
Abstract
Certain bioactive peptides are reported to be able to alleviate hepatic fibrosis. Our previous work has confirmed the hepatoprotective effect of corn peptides (CPs) that are prepared from a high protein by-product, corn gluten meal, on acute liver injury in an animal model. However, the antifibrotic activity of CPs remained to be elucidated. In this study, the hepatoprotective effect of CPs on thioacetamide (TAA)-induced liver fibrosis was tested. Results showed that CPs (100 mg/kg body weight) significantly decreased the levels of alanine transaminase/aspartate transaminase, laminin, type IV collagen, and type III collagen in serum and increased the serum albumin levels and total antioxidant capacity. Additionally, with CP treatment (100 mg/kg body weight), a significant decrease was observed in the levels of malondialdehyde, nitric oxide, hydroxyproline, transforming growth factor β1, and lactate dehydrogenase activity as well as the liver index, while the activity of superoxidedismutase was significantly increased in livers. The histological and morphological analysis showed that the hepatocyte structure in CP-treated rats was superior to that of TAA-injured rats, and inflammation and fibrosis were also ameliorated. Therefore, CPs can be used as an option for prevention and adjuvant therapy of liver fibrosis.
Introduction
T
TAA, a well-known hepatotoxin, disrupts liver metabolism and causes hepatocellular necrosis. It is documented that drinking water containing 4 mM TAA for 10–12 weeks efficiently leads to cirrhosis in Wistar rats. 5 TAA has negative effects on DNA, RNA, and protein synthesis of enzymes in liver cells, prolonging the cell mitosis process and obstructing the transfer of RNA from the nucleus to the cytoplasm. The TAA-induced hepatic fibrosis injury model in rats is most similar to the pathology in humans, and also to the pathology of liver cirrhosis caused by virus. 6 TAA-induced hepatic cirrhosis has been observed to be associated with depletion of antioxidants and enhancement of lipid peroxidation. 7,8 Alleviating oxidative stress seems to play a positive role in preventing or reversing hepatic fibrosis. For this purpose, hydroxyl radical scavengers like selenium, 9 curcumin, 10 and taurine 11 have been applied in the treatment of TAA-induced liver cirrhosis.
Corn peptides (CPs), prepared from corn gluten hydrolysate, have been reported to possess superoxide dismutase (SOD)-like antioxidant activity, 12 scavenging effect on Diphenyl-picrylhydrazyl radical 13 and hydroxyl radical. 14 Our previous studies have confirmed the potent hepatoprotective effect of CPs on acute chemical liver injury induced by CCl4, alcohol, and D-galactosamine, 15 –17 and on the Bacillus Calmette-Guerin/lipopolysaccharide-induced immune liver injury model. 18 It is documented that hepatic fibrosis can be alleviated by bioactive peptides such as carnosine, 19 the vasonatrin peptide originating from animals, 20 and synthetic peptides. 21 In this study, we tested the hypothesis that CPs may have a hepatoprotective effect on chronic liver injury by evaluating the protective effect of CPs on TAA-induced liver fibrosis.
Materials and Methods
Chemicals and reagents
Alcalase was provided by Novozymes Co. (Copenhagen, Denmark). TAA was obtained from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). Rat laminin (LN), type III collagen (PC-III), type IV collagen (IV-C), hydroxyproline (HYP), and transforming growth factor β1 (TGF-β1) detecting ELISA kits were purchased from R&D Systems, Ltd. (Minneapolis, MN, USA). Aspartate transaminase (AST), alanine transaminase (ALT), malondialdehyde (MDA), SOD, lactate dehydrogenase (LDH), total antioxidant capacity (T-AOC), albumin (ALB), nitric oxide (NO), and Coomassie Brilliant Blue staining detection kits were purchased from Jiancheng Biological Engineering Institute (Nanjing, China). Standard feeds, which were nutritionally consistent with the national standard GB 14924.3-2010, 22 were obtained from the Hubei Laboratory Animal Center (Wuhan, China).
All other chemicals and reagents used were obtained from local sources and were of analytical grade.
Preparation of concentrated corn proteins
The concentrated corn proteins were prepared from a corn gluten meal provided by Zhengda Co. (Wuhan, China), as previously described by Guo et al. 16,18 The crude protein content was 83.08%, as determined by the Kjeldahl method.
