
Abstract
The present study examined, for the first time, the in vitro wound healing potential of chitosan green tea polyphenols (CGP) complex based on the activation of transglutaminase (TGM) genes in epidermal morphogenesis. Response surface methodology was applied to determine the optimal processing condition that gave maximum extraction of green tea polyphenols. The antioxidant activity, scavenging ability, and chelating ability were studied and expressed as average EC50 values of CGP and other treatments. In silico analysis and gene coexpression network was subjected to the TGM sequences analysis. The temporal expressions of TGMs were profiled by semi-quantitative reverse transcription (RT)-PCR technology within 10 days after wounding and 2 days postwounding. CGP showed the effectiveness of antioxidant properties, and the observations of histopathological photography showed advanced tissue granulation and epithelialization formation by CGP treatment. In silico and coexpression analysis confirmed the regulation via TGM gene family in dermatological tissues. RT-PCR demonstrated increased levels of TGM1–3 expression induced by CGP treatment. The efficacy of CGP in wound healing based on these results may be ascribed to its antioxidant properties and activation of the expression of TGMs, and is, thus, essential for the facilitated repair of skin injury.
Introduction
G
Multiple studies have demonstrated a beneficial effect of many growth factors on the process of healing, including experiments with exogenous growth factors for facilitating wound repair. 9 –12 In recent years, the study of genes that facilitate the activation of growth factors has revealed novel and specific roles in wound healing for a wide range of gene families. Transglutaminase (TGM) is a gene family of structurally and functionally related enzymes that stabilize protein assemblies. TGMs are characterized by a papain-like catalytic triad that is responsible for a Ca2+-dependent trans-mediating activity which catalyzes a range of reactions leading to post-translational modifications of proteins. 10,11 Generally, the unique structural features of tissue TGMs lead to a wide range of biological activities with physico-pathological implications. Different TGMs have previously been characterized and found to have specialized in the cross-linking of proteins in different biological processes. 11 Functions of TGMs include fibrin clot stabilization in hemostasis, semen coagulation, formation of envelopes in keratinization, and stabilization of extracellular matrix structures. 11,12 The essential role of TGMs in these processes was studied by the serious impairment of wound healing and keratinization. 12 Due to the multiple cellular functions, TGMs can potentially target the tissue repair process at different levels.
In contrast to those previous studies, little attention was given to TGM involvement in chemically induced keratinocyte differentiation. We hypothesized that chitosan green tea polyphenols (CGP) complex activates a differentiation pathway in normal epidermal keratinocytes and investigated the relationship of CGP complex-induced expressions of TGMs using an in vitro wound-healing model. The purpose of this study is to confirm TGMs activation by CGP complex treatment in contrast to antioxidant properties that may play a crucial role in wound healing by initiating the proliferatives phase of repair. Since dermal injury experiments have demonstrated up/down-regulation of growth factors, 13 changes in TGMs within it may be predominantly associated with the growth factor's secretion during hemostasis.
Materials and Methods
Chemicals and animals
EGCG, HCl, NaOH, and acetone were purchased from Sigma-Aldrich (Mississauga, Canada). Chitosan was obtained from the Department of Biomedical Engineering, University of Cincinnati (Cincinnati, OH, USA), with an N-deacetylation degree of 90% and an average molecular weight of 150,000. All animal experiments were conducted in accordance with accepted standards of humane animal care. Ten-week-old bred male mice weighing 150–180 g were obtained from the Research Institute of Surgery, China Pharmaceutical University. The mice were housed individually in cages and bred under controlled temperature (24°C±1°C).
Optimization of green tea polyphenols extraction
Since polyphenols are sensitive to environmental factors, it is important to develop an optimal extraction method to shorten the extraction time, reduce organic solvent consumption, and prevent environmental pollution. In general, efficiency of the extraction of polyphenols is influenced by multiple parameters such as temperature, time solvent content, and their effects may be either independent or interactive. 14 Response surface methodology (RSM) is a collection of mathematical and statistical techniques that are widely applied in the processing industry to determine the effects of several variables for optimizing conditions. 15 –17
In this study, a central composite design was performed with three independent variables on the extraction yield: X 1, extraction temperature (°C); X 2, MtOH concentration (%); X 3, extraction time (h). Generally, crude green tea polyphenols extraction was performed as described by Slinkard and Singleton. 18 The green tea leaves (10 g) were extracted for polyphenols by MtOH extraction with different concentrations (30%, 40%, 50%, 60% and 70%) for a given time (extraction time ranging from 2 to 6 h), while the temperature of the water bath ranged from 20°C to 80°C. After extraction, the mixture was centrifuged at 1238 g for 15 min, and the supernatant was collected. The residue was extracted twice under the same conditions. Supernatants from extraction were stored in a freezer at −30°C for analysis.
The RSM was applied to the experimental data using a commercial statistical package, Design-Expert version 7.1 (Statease, Inc., Minneapolis, MN, USA). Experiments were randomized in order to minimize the effects of unexplained variability in the observed responses due to extraneous factors. 19 The range of independent variables and their levels are presented in Table 1. The independent variables and their ranges were chosen based on preliminary experimental results. The adequacy of the model was checked, accounting for R 2 and adjusted-R 2 . Three-dimensional response surfaces and contour plots for the response are the graphical representations of the regression equations.
