demonstrated that AC extracts may be used as an adjuvant anticancer agent for human liver cancer. 11 However, mass production of AC in nature is difficult because it only inhabits broad-leaved forests at an altitude of 200–2000 m in Taiwan. It is not accessible to the public because of its scarcity and high production cost. To solve the high production cost and low production yield of AC, our group developed culture methods by inoculating AC on germinated brown rice (CBR) known to be rich in nutrients and biologically active compounds. The anticancer activities of CBR and its mechanism of action have yet to be determined in detail. The objective of this study was to investigate the anticancer activities of CBR on HT-29 human colon cancer cells.
Materials and Methods
Reagents and chemicals
AC grown on germinated brown rice (CBR) (Kucari 1102) and AC (Kucari 1101) were obtained from The Cell Activation Research Institute (Seoul, Korea). Minimum essential medium (MEM), RPMI 1640 medium, fetal bovine serum (FBS), penicillin-streptomycin, and trypsin-ethylenediaminetetraacetic acid (EDTA) were obtained from Gibco BRL (Grand Island, NY, USA). Propidium iodide (PI), NP-40, and RNase A were obtained from Sigma Chemical Co. (St. Louis, MO, USA). Anti–cyclin-dependent protein kinase 4 (anti-CDK4; Cell Signaling Technology Inc., Danvers, MA, USA), anti–cyclin D1 (Cell Signaling Technology), anti-CDK2 (Cell Signaling Technology), anti–cyclin E1 (HE12; Cell Signaling Technology), anti-Bcl2 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-Bax (Cell Signaling Technology), anti–caspase-9 (C9; Cell Signaling Technology), anti–cytochrome C (Cell Signaling Technology), anti–β-catenin (Cell Signaling Technology), phospho-GSK-3β (serine 9; Cell Signaling Technology), anti–β-actin, and horseradish peroxidase (HRP)–conjugated anti–rabbit and anti–mouse IgG antibodies (Santa Cruz Biotechnology) were obtained from the suppliers, respectively. The chemiluminescence detection kits were purchased from Biosesang (Seoul, Korea) and EZ-CyTox assay kit from Daelillab Service Co. (Korea).
Preparation of CBR extract
CBR was cultured using a similar method as previously described. 31 Authenticated voucher specimens of AC (Kucari 1101) and CBR (Kucari 1102) were deposited in the Herbarium at the College of Bioscience and Biotechnology, Konkuk University (Seoul, Korea). AC mycelia (Kucari 1101) was inoculated on germinated brown rice and cultured at 20–25°C for 4 weeks. The powdered material (1 kg) was extracted under reflux with 80% ethanol (EtOH). The total extract (178 g, yield [w/w], 17.8%) was dissolved with water. After removing the insoluble solid particles by filtration, the liquid phase was sequentially extracted by solvents with increasing polarity (hexane, ethyl acetate [EtOAc], butanol [BuOH], and water; 1:10 [w/v] for all solvents) to yield four fractions. The fractions were concentrated to dryness by a rotary evaporator. Thus, we obtained the following fractions: hexane fraction (16 g, yield [w/w] 1.6%), EtOAc fraction (4.5 g, yield [w/w] 0.45%), BuOH fraction (8.25 g, yield [w/w] 0.825%), and water fraction (10.86 g, yield [w/w] 1.086%; Fig. 1).
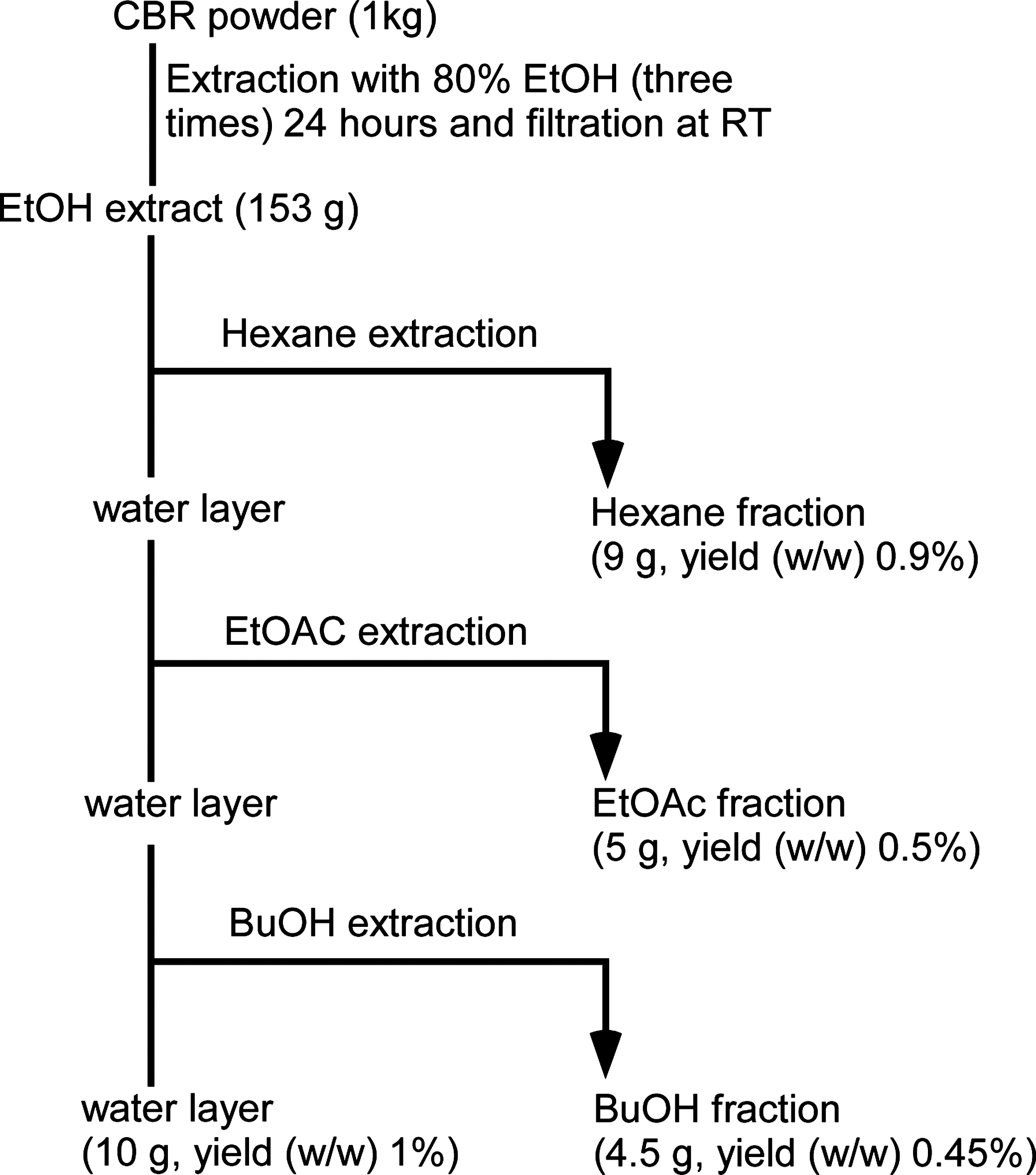
Scheme depicting the methodology used to obtain hexane, EtOAc, BuOH, and water fractions. EtOAc, ethyl acetate; BuOH, butanol.