Preparation of CPs
A 0.03 g/mL concentrated corn protein suspension was incubated at 90–100°C for 30 min. Subsequently, Alcalase was added at a 1% (w/w) ratio and the reaction conditions were set at 40°C, pH 8.0 in a reactor equipped with polymeric cellulose membranes with a 5 kDa molecular weight cutoff (MwCO) and a peristaltic pump. The level of reaction mixture in the vessel was maintained at a constant level by the addition of corn protein pretreated as described earlier. 16,18 The permeate fractions (Mw<5 kDa) were collected and lyophilized for the following research.
Treatment of animals
Wistar rats (8 week old, males, SPF, 180–200 g body weight [bw]) were purchased from the Hubei Laboratory Animal Center (Wuhan, China), and were housed in stainless steel wire-bottomed cages with free access to feed and water at 22±2°C with a 12-h light–12-h dark cycle. All animals received humane care and the studies reported here have been carried out in accordance with the Guidelines for the Humane Treatment of Animals set by the Association of Laboratory Animal Sciences at College of Animal Sciences and Technology, Huazhong Agricultural University.
After 1 week of acclimation, 40 rats were randomly divided into four groups. The duration of the study was 8 weeks divided into two phases of 4 weeks each. Group I, the control group, received neither TAA nor CPs in either phase. In phase 1, groups II (the model group), III (CPs, 100 mg/kg bw), and IV (CPs, 400 mg/kg bw) received TAA (200 mg/kg bw) intraperitoneally thrice a week and normal saline or CPs by gavage daily. In phase 2, group II received no treatment and a spontaneous recovery was observed. Groups III and IV continued to be administered CPs in the second phase to study the hepatoprotective role of CPs. The animal protocols are outlined in Table 1. At the end of the second phase, all animals were sacrificed under the ether anesthesia. Livers were removed, weighed, and dissected into two parts, one for biochemical studies and the other for histopathological examination. Blood samples were collected for subsequent analyses.
Notes: Standard laboratory conditions (12-h light/12-h dark cycles, temperature 22±2°C, relative humidity 50%) with feed and water provided ad libitum.
TAA, thioacetamide, 200 mg/kg bw, intraperitoneally; CP100, corn peptides, 100 mg/kg bw, i.g.; CP400, corn peptides, 400 mg/kg bw, i.g.; NS, normal saline, 1.0 mL/kg bw, i.g., intragastrically.
Serum biochemical analysis
Blood samples were centrifuged at 6000 g for 5 min at 4°C to obtain blood serum. The activities of AST/ALT and the levels of ALB, HYP, LN, PC-III, IV-C, and T-AOC were determined using analysis kits according to the manufacturer's protocols.
Liver biochemical assays
The livers were washed with cold physiological saline and connective tissue was removed. Liver homogenate was prepared with physiological saline at the concentration of 0.1 g/mL. The activities of SOD and LDH, and the levels of MDA, NO, TGF-β1, and HYP were determined using analysis kits according to the manufacturer's protocols.
Histopathological analysis
The liver tissue for histopathological analysis was fixed in 10% buffered formalin saline, dehydrated, and embedded in paraffin. Subsequently, the embedded tissues were cut into 4 μm sections, followed by staining with hematoxylin and eosin for general histology studies; the other sections were stained using the Masson-Trichrome (M-T) staining method, and marking the interstitial collagen blue. The histopathological characters were observed and recorded under a HPLAS-1000 colorized pathology image analyzer. The fibrosis degree of liver slides was graded according to the standard established by the Chinese Society of Hepatology. 23
Identification of CPs
The primary structure of CPs (Mw<5 kDa) was detected by high performance liquid chromatography (HPLC)-MS/MS, equipped with a ZORBAX SB-C18 column (4.6 mm×250 mm×5 μm, Agilent Co., Santa Clara, CA, USA). HPLC analysis with UV detection was performed by injecting thirty microliters of CP solution into the column at a flow rate of 1.0 mL/min at 25°C, with linear gradient elution (0–60 min, A 90–40%, B 10–60%), using a mobile phase of 0.1% trifluoroacetic acid solution (eluent A) and 0.1% trifluoroacetic acid acetonitrile solution (eluent B). The mass spectrometry was operated in the positive electrospray ionization (ESI+) mode, at an electrospray voltage of 3500 V, using high-purity nitrogen as the drying and ESI-nebulizing gas. Spectra were recorded in the mass/charge (m/z) range of 240–2100. The peptides were identified with LCMSD-Trap Data Analysis software and further validated using the online mass spectrometry database provided by the UCSF Mass Spectrometry Facility (see Supplementary Data, available online at
Statistical analysis
Quantitative data are expressed as mean±standard deviation and statistical comparisons were made by analysis of variance followed by the Tukey's test. The chi-squared test was used for statistical comparisons of the degree of fibrosis in liver slides. Differences were considered statistically significant when P<.05.