CGP complex synthesis
Since the process of transporting bioactive polyphenols through chitosan as a polymer can be consistent with Fick's diffusion law, the mechanism of interaction of polyphenol and chitosan could be ascribed to the development of noncovalent forces in equilibrium with the soluble complex. Since this equilibrium is irreversible under acidic conditions, these soluble complexes may aggregate and precipitate from solution according to Popa et al. 7 CGP complex was obtained by interaction between the reactants of polyphenols, and chitosan was prepared according to our previous study. 20 Chitosan was dissolved by stirring in 1 M HCl solution, followed by the neutralization with 1 M NaOH solution, up to pH 6.0. Afterward, the 1% aqueous polyphenols solution was added to chitosan, and the system was stirred continuously at room temperature for 6 h. After the reaction, the precipitate was separated through centrifugation at 2200 g for 10 min. The unreacted polyphenols were removed by repeated washing with distilled water. Finally, the product was washed with acetone and vacuum dried at room temperature. The polyphenol content was expressed as mg gallic acid equivalents (GAE) per 100 g of plant material.
Antioxidant assay
The total antioxidant activity of CGP complex and other samples (chitosan, EGCG, and polyphenols) were determined according as previously described. 21,22 The antioxidant ability was calculated as follows: Antioxidant activity (%)=[(Ac−As)/Ac]×100, where Ac is the absorbance of the resulting solution measured at 234 nm and As is the sample solution after reaction with 100%, indicating the strongest antioxidant activity. The reducing power was determined according to the method of Oyaizu (1986), and the absorbance was measured at 700 nm against a blank. 23 A higher absorbance indicates an increased reducing power. The scavenging ability was determined according to a previously published 1,1-diphenyl-2-picrylhydrazyl (DPPH) method. 24 Chelating ability was performed according to the method of Dinis et al., and the absorbance of the mixture was determined at 562 nm against a blank. 25 Half maximal effective concentration (EC50) value is the effective concentration at which the antioxidant activity was 50% or the absorbance was 0.5, which was obtained from linear regression analysis. Each experiment was repeated thrice (n=3).
Wound-healing assay
Twenty mice were randomly chosen and divided into two groups. Group A received distilled water as control; Group B received CGP complex with Average EC50 value (AE50) (3.50 mg/mL) according to our preliminary research. All animals were anesthetized with 3-cm×3-cm midline areas of their backs delineated and shaved. Two 10-mm-diameter full-thickness wounds were then placed on the dorsum of each mouse in the shaved area. Relative wound surface areas were compared 4 days after injury. For each animal, the wound surface areas were averaged. Excision wounds were formalin fixed and embedded in paraffin. Sections (5–10 μm) were stained with hematoxylin and eosin and photographed under 100× magnification.
In silico analysis
The TGM sequences of the mouse (Mus musculus) were obtained from MGI (
TGM, transglutaminase.
Semi-quantitative reverse transcription-PCR
Mouse skin keratinocytes cultures were performed adopted from Dlugosz et al. with some modifications. 28 Generally, the skins of mice were peeled off and floated on the surface of 0.4% trypsin at 4°C for 24 h. Epidermal sections were washed with phosphate-saline buffer (Sigma-Aldrich). The keratinocytes in the suspension were pelleted by centrifugation at 1000 g for 10 min and suspended in the serum-free medium of keratinocytes containing bovine pituitary extract. Total RNA was extracted with the RNeasy Micro kit (Qiagen, Valencia, CA, USA) and treated with RNase-free DNase I (Qiagen) to enable digestion. Reverse transcription (RT) was performed with Script Reverse Transcriptase (Bio-Rad, Hercules, CA, USA) to synthesize first-strand complementary DNA (cDNA). The resulting cDNA was used for PCR with the sequence-specific primer pairs listed in Table 3. The amplification was performed under the following conditions: one cycle at 94°C for 1 min, 35 cycles of 94°C (1 min)–55°C (1 min)–72°C (1 min), and, finally, one cycle at 72°C (10 min). Aliquots of the PCR products were subjected to 0.8% agarose gel electrophoresis and stained with ethidium bromide. The PCR products were visualized under UV light and photographed, which were scanned into a computer. Relative quantitative analysis was performed, and the ratios of target mRNA to housekeeping β-ACTIN mRNA concentrations were calculated. Each experiment was repeated thrice (n=3).
GC%, guanine–cytosine content.
Statistical analysis
Statistical analysis was performed using the general linear model of SAS 9.0 (SAS Institute, Inc., Cary, NC, USA, 2009). P<.05 was considered statistically significant.
Results
Modeling fitting and extraction procedure optimization
The experimental data for yield of green tea polyphenols (mg GAE/100 g) under different treatment conditions and polynomial response surface model fitted to the response variable (Y) are presented in Supplementary Table S1 (Supplementary Data are available online at
Significant at 0.05 level.
df, degree of freedom.
The results of the polyphenol yield affected by extraction temperature, time, and MtOH content are shown in Figure 1a–c, respectively. The maximum predicted value was shown by the surface confined in the smallest ellipse in the contour diagram. Figure 1a showed the interaction between extraction temperature and MtOH content at constant extraction time (4 h). It could be found that the yield of polyphenols increased with increasing MtOH concentration and temperature. Generally, higher temperatures and MtOH concentration result in a hydrolysis of a plant cell wall that favors polyphenols release and, thus, increases the polarity of solvent.
19
In addition, the polyphenols yield increased in a parabolic manner with a constant MtOH concentration (50%) as well as an increase in extraction temperature and treatment time (Fig. 1b). However, the yield of polyphenols was not pronounced with increases in time and MtOH concentration (Fig. 1c), probably due to the degradation of polyphenols. The suitability of the model for predicting optimum response values is presented in the following equation:

Response surface plots showing the effect of
These results demonstrated that a higher yield of polyphenols could be obtained under the optimal extraction conditions: at temperature 77°C for 6 h, and MtOH concentration at 71%. This set of conditions was determined to be feasible by the RSM approach and was also used to validate the experiments and to predict the values of the responses using the model equation.