Cell culture
HT-29 cells (human colon cancer cells) and CT-26 cells (murine colon cancer cells) were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were cultured in RPMI 1640 medium and MEM medium (Gibco BRL) supplemented with 10% heat-inactivated FBS (Gibco BRL), 100 U/mL of penicillin, and 100 μg/mL streptomycin at 37°C in a humidified incubator with 5% CO2.
Cell proliferation assay
The effect of 80% EtOH extracts of CBR, AC, and each CBR fraction (hexane, BuOH, EtOAc, and water) on HT-29 and CT-26 cell proliferation was measured using the EZ-CyTox kit (Daelillab Service Co.). The assay was done as the manufacturer's protocol. HT-29 or CT-26 colon cancer cells (1×104 cells/well) were plated on a 96-well plate and incubated in the presence or absence of 80% EtOH extracts of CBR and AC or CBR fractions (hexane, BuOH, EtoAc, and water) at concentrations of 0, 50, 100, 200, and 400 μg/mL for 48 and 72 h, respectively. A fixed amount (10 μL) of EZ-CyTox reagent was added to each well and incubated for an additional 1–2 h at 37°C. Cell proliferation levels were detected at an optical density of 450 nm, using an enzyme-linked immunosorbent assay Multi-Detection Reader (Tecan, Mannedorf, Switzerland).
Flow cytometric cell cycle analysis
Cell cycle progression was determined using flow cytometry analysis, as described previously. 31 HT-29 cells (3×105 cells/well) were plated on six-well plates in the presence or absence of CBR EtOAc fraction for 48 and 72 h. Then, cells were harvested by trypsinization, washed twice with phosphate buffered saline (PBS), and fixed with 70% ice-cold EtOH. After centrifugation, cells were stained with a staining solution 0.2% NP-40, RNase A (30 μg/mL), and PI (50 μg/mL; Sigma Chemical Co.). Cellular DNA contents were analyzed by flow cytometry, using a Becton-Dickinson laser-based flow cytometer (Becton-Dickinson, Franklin Lakes, NJ, USA). At least 10,000 cells were used for each analysis, and the results were displayed as histograms. The average percentage of cells in each phase of the cell cycle was determined over three independent experiments.
Determination of the apoptotic cell population by Annexin V-fluorescein isothiocyanate staining
HT-29 cells (3×105 cells/well) were plated on six-well plates for 54 h, and then replaced with fresh media in the presence or absence of CBR EtOAc fractions (0, 50, 100, and 200 μg/mL). Apoptotic cell death was identified by double staining with FITC-conjugated Annexin V and PI, using the Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kit (TREVIGEN®, Inc., Gaithersburg, MD, USA), according to the manufacturer's instructions. Data analysis was performed in a Becton-Dickinson FACSCalibur flow cytometry using CellQuest software (Becton-Dickinson).
Preparation of mitochondrial and cytosolic fractions
Cells were resuspended in cytosolic extraction buffer (250 mM sucrose, 70 mM potassium chloride, 137 mM sodium chloride [NaCl], 4.3 mM disodium phosphate, 100 mM dithiothreitol, and 200 μg/mL digitonin). After 20 min of incubation on ice, cell lysates were centrifuged. Supernatants contained cytoplasmic fractions and mitochondria were isolated from the pellet with mitochondria extraction buffer (50 mM Tris [pH 7.4], 150 mM NaCl, 2 mM EDTA, 0.2% Triton x-100, and 0.3% NP-40).