Results
Liver index
As shown in Figure 1A, a marked elevation in the liver index (a ratio of liver weight [g] to body weight [kg]) was observed in TAA-treated animals versus the control group (P<.01). The cotreatment and post-treatment with CPs (100 and 400 mg/kg bw) significantly ameliorated symptoms of hepatomegaly, which was supported by the decreased liver index compared to the TAA-treated group (P<.01).
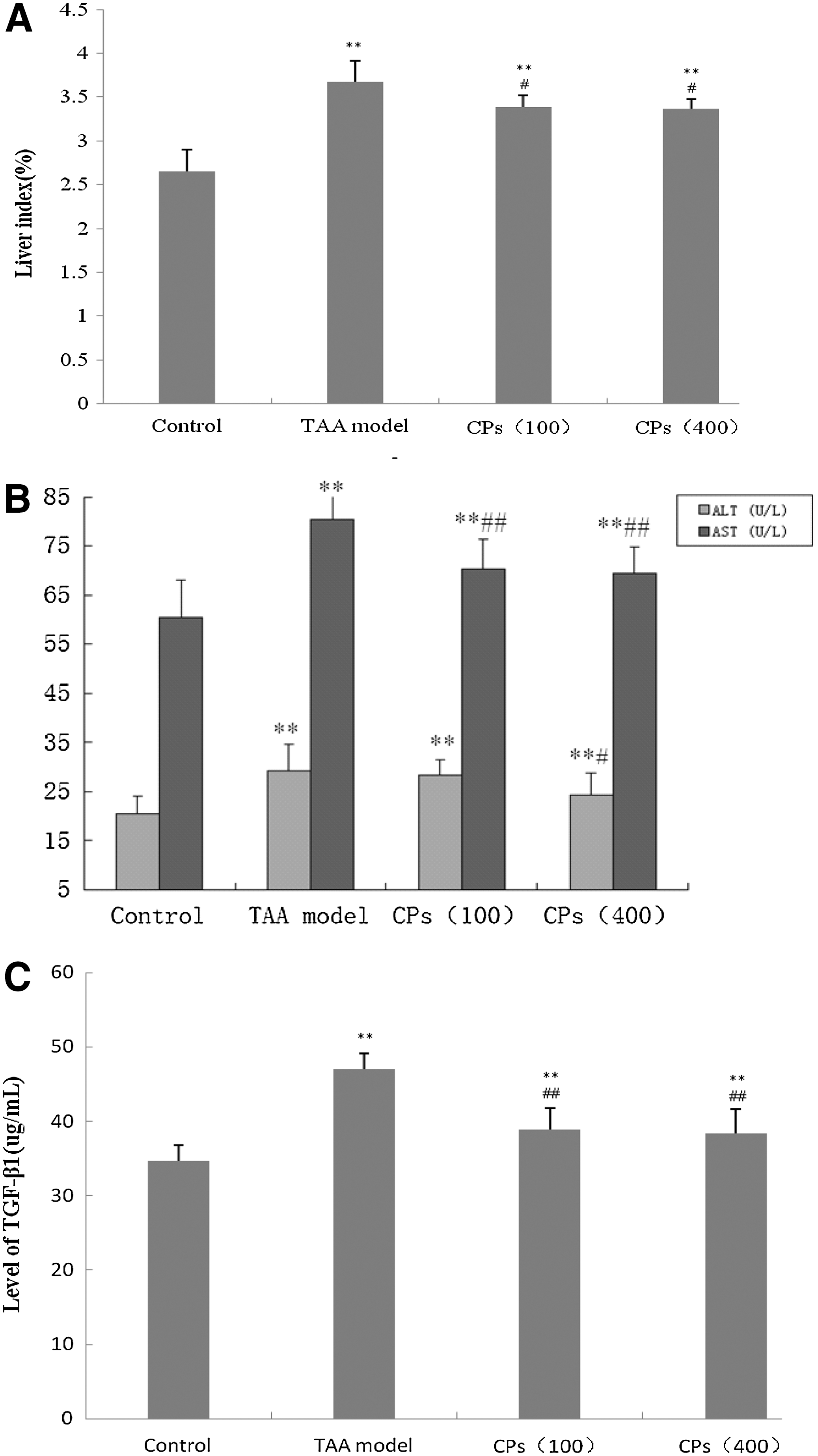
Effects of CPs on the liver index and biochemical markers.