Antioxidant properties
All samples showed consistent dose-dependent antioxidant activity and exhibited moderate to high antioxidant activities of 64%±2.2%–87%±0.8% at 10 mg/L (Fig. 2a). However, at 1 mg/mL, the antioxidant activities were 27%±1.2%, 35%±0.8%, 40%±1.7%, and 45%±2.5% for chitosan, EGCG, polyphenols, and CGP treatment, respectively. Among those, the CGP obtained from the interaction of chitosan and polyphenol was capable of increased inhibition of oxidation, Similarly, all samples showed consistent reducing power with an increased concentration. At 10 mg/mL, the reducing ability of CGP was 0.87±0.02; whereas others were in the range of 0.62±0.04–0.68±0.01 (Fig. 2b). However, the scavenging capability of polyphenols in DPPH radicals showed a slight decrease from 4–10 mg/mL, which may be due to the degradation of polyphenols. The scavenging ability of CGP was 75%±1.5% at 10 mg/mL (Fig. 2c), while chitosan, EGCG, and polyphenols showed scavenging abilities of 64%±1.3%, 68%±2.4%, and 70%±1.0% at 10 mg/mL, respectively. From 8 to 10 mg/mL, polyphenols and chitosan showed a plateau of scavenging abilities at 60%±1.4%–68%±2.5%, indicating that the chitosan and polyphenols were not as effective for DPPH radicals scavenging as CGP. At 10 mg/mL, the chelating ability of all samples on ferrous ions was in the range of 78%±1.1%–84%±1.0% of chitosan, EGCG, and polyphenols, respectively (Fig. 2d). It seemed as though all ferrous ions of CGP (92%±1.4%) were chelated at 10 mg/mL, but the effect of ion-chelating ability of CGP was complex. These results could be attributed to the CGP complex, enabling the binding ability with the phenol hydroxyl groups and amino groups of the chitosan molecules.
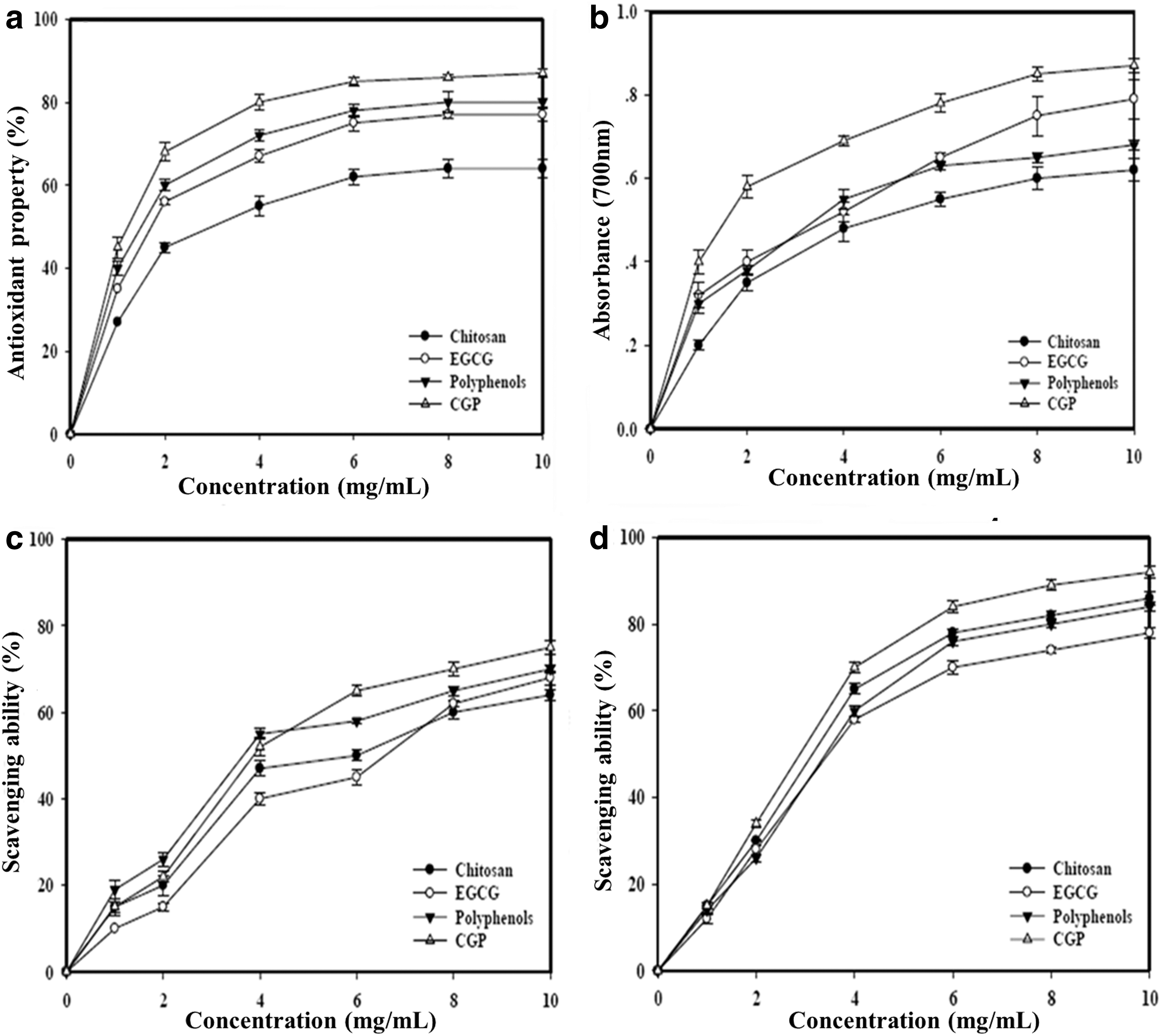
Antioxidant assay of CGP.
The antioxidant properties are presented in Table 5 and summarized as EC50 values for each assay. EC50 values were in the range of 3.10±0.24–5.30±0.16 mg/mL for polyphenols, EGCG, and chitosan, respectively, in which CGP (2.32±0.34 mg/mL) exhibited the greatest efficacy. Both EC50 values of reducing powers and scavenging abilities in DPPH radicals were in the range of 3.05±0.25–6.25±0.24 mg/mL and 5.19±0.37–6.48±0.45 mg/mL, indicating that CGP was most potent for these two antioxidant attributes. With regard to chelating abilities in ferrous ions, the EC50 values were in the range of 3.51±0.18–4.70±0.32 mg/mL. Overall, CGP was the most powerful in antioxidant activity, scavenging ability, and chelating ability in ferrous ions.
EC50 value: the effective concentration at which the antioxidant activity was 50%; the absorbance was 0.5 for reducing power; 1,1-diphenyl-2-picrylhydrazyl (DPPH) were scavenged by 50%; and ferrous ions were chelated by 50%, respectively. EC50 value was obtained by interpolation from linear regression analysis.
EC50 value obtained from linear regression analysis.
Values with the same letter in the same column do not differ significantly at P<.05; each value is expressed as mean standard deviation (n=3).
CGP, chitosan green tea polyphenols; EGCG, epigallocatechin-3-gallate.
Wound-healing assay
The mice were administered 1.0 AE50/kg body weight according to our preliminary test. The wound healing of control and CGP treatment were recorded as the days from injury to complete epithelization (Fig. 3). By day 4, wounds treated with CGP showed more epithelial tissue recovery than the control (Fig. 3a, c). On day 10, a notable wound closure occurred, because the epithelization of CGP treatment was complete (Fig. 3b). In contrast, wounds of controls healed more slowly at day 10 (Fig. 3d). Furthermore, the healing tissues of the wounds were evaluated by examining the histopathology of the CGP treatment and control samples at 4, 8, and 10 days after wounding (Fig. 4). At day 4, the epidermis was fully interrupted; the wound gap was filled with necrotic materials and inflammatory exudates; and no granulation tissue was observed at the time (Fig. 4a). CGP treatment demonstrated that wound gaps were filled with necrotic debris, and proliferation of blood vessels was formed with earlier granulation tissues (Fig. 4d). On the 8th day, the necrotic and inflammatory materials were filled by the proliferation of fibroblasts, which started filling up the gap (Fig. 4b). However, a large amount of necrotic debris was still observed. CGP treatment resulted in less acute inflammatory material, and the epidermis was smaller, which was filled by forming granulation tissues. The intact epidermal tissue was under growing and only a minimal gap was observed at the epithelial level (Fig. 4e), On the 10th day, the control group was still filling the gap and was superficially surrounded by fibrosis containing residual inflammatory debris (Fig. 4c). The CGP treatment of wounds improved significantly, the regrowth of the epidermis was nearly complete, and the wound scar was clearly formed (Fig. 4f). Therefore, the histological observations indicated that CGP was found to be effective in wound healing due to an acceleration of the reformation of granulation tissues in the wound-healing process.

Observation of excision wound healing:

H&E stained section of the skin tissue granulation of
In silico identification
TGMs have been proposed to participate in the process of wound healing, and they are activated in all layers of the skin, corresponding to the epidermis, the dermis, and the panniculus carnosus; particularly, plasma TGMs are activated by thrombin and contribute to the formation of a fibrin clot at the site of injury. 29,30 So, a comprehensive in silico investigation of the TGM gene family of M. musculus from the recently available online database, its chromosomal location, domain structure, phylogenetic tree construction, coexpression gene network generation, and array expression value collection have been attempted.
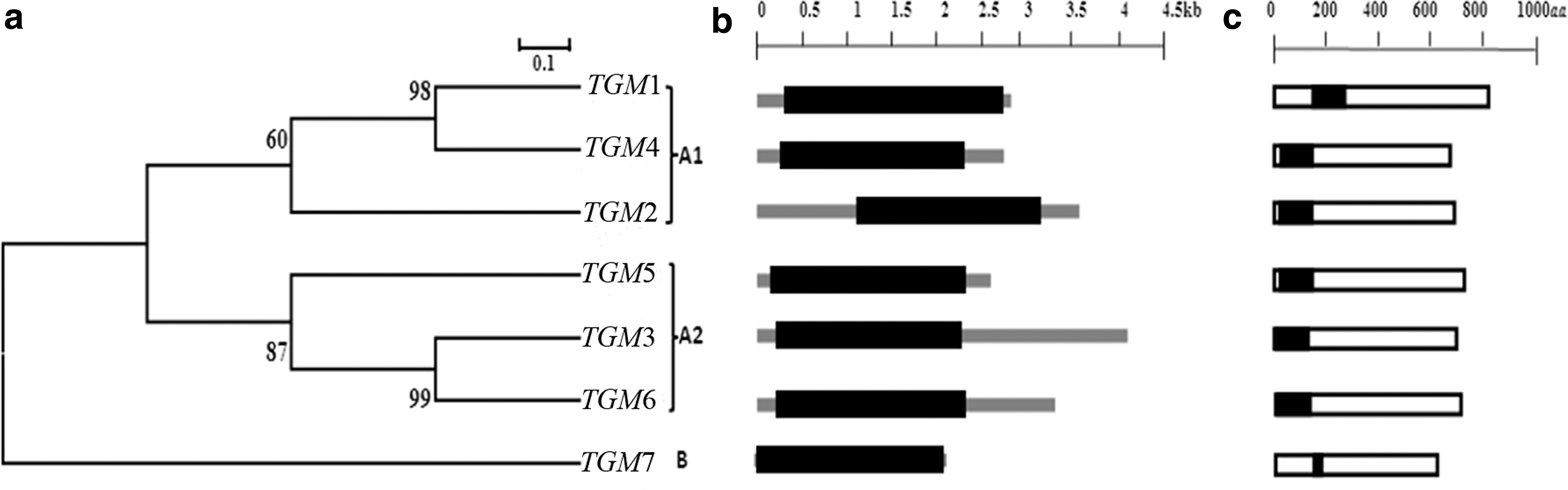
TGM sequences of M. musculus obtained from MGI were performed using various bioinformatics tools. The presence of the conserved TGM domain is a typical feature for consideration as a member of the TGM family. The domains of TGM proteins revealed highly conserved sequences in all the seven TGM proteins. The phylogenetic tree with bootstrapping values revealed the existence of two major groups. Group A was further divided into two subgroups: A1 and A2 (Fig. 5a). The TGM gene family with high bootstrapping values suggested common origins of TGM genes for each subgroup. The putative gene structures of the predicted TGM genes in terms of ORF distribution patterns are shown in Figure 5b, and the TGM7 seemed to contain a long ORF structure. However, the putative domains of TGM proteins with 109–117 amino acids were 100% conserved in six TGM proteins, while only TGM7 showed a variation and less amino acid residues (67 aa) (Fig. 5c). All the TGM genes were distributed on three chromosomes as provided in the chromosome map of MGI (Supplementary Fig. S1). Chromosome 2 maintained a maximum of five TGM genes, whereas the other two were located on chromosome number 9 and 14. The investigations suggested that TGM1, TGM2, and TGM3 were essential to early wound healing of the injured skin. The minor expression of TGM1 in simple epithelia, including endothelial cells, might play a role in wound healing. 31,32 TGM2 might be induced in response to acute-phase injury cytokines. 33,34 TGM3 could catalyze the substrates to form a complex that was related to the differentiation of the stratified squamous epithelia. 35,36 Thus, these TGMs may be involved in inflammatory and endothelial cell's construction by stabilizing the extracellular matrix, and the end product of the process is the replacement of the fibrin matrix with granulation tissue. 35,37 As the reason of that fact, coexpression data of TGM1, TGM2, and TGM3 in mouse were obtained from COXPRESdb. On that basis, a gene network of coexpressed genes for TGM1, TGM2, and TGM3 was constructed to focus on their functional relationships based on the correlation ranks. The density of genes in correlation spaces and the genes encoding the proteins in each organelle were identified by the KEGG annotations (Fig. 6). Genes that were strongly coexpressed were automatically picked, added to the network, and represented as small gray-shaded nodes. Generally, three coexpressed genes were directly associated with TGM1 as the following annotations: Hmox with the annotation of response to oxidative stress (GO: 0006979) and oxidation reduction (GO: 0055114), Ms4a6d with the annotation of membrane formation (GO: 0016020), and Crym with the annotation of cytoplasm formation (GO: 0005737) (Fig. 6a). Coexpressed genes directly connected to TGM2 were identified by the following GO annotations: Ccrl2 with the annotation of membrane formation (GO: 0016021) and plasma membrane formation (GO: 0005886), Tmem176p with the annotation of integral to membrane formation (GO: 0016021), Tinagl1 with the annotation of extracellular region construction (GO: 0005576), and cytoplasm formation (GO: 0005737) (Fig. 6b). Sprrs involved in the wound-healing process with the annotation of cytoplasm keratinization (GO: 0031424) was identified as the TGM3 coexpressed gene (Fig. 6c). Furthermore, the correlation of expression patterns among TGM1, TGM2, and TGM3 was found and shown in Supplementary Figure S2. The correlation value between TGM1 and TGM2 (0.285) was higher and similar to TGM2 and TGM3 (0.234) (Supplementary Fig. S2a, b). However, the correlation value between TGM1 and TGM3 was lower (0.208) (Supplementary Fig. S2c). To evaluate the expression patterns of TGM1–3 against their human (Homo sapiens) counterparts in the wound-healing process, the array express values of TGM genes in epidermis were collected from ArrayExpress (Supplementary Table S2). Accordingly, the expressions of TGM1–3 genes were highly involved in the up processes of building mouse skin epidermis, while the human TGM1–3 genes were involved in both up and down processes of constructing fibroblast and skin epidermis.
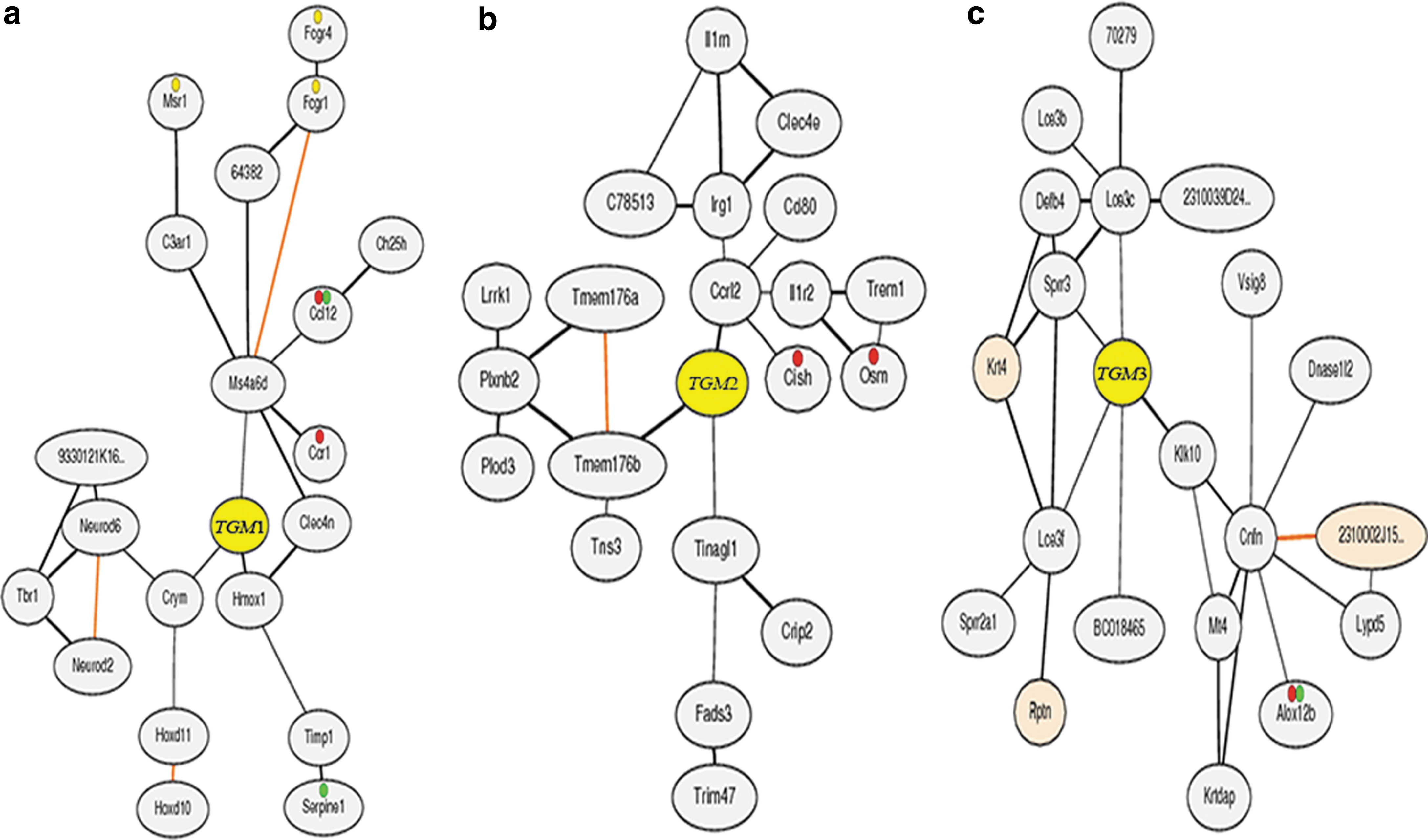
Co-expressed gene network of
Temporal expression of TGM
An in silico analysis indicated that TGMs, particularly TGM1–3, play important roles in the wound-healing process. Activation of TGMs by CGP was confirmed by expression profiling of each TGM mRNA in skin specimens by RT-PCR through 10 days after excision (Fig. 7) and 2 days posthealing. After excision, there was an increase in each TGM mRNA level. On comparing the mRNA expression levels against β-actin in the control group, it was shown that the peak level for TGM1 mRNA occurred at 24 h, while peak levels of TGM2 and TGM3 were at 72 and 168 h, respectively (Fig. 7a). All the mRNA levels decreased after the peak time and were normalized in 10 days in the control group. However, after CGP treatment, increased expression levels of TGM1 and TGM2 remained constant for 10 days, while the expression profile of TGM3 was similar to the control treatment (mouse received distilled water) (Fig 7b). Moreover, the ratios of the dramatic increase in expression levels of TGM1–3 were observed at 24 h after the wound healing (Fig. 8a). For control treatment, the expression values of CGP treatment appeared to be larger (Fig. 8b) at 24 h, after which the expression levels of all TGM1–3 were closely normalized for both the control and the CGP treatment group from 24to 48 h. According to the results, the significant increase in expression levels of TGM1–3 under CGP treatment after the excision wound model in the present study was correlated with the temporal expression levels of TGM genes. This result could be attributed to the activation of TGMs expression by CGP treatment, and the functionality of CGP treatment may be related to its antioxidant properties of skin function homeostasis.
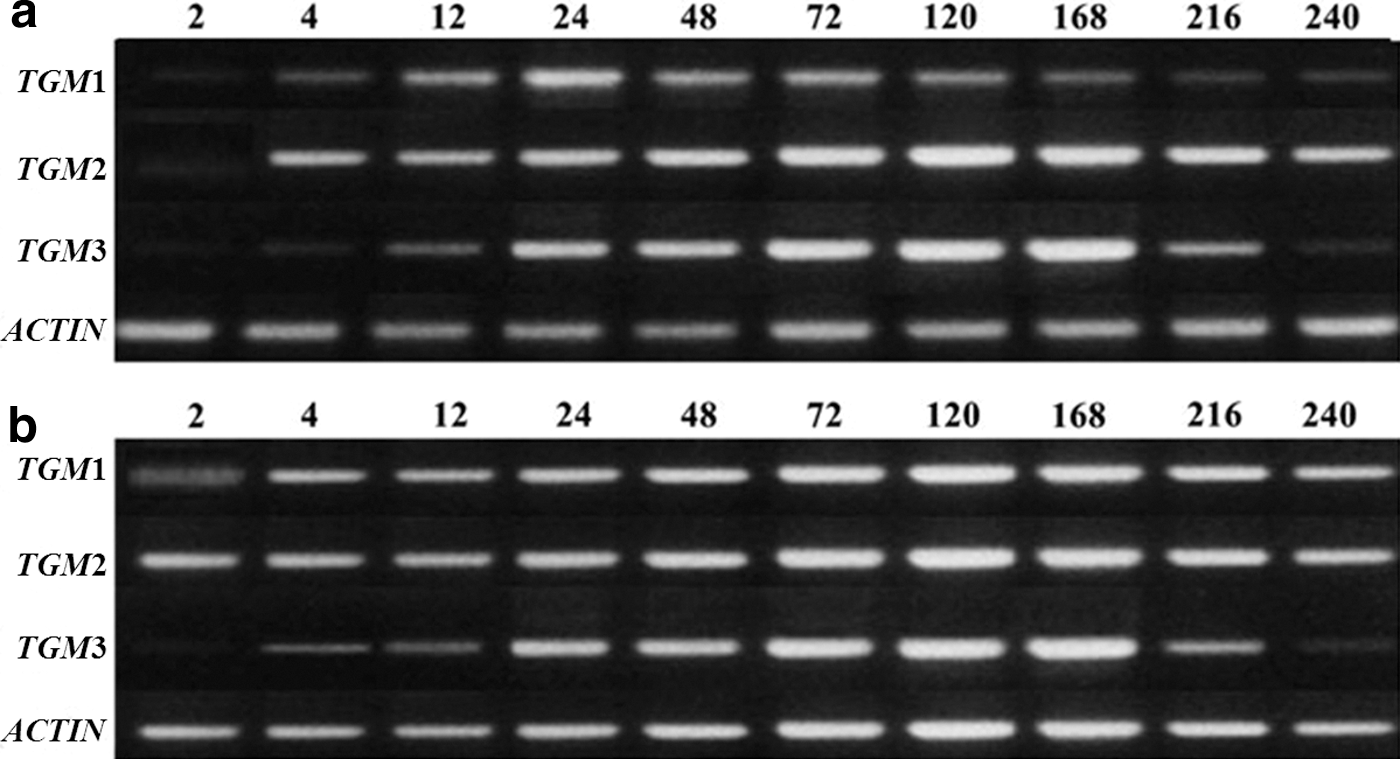
Reverse transcription-PCR analysis of mRNA expression of epidermis fibroblast TGM1–3 during skin wound healing in mice of
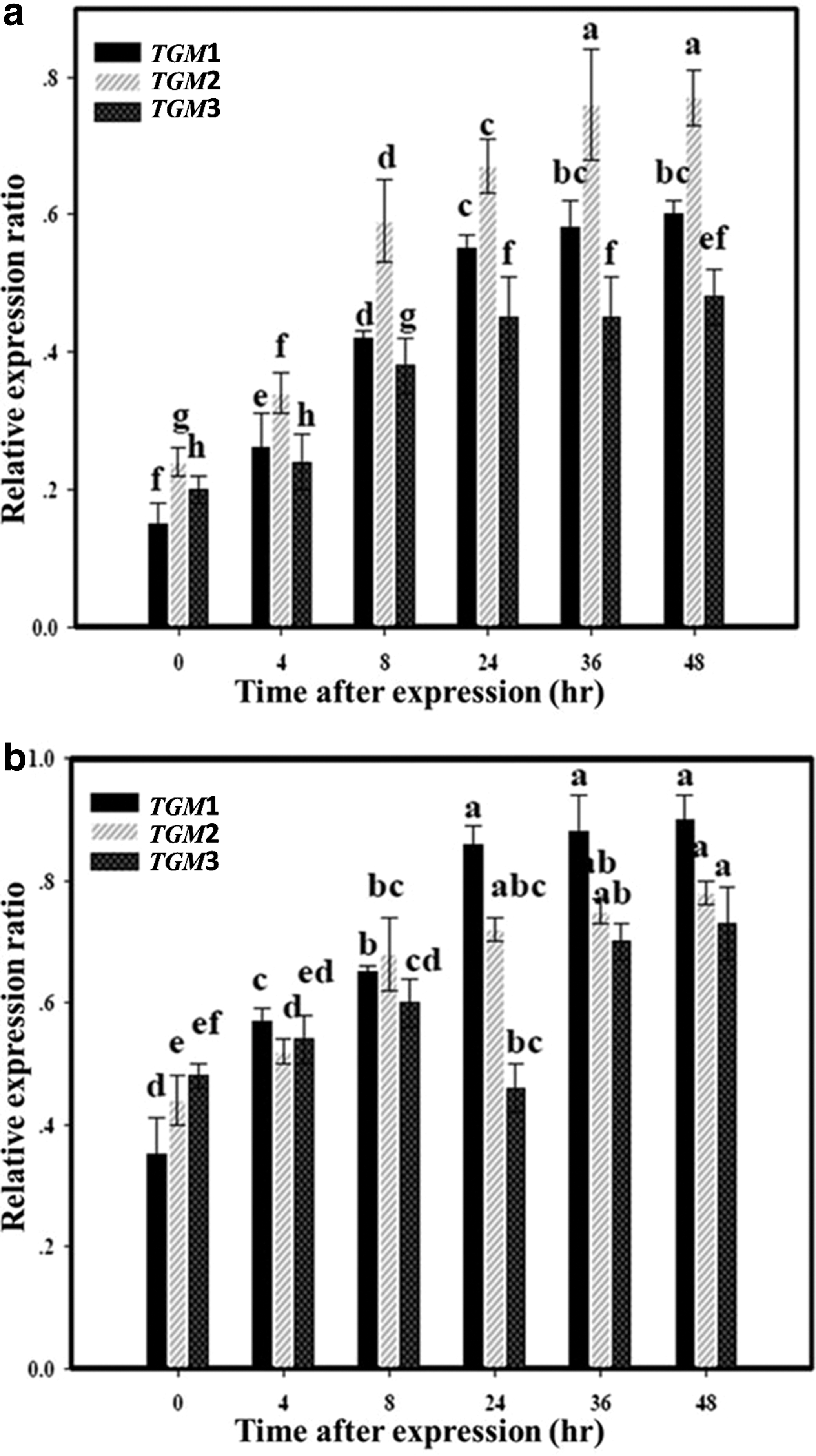
Temporal expression of TGM1–3 for epidermal fibroblasts after excision. The data of 0 h after excision represent the results obtained from normal mice:
Discussion
Wound healing is a complex process that starts with the formation of granulation tissue and ends with scar formation. In most of the cases, complications in wound healing are due to inflammation. 38,39 Inflammation results in a continuous generation of reactive species, such as the superoxide radical or the nonradical hydrogen peroxide. 39 Evidence for the potential role of oxidants in the pathogenesis of many diseases suggests that antioxidants may be of therapeutic use in these conditions. 40,41 Plant polyphenols have been shown to possess numerous biological properties, including hepatoprotective, antithrombotic, anti-inflammatory, and wound-healing activities, many of which may be related to their antioxidant and free radical scavenging ability. 42 Chitosan appears to have no adverse effects after implantation in tissues, and it has been used to inhibit fibroplasia in wound healing and to promote tissue growth and differentiation in culture. 43 Chitosan is widely recognized as the ideal wound covering that mimics many properties of the skin. Due to their biocompatibility, ability to absorb exudates, and film-forming properties, chitosan products are useful for burn and wound management. 44
An important step in this study was to determine the optimal extraction conditions for maximizing green tea polyphenols yield and the performance of the extraction of polyphenols by using RSM. Among the three parameters studied, extraction temperature and concentration of solvent had significant effects on the extraction rate of polyphenols. To facilitate the operation of verification experiment according to the mathematical model optimization analysis, the extraction conditions were adjusted as follows: ethanol concentration 70%, extraction time 6 h, and extraction temperature 77°C. Then, the process of CGP complex synthesis was studied through an interaction of green tea polyphenols and chitosan based on Popa's report. 7 Green tea has been reported to exhibit antimutagenic and cancer chemopreventive properties, and to prevent oxidative modification of DNA, proteins, lipid, and small cellular molecules by reactive oxygen species. 2 –4 In recent years, research on the role of antioxidants from tea polyphenols in wound healing has been studied. 1 –3 Antioxidant activities of chitosan derivatives have also been widely reported, including trapping peroxide radicals, inhibition of thiobartituric acid reactive substrate formation, and scavenging activities against hydroxyl radicals. 5,6 Chitosan, which provides a nonprotein matrix for tissue growth, also helps in natural blood clotting and blocks nerve endings to reduce pain. 5,6 Due to these unique antioxidant and wound-healing properties, green tea extraction and chitosan have been proposed for diverse applications in biomedical and pharmaceutical fields. The present research demonstrated that the green tea polyphenols, chitosan, and CGP complex have various degrees of antioxidant properties, including antioxidant activity, scavenging activities for DPPH radicals, and ion-chelating activity. These antioxidant effects may vary with the concentration; CGP complex exhibited the most potent antioxidant properties. In light of the activity observed in this study, the results revealed that CGP with presumed antioxidant properties may be used as an ingredient in the pharmaceutical industry. The histopathological study also confirmed the effectiveness of accelerating phases of tissue granulation, collagenation maturation, and scar maturation by CGP treatment. Due to the wound-healing process, the primary function of CGP in the acceleration of the wound-healing process could be related to the good presence of antioxidant properties.
Furthermore, this study also demonstrated that CGP significantly potentiated TGM1–3 activation during the skin wound-healing process. Coexpression values demonstrated that TGM1–3 were up-regulated in building mouse skin tissues. Meanwhile, RT-PCR profiles showed that the temporal characteristics of the TGM1–3 mRNA expression levels may have the potential to indicate the wound vitality. The activation of TGMs by CGP treatment showed several important properties in wound healing, which make it useful as a bio-agent for wound treatment. These results suggest that activation of TGMs by the antioxidant materials may be a viable approach for facilitating wound healing and tissue remodeling. Therefore, CGP is important not only for development and maturation of the epidermis, but also for the progression of cutaneous regeneration, and it is possibly involved in major changes in the tissue formation process.
In conclusion, the present research showed accelerated healing of excision wound by CGP complex. Although a couple of studies have demonstrated improved wound healing by plant polyphenols and chitosan, 2,3,5 the application of a controlled drug delivery mode for wound healing is a matter of concern, Controlled release of drugs to a given target generally involves prolonging the action of the active drug over time by allowing continual release from a polymeric dosage form. 45 There is, however, little literature on the controlled delivery of drugs from polymeric wound dressings. Previously, we reported the CGP complex, as a controlled released bio-agent hastened the cell growth in the epithelial layer of the skin properties via the incision wound model. 20 It is proposed to incorporate green tea polyphenols in the chitosan polymer to ensure the slow release and better antioxidant activities. Further biodegradation of chitosan material follows a course that is identical to normal wound healing. Moreover, CGP demonstrated the significant activation of TGM1–3 expression that could aid in the formation of the gene network of the wound-healing process as well. Hence, our results could be considered unique. Since CGP can play an important role in wound healing, this research suggests a scientific basis for its utility in medical treatment, and, thus, further studies using human skin wound models are required.
Footnotes
Acknowledgments
This research was supported by the grant of New Material Development, National Fund of Engineers, and Natural Science, People's Republic of China.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.