Immunoblot analysis
Immunoblotting analysis was performed as described previously. 32,33 HT-29 cells were treated with CBR EtOAc fractions at various concentrations (0, 50, 100, and 200 μg/mL) for 24 h. Twenty micrograms aliquots of protein were subjected to electrophoresis on 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis polyacrylamide gels and electrophoretically transferred onto polyvinylidene fluoride membranes (Bio-Rad Laboratories, Berkeley, CA, USA). The membranes were incubated in 5% skim milk solution, and then with antibodies against β-catenin, phospho-GSK-3β, Cdk4, cyclin D1, Bcl2, caspase-9 (1:2000), and β-actin (1:3000). The membranes were washed thrice in a 1×PBS-T buffer and incubated with HRP-conjugated secondary antibodies (1:5000) for 1–2 h. The membranes were washed and detection of the immunoreactive bands was performed using the enhanced chemiluminescence Western blotting detection system Biosesang.
Immunofluorescence microscopy
Cells were fixed with 10% neutral buffered formalin in PBS for 15 min and permeabilized with 0.2% Triton X-100 for another 10 min at room temperature. The cells were stained with anti-β-catenin antibody (Cell Signaling Technology), Alexa Fluor 594 conjugated anti-rabbit IgG (Cell Signaling Technology), and 4′,6-diamidino-2-phenylindole (Cell Signaling Technology). The cells were viewed using an inverted fluorescence microscope (200×objective) and processed using Axiovision image analysis 4.7 software (Carl Zeiss Vision GmbH, Munich, Germany).
High performance liquid chromatography analysis
Compounds in the extracts were analyzed by high performance liquid chromatography (HPLC) using an Agilent 1260 Infinity HPLC system (Santa Clara, CA, USA) with the reversed phase column (Luna C18, 250 mm×4.6 mm, 5 μm diameter; Phenomenex, Torrance, CA, USA). The flow rate and injection volume were 1.2 mL/min and 5–20 μL, respectively. The peaks were detected at 260 nm and collected at 30°C. Adenosine was purchased from Sigma-Aldrich (St. Louis, MO; 99%) and used without further purification. One milligram of adenosine was dissolved in 1 mL of 50% EtOH and filtrated using 0.45 μm membrane filters. While the mobile phase for the EtOH extract of CBR was 8% aqueous methanol, for the fractions separated based on the hydrophobicity, it was 6% aqueous methanol containing 0.1% monopotassium phosphate.
Statistical analysis
Values are presented as percentage±standard deviation of control. Student's t-test or one-way analysis of variance/Dunnett's t-test was used to analyze the statistical significance between the CBR-treated and control groups. Statistical analysis was performed using SPSS, version 12 (SPSS Inc., Chicago, IL, USA).
Results
Antiproliferative activity of AC grown on germinated brown rice (CBR) in HT-29 cell lines
Antiproliferative activity of 80% EtOH extracts of CBR and AC against colon cancer cell lines, HT-29 cells, and CT-26 cells, was determined. The half-maximal inhibitory concentration (IC50) of CBR 80% EtOH extract (IC50, 256.78±7.15 μg/mL) was significantly lower compared with AC 80% EtOH extract (IC50, 467.31±44.77 μg/mL) in HT-29 cells (Table 1 and Fig. 2A). Similarly, inhibitory effects on CT-26 cell growth in response to CBR 80% EtOH extract (IC50, 538.57±145.14 μg/mL) was stronger than AC 80% EtOH extract (IC50, 585.06±192.21 μg/mL; Table 1 and Fig. 2B). This suggests that the inhibitory efficacy of CBR 80% EtOH extract on colon cancer cell proliferation is more effective than ordinary AC EtOH 80% extract. Next, CBR was further separated into four fractions; hexane, EtOAc, BuOH, and water (Fig. 1), which were tested against HT-29 cell proliferation. CBR EtOAc fraction showed pronounced activity against HT-29 cells among these fractions (P<.01; Fig. 3A). Therefore, CBR EtOAc fraction was chosen for further studies. HT-29 cells were treated with CBR EtOAc fraction (0, 1, 10, 100, and 500 μg/mL) for 48 and 72 h. CBR EtOAc fraction significantly inhibited HT-29 cell proliferation in a dose- and a time-dependent manner (Fig. 3B). However, CBR EtOAc fraction did not affect mouse macrophage RAW264.7 cell viability (data not shown).

Inhibitory effect of 80% EtOH extracts of CBR and AC on colon cancer cell growth.

Time- and dose-dependent inhibitory activity of CBR fractions on HT-29 human colon cancer cell growth.
AC, Antrodia camphorata; CBR, Antrodia camphorata grown on germinated brown rice; IC50, 50% inhibitory concentration for cell viability; EtOH, ethanol.
Modulation of CBR EtOAc fraction on cell cycle progression
Based on the antiproliferative activity of CBR EtOAc fraction on HT-29 cells, we explored the potential antiproliferative mechanism, using cell cycle analysis. Cell cycle analysis data showed that CBR EtOAc fraction induced HT-29 cell arrest at G0/G1 phase of cell cycle (P<.01 vs. control). The rates in G0/G1 phase at the concentration of 0, 50, 100, 200, and 400 μg/mL were 53.77±3.97%, 56.15±4.63%, 59.15±2.98%, 69.85±3.42%, and 66.25±1.54%, respectively. The number of S and G2/M phase cells significantly decreased at 400 μg/mL of CBR extract (Fig. 4A). The rate of sub-G1 population at the concentration of 0, 50, 100, and 200 μg/mL was 0.55±0.06%, 4.42±0.01%, 9.97±0.18%, 16.68±1.06%, respectively (Fig. 4B).
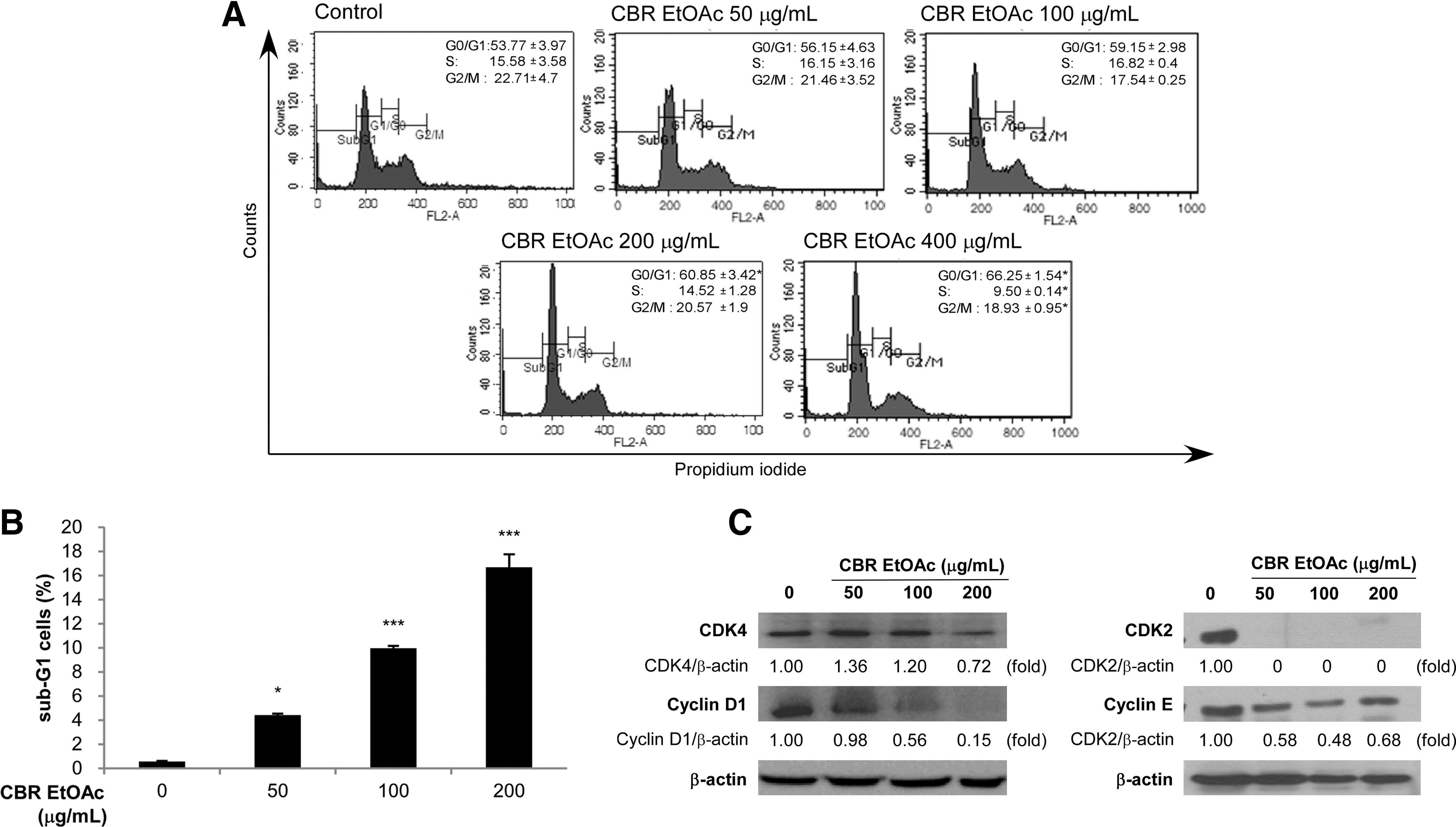
Effect of CBR EtOAc fraction on cell cycle progression. Cells were incubated in the presence or absence of CBR EtOAc fraction for the indicated time period.
Many cancer cells overexpress cyclins and CDKs, resulting in dysregulated and unchecked cell proliferation. 34 We investigated whether CBR EtOAc fraction could modulate the expression of CDKs and cyclins, involved in G0/G1 cell cycle arrest. Recent studies demonstrated that CDKs and cyclins were decreased in human cancer cells arrested at G0/G1 phase by anticancer drug treatment. 35 Marked decline in CDK4 and cyclin D1 occurred upon exposure of CBR EtOAc fraction to HT-29 cells (Fig. 4C). Reduced CDK2 and cyclin E protein levels were observed in CBR EtOAc fraction-treated HT-29 cells in a dose-dependent manner (Fig. 4C). These data indicated that CBR EtOAc fraction treatment caused G0/G1 arrest.
Effect of CBR EtOAc fraction on apoptosis in HT-29 cells
The degree of cellular apoptosis was evaluated by annexin V-FITC/PI double staining assay, which determined the percentage of cells in early (annexin V-positive cells) and late apoptosis (annexin V- and PI-positive cells). A dose-dependent increase in the numbers of apoptotic cells was observed after 54 h treatment. In the control group, 0.28+1.66% (early apoptosis+late apoptosis) were positive for annexin V-FITC staining, while CBR EtOAc fraction treatment resulted in 1.14+9.59%, 0.31+62.00%, and 0.17+92.04% at 50, 100, and 200 μg/mL, respectively (Fig. 5A). Apoptosis is regulated by apoptosis signaling molecules, including caspases, Bcl-2 protein family members, and cytochrome C. 36 Treatment of tumor cells with CBR extract resulted in an increase in the expression of Bax and repression of Bcl-2. Release of cytochrome c from mitochondria to the cytosol is essential in the mitochondrial pathway of apoptosis. In response to CBR EtOAc, the amount of released cytochrome c (cytosol) was increased while the level of cytochrome c in mitochondria was decreased. The decrease in the expression of procaspases-9 was also observed in CBR EtOAc–treated HT29 cells (Fig. 5B).

CBR EtOAc fraction-induced apoptosis in HT-29 human colon cancer cells.
CBR EtOAc fraction decreased the level of β-catenin protein and phosphorylated GSK-3β (inactive form) expression
Tumorigenesis is accompanied by the overexpression of β-catenin, an oncogenic protein. The accumulated nuclear β-catenin stimulates the transcription of many oncogenes, such as cyclin D137,38 and Myc. 39 To evaluate the effect of CBR EtOAc fraction on the level of β-catenin protein, HT-29 cells were treated CBR EtOAc fraction. As seen in Figure 6, CBR EtOAc fraction markedly decreased the level of β-catenin proteins and its nuclear translocalization in HT-29 cells. In concert with Western blot analysis, CBR treatment attenuated the intensity of β-catenin stained HT-29 cells. We next examined the possible mechanism for the decline in β-catenin protein levels by CBR EtOAc fraction. β-Catenin is one of the most well-known substrates of GSK-3β. Inhibition of GSK-3β through phosphorylation by AKT led to cytosolic β-catenin accumulation; thus, increasing its translocalization into the nucleus. 37,38 Further, CBR EtOAc fraction significantly suppressed phosphorylated GSK-3β (an inactive form) in HT-29 cells in a dose-dependent manner (Fig. 6). Based on these data, the inhibitory effect of CBR EtOAc fraction on HT-29 cell growth might be due to the suppression of β-catenin signaling pathway.
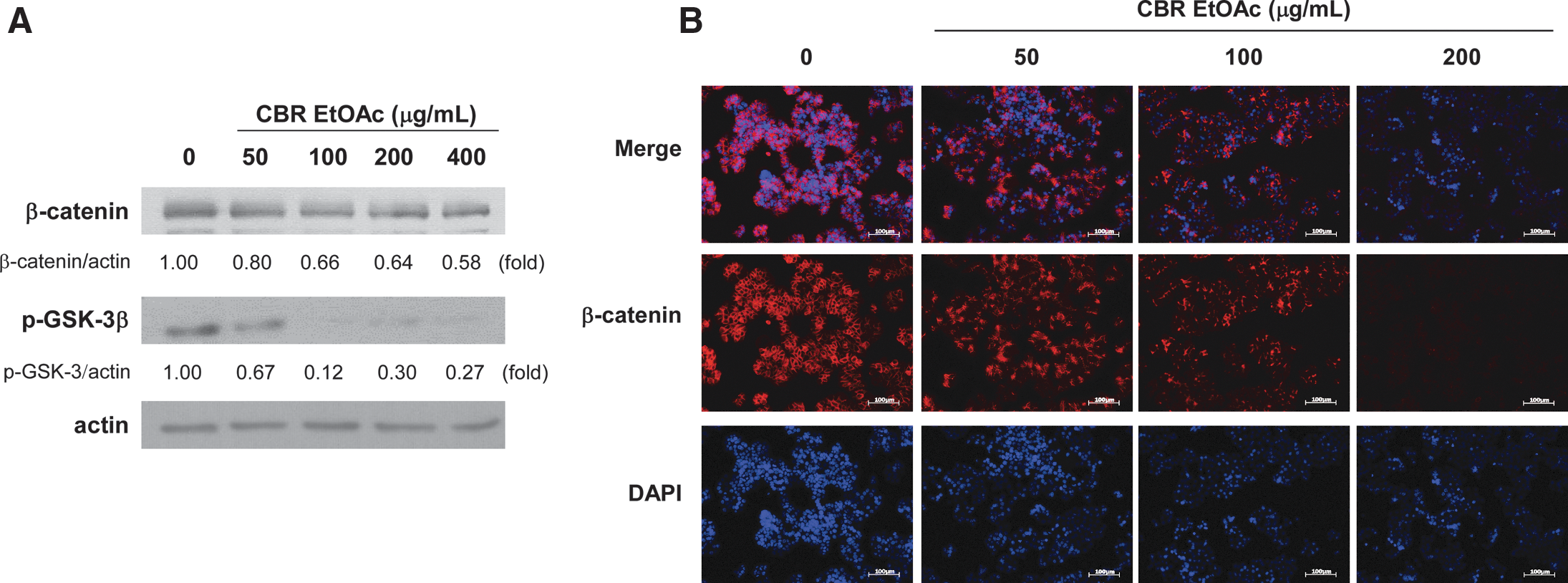
Effect of CBR EtOAc fraction on the β-catenin signaling in HT-29 cells. HT-29 cells were incubated in the absence or presence of CBR EtOAc fraction (0, 50, 100, and 200 μg/mL) for 48 h.
Chromatogram of adenosine of extracts of CBR
HPLC assay found that in the EtOH extract of CBR and AC, the authentic adenosine was observed at the retention time of 16 min. When the concentration of the EtOH extract of CBR was 36.29 mg/mL that of adenosine was determined to be 0.015% (Fig. 7). However, adenosine was not detected in AC extract. In the HPLC experimental condition for the fractions separated based on the hydrophobicity, the authentic adenosine was observed at the retention time of 22 min. The concentrations for n-hexane, EtOAc, BuOH, and water fractions were prepared as 29.6, 14.4, 34.6, and 120 mg/mL, respectively. Adenosine was observed in the EtOAc fraction only and its concentration was 0.0196% (Fig. 8). To investigate the observed strong antiproliferative effects of CBR EtOAc fraction among fractions, the apoptosis-inducing effect of adenosine (5 mM) was determined in HT-29 cells. 6.29% and 5.94% of the cells underwent early and late apoptotic death by adenosine treatment for 54 h, respectively.
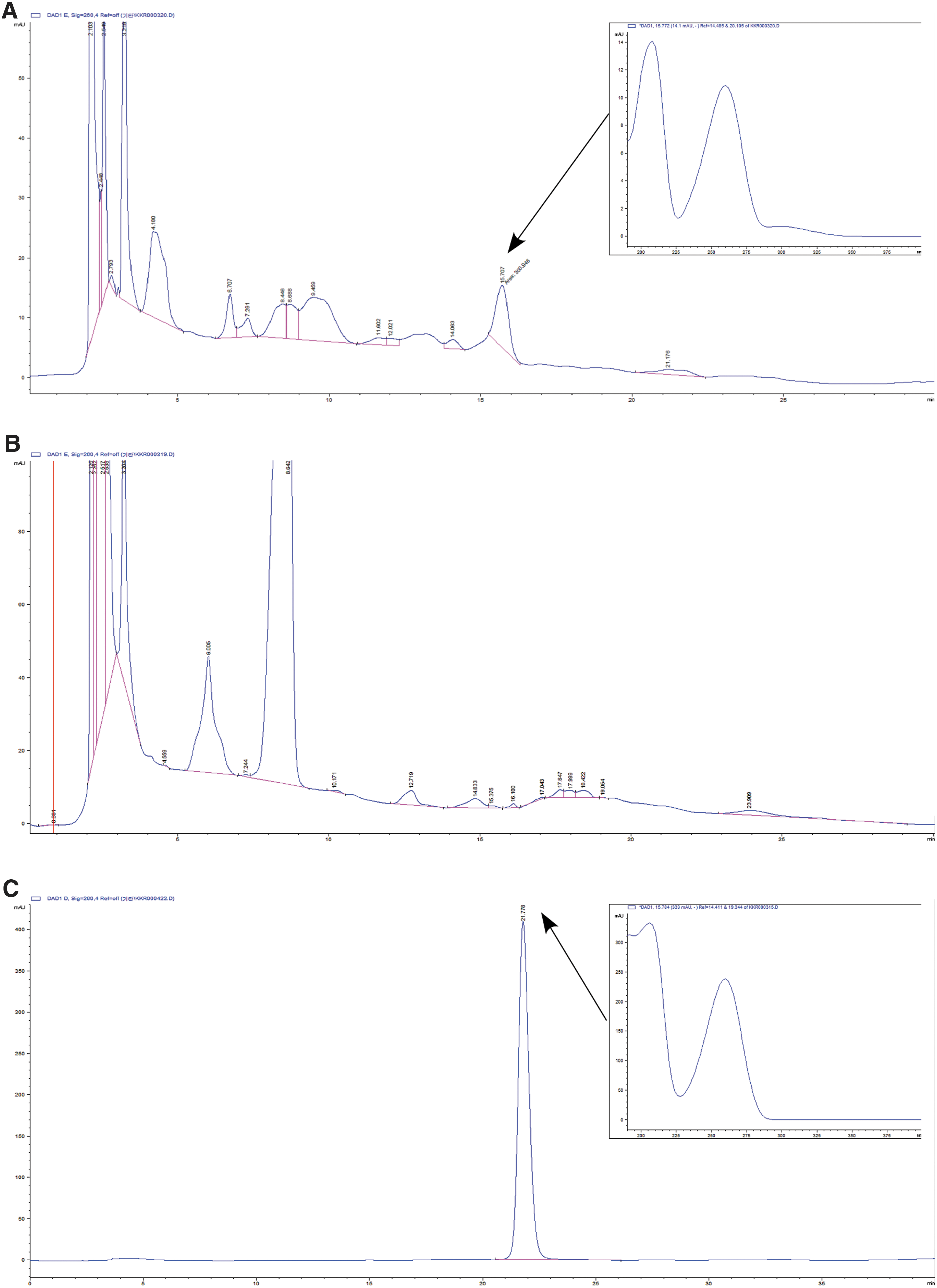
High performance liquid chromatography chromatograms of ethanol extract
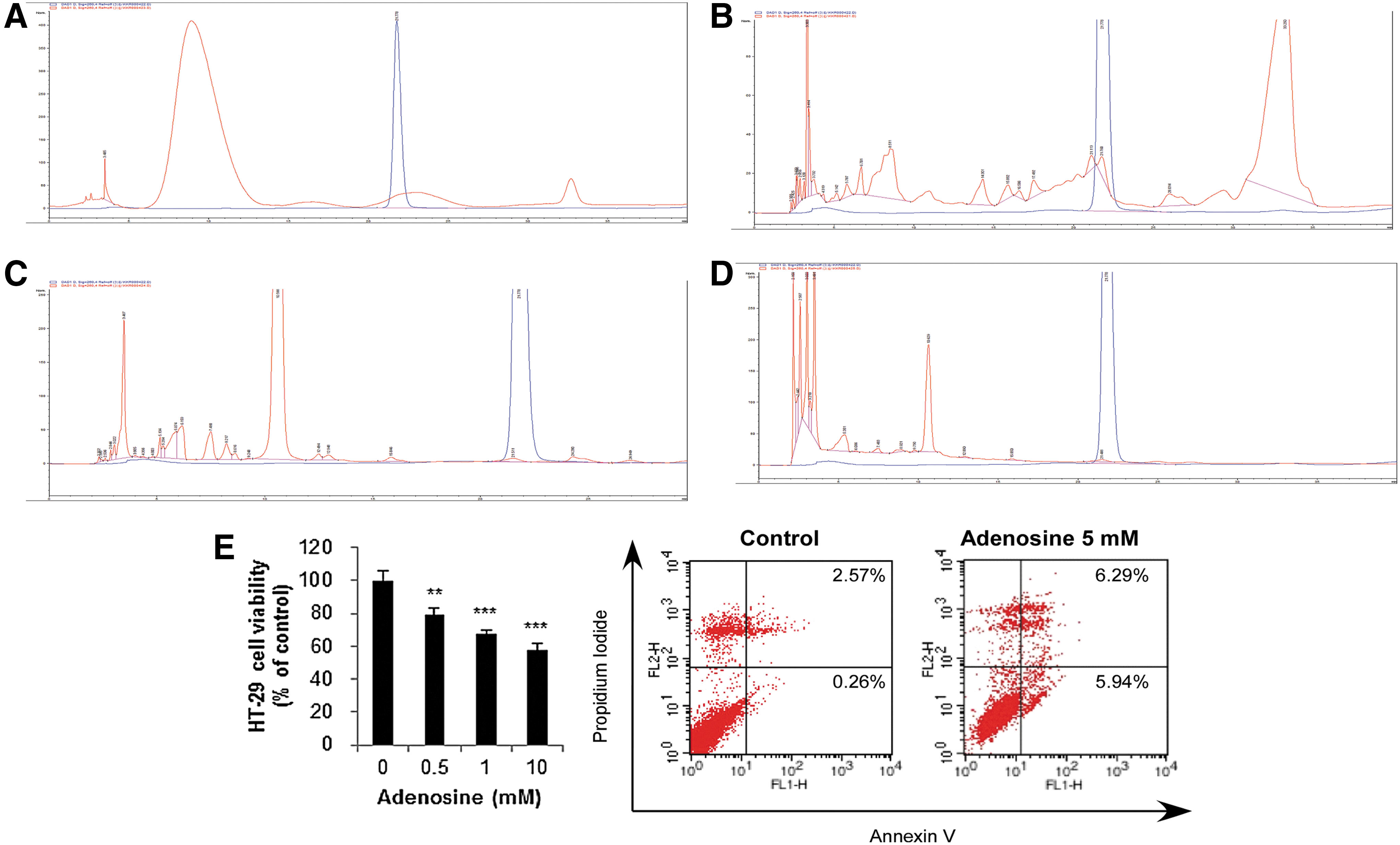
The chromatograms of
Discussion
Colorectal cancer is one of the most common cancers in the Western world and the second ranked cause of cancer morbidity and death. 40 Anticancer therapies cannot eradicate colon cancer due to drug resistance. 1 Therefore, it is necessary to develop novel anticancer drugs with reduced side-effects and better bioavailability. The use of active medicinal compounds or extracts from traditional medicines or other natural sources is considered one of the alternative treatment approaches of cancer. AC (Polyporaceae), popularly known as Niuchangchih or Changchih in Taiwan, is a widely used medicinal mushroom and shows multiple pharmacological activities, including anticancer and anti-inflammatory activities. 7,15 For massive production of AC, our group developed a novel culture method by inoculating AC on germinated brown rice (CBR). In this study, we elucidated the tumor-suppressive activity of CBR, using HT-29 cells. HT-29 cells are known to contain a point mutation at codon 273 (R→H) of p53 genes, which makes it less responsive to apoptosis inducers. 41
In this study, we compared the antiproliferative activity of CBR and AC. Interestingly, antiproliferative activity of CBR was more potent than AC. These results might be due to the presence of different bioactive compounds between CBR and AC EtOH extract. Identification of these bioactive ingredients will strengthen our hypothesis. Next, we compared the inhibitory effect of CBR hexane, EtOAc, BuOH, and water fractions on HT-29 colon cancer cell growth. CBR EtOAc fraction showed the most pronounced inhibitory activity on HT-29 cell growth among these fractions. Deregulation of the cell cycle machinery has been associated with cancer initiation and progression. 42 It has been reported that many anticancer agents arrest cell cycle at the G0/G1, S, or G2/M phases and then induce apoptotic cell death. 43 CBR EtOAc fraction resulted in an accumulation of HT-29 cells at G0/G1 cell phase. Cyclin D1 activate CDK4, which is mainly involved in regulating the G1/S restricting point. Cyclin D1 is overexpressed in ∼30% of human adenocarcinomas of the colon, which attributes to the abnormal growth and tumorigenecity 44 and therefore, is considered to be a good molecular target for cancer therapy or chemoprevention. 35,45 Cyclin E activates CDK2 protein right before starting the S phase. We found that CBR EtOAc treatment downregulated CDK4, cyclin D1 CDK2, and cyclin E expression in HT-29 colon cancer cells.
Annexin V assay data indicated that CBR EtOAc fraction inhibited the growth of the human colon cancer cells through apoptosis induction. The induction of apoptosis is almost always associated with the activation of caspases and apoptosis-related molecules. 46 Bcl-2 family proteins form a link between apoptosis and mitochondrial physiology. They mainly regulate the mitochondria initiated caspase activation pathway in apoptosis. Bcl-2, an antiapoptotic protein, preserves mitochondrial integrity and protects from cell death. Inversely, Bax, a proapoptotic protein, is expressed selectively during apoptosis promoting cell death. Our data indicated that CBR EtOAc fraction increased the expression of Bax, and suppressed Bcl-2 levels. The release of cytochrome c from mitochondria to cytosol after being induced by apoptosis-inducing agents results in the formation of apoptosome, composed of cytochrome c, apoptosis protease activating factor-1, deoxyadenosine triphosphate, and procaspase-9, which activates caspase-9. 47,48 We observed the release of cytochrome-c and the decrease in the expression of procaspases-9 in CBR EtOAc–treated HT-29 cells.
It is known that dysregulation of the Wnt signaling pathway triggers colon cancer initiation and progression 49 by overexpressing cyclin D1 and c-myc proteins. Thus, we hypothesized that CBR EtOAc fraction might exert an inhibitory effect against Wnt signaling pathway since CBR EtOAc fraction suppressed cyclin D1 expression in HT-29 cells. Aberrant β-catenin led to colon cancer development. In unstimulated cells, β-catenin is sequestered in the cytoplasm by the destruction complex (polyposis coil [APC], axin, and GSK-3β). However, upon stimulation of Wnt, the destruction complex gets inactivated, and β-catenin is translocated to the nucleus and serves as a cofactor for Tcf/Lef transcription factor. Nuclear β-catenin causes the transcription of several target oncogenes, such as cyclin D1, c-myc, and MMP-7. 6 CBR EtOAc fraction reduced the β-catenin expression in a dose-dependent manner in HT-29 cells. Many groups reported that inactivated GSK-3β leads to β-catenin stabilization. 50 A dramatic increase in phosphorylated GSK-3β (Ser9; inactive form of GSK-3β) is observed in various kinds of cancers, including late papillomas and squamous cell carcinomas. 51 –53 We observed that CBR EtOAc fraction reduced the expression of phosphorylated GSK-3β (Ser9; inactive form of GSK-3β) protein.
Several studies have demonstrated that extracellular adenosine induces apoptosis in various cancer cells through diverse signaling pathways, linked to adenosine transporters and adenosine receptors. 54,55 The stronger antiproliferative activity of the CBR EtOAc fraction on HT-29 cells might be due to the presence of adenosine since it was only detected in CBR EtOAc fraction. However, the 0.0196% concentration of adenosine detected in CBR EtOAc extract would not be enough to exert anticancer activity, but the adenosine in CBR EtOAc extract mighty enhance its anticancer activity by acting synergistically with other bioactive compounds in CBR. Therefore, we cannot rule out the enhanced anticancer activity of CBR due to the presence of other bioactive compounds.
In conclusion, this is the first investigation demonstrating that CBR has greater potential as a novel chemopreventive agent than AC against colon cancer. We found that the inhibitory effect of CBR EtOAc extract on the proliferation of colon cancer cells was due to the induction G0/G1 cell cycle arrest and apoptosis. Especially, we found that the CBR EtOAc fraction inhibited β-catenin signaling pathway, supporting its suppressive activity on the level of cyclin D1. These data suggest that CBR might be used as a chemopreventive agent against colorectal cancer.
Footnotes
Acknowledgments
This work was supported by the SMART Research Professor Program of Konkuk University, Seoul, Korea. We, the authors, would like to appreciate Sun mi Kim from Cell Activation Research Institute (CARI, Seoul, Korea) for assisting Annexin V assay and cell proliferation assay.
Author Disclosure Statement
No competing financial interests exist.