Effects of CPs on indexes of liver function in hepatic fibrotic rats
As displayed in Table 2 and Figure 1B, TAA treatment significantly upregulated serum AST/ALT, LDH activity and NO levels, and downregulated serum ALB compared to the control group (P<.01). A significant decrease of serum AST/ALT activities was observed in animals treated with CPs (100 and 400 mg/kg bw) at P<.01, respectively. The content of ALB was dramatically enhanced at P<.01 in the CP-treated groups, suggesting that liver damage was alleviated. The obviously decreased LDH activity could be another index of improved liver function. The degree of inflammatory response (NO content) was also markedly lowered at P<.001. However, no significant difference was observed between the two CP-treated groups.
Data are mean±SE values.
P<.05, ** P<.01, *** P<.001, significantly different from the control group by the Tukey's test.
P<.05, ## P<.01, significantly different from the TAA-induced model group by the Tukey's test.
ALB, albumin; LDH, lactate dehydrogenase; NO, nitric oxide.
Effects of CPs on antioxidant capability in hepatic fibrosis rats
As shown in Table 3, TAA treatment disrupted the balance of the REDOX system, inhibiting the SOD activity, exhausting the T-AOC value, and increasing the MDA content when compared to the control group (P<.001). Additionally, the liver SOD activity and the serum T-AOC value were obviously increased with CP treatment (P<.05), while the liver MDA production was significantly decreased (P<.001).
Data are mean±SE values.
P<.05, *** P<.001, significantly different from the control group by the Tukey's test.
P<.05, ## P<.01, ### P<.001, significantly different from the TAA-induced model group by the Tukey's test.
MDA, malondialdehyde; SOD, super oxygen dehydrogenase; T-AOC, total antioxidant capability.
Effects of CPs on indexes of liver fibrosis in hepatic fibrotic rats
As displayed in Table 4, the four hepatic fibrosis indicators of IV-C, LN, PC-III, and HYP were significantly increased in the model group compared to the controls (P<.01). However, treatments with CPs (100 and 400 mg/kg bw) reversed the elevation of IV-C and LN (P<.05). In addition, a dramatic decrease was observed in the PC-III and HYP in the CP (100 and 400 mg/kg bw) groups compared with those receiving TAA only (P<.01).
Data are mean±SE values.
P<.05, ** P<.01, *** P<.001, significantly different from the control group by the Tukey's test.
P<.05, ## P<.01, significantly different from the TAA-induced model group by the Tukey's test.
IV-C, type IV collagen; LN, laminin; PC-III, type III precollagen; HYP, hydroxyproline.
Effect of CPs on production of liver TGF-β1 in hepatic fibrosis rats
As revealed in Figure 1C, when compared to the control group, the level of hepatic TGF-β1 was significantly higher in the TAA group (P<.001). CP treatment markedly lowered the content of liver TGF-β1 (P<.001).
Histopathological evaluation
A normal rat liver shows a limpid central vein and hepatic cells with prominent nuclei and uniform cytoplasm. As shown in Figure 2A, the hepatic lobular architecture was normal, and no connective tissue proliferation was observed. However, the rat livers treated with TAA for 4 weeks showed serious degeneration of hepatocytes with cell necrosis and monocyte infiltration, including marked fatty degeneration, necrosis, and inflammatory cell infiltration. As demonstrated in Figure 2B, histological analysis using M-T staining showed obvious collagen deposits as well as large fibrous septa. Administration of CPs significantly attenuated the deposition of collagen fibers. CP treatment attenuated hepatocyte injury and the deposition of collagen fibers in a dose-dependent manner. As shown in Figure 2C-ii and 2D-ii, livers showed only a few inflammatory cells and short bundles of collagen, especially the ones treated with CPs (400 mg/kg bw).

Photomicrographs of liver sections. The sections were stained with hematoxylin and eosin (H&E)
The degree of fibrosis revealed by liver slides is shown in Table 5, which was graded according to the standard established by the Chinese Society of Hepatology and was categorized into five grades (S0–S4) based on severity. Statistical comparisons of the fibrosis degree of liver slides were made by means of the Chi-Square Test. Results showed that almost no inflammation was observed in group I, whereas the group II was mainly focused on S3 and S4 (S3, 20%; S4, 80%). Fibrosis symptoms were significantly ameliorated in CP treatment groups compared with group II (P=.003 and 0.001, respectively), wherein no S4 was observed. With an increase in CP concentration, S1 increased from 20% to 60% and S3 decreased from 50% to 10%. However, there was no significant difference between the two different dosages (P=.097).
Grades: S0, there is no or slight inflammation around the portal area, but no symptom of liver fibrosis; S1, inflammation and fibrosis merely concentrated around antrum and hepatic lobule; S2, fibrous septum around the portal area appeared and the inflammatory response was slighter; S3, fibrous septum around the portal area existed, the structure of hepatic lobule disordered, and the inflammatory response was more severe; S4, early cirrhosis formed and the inflammatory response was heavy.
Identification of CPs
As shown in Figure 3, six major peaks in the total ion current chromatogram of CPs were numbered in order according to the retention time. The primary structure was identified and summarized in Table 6. Except for the hexapeptide Pro-Val-Val-Leu-Ile-Asp (PVVLID), a terminal phenyl group was found.
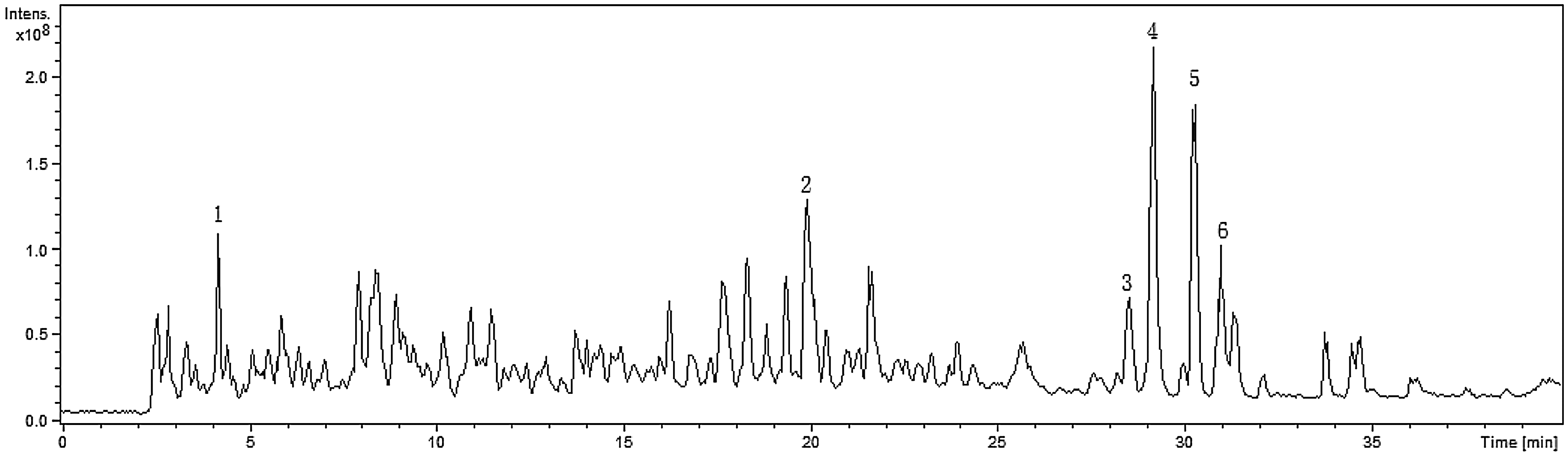
Total ion current chromatogram of CPs (Mw<5 kDa).
Discussion
To date, due to specific disadvantages, such as cost, side effects, and poor therapeutic potentials, no satisfying treatment or therapeutic drug for liver diseases has been established. Therefore, finding effective ways to inhibit liver fibrosis and prevent the progression to cirrhosis is of significance. In this study, we demonstrated that corn gluten hydrolysate CPs are effective for prevention and treatment of liver fibrosis induced by TAA, and improve the hepatic biochemical indexes in CP-treated rats to a different extent, indicating the hepatoprotective activity of CPs on chronic liver injury.
AST/ALT are aminotransferases in cells, the activities of which are conventional and sensitive indicators of liver injury. 4 Increasing AST/ALT levels indicate an inflammatory state in liver cells that might lead to hepatic fibrosis. 24 In chronic liver diseases such as alcoholic hepatitis, hepatic fibrosis, and cirrhosis, the serum ALB levels are decreased due to protein synthesis disorder in hepatocytes. 25 NO and LDH are closely related to the liver damage when their contents increased abnormally. The membrane injury and lipid peroxidation of hepatocytes are a latent source of fibrogenic reactive oxygen species (ROS), which were demonstrated in almost all the clinical and experimental liver fibrosis. 26 In this study, these effects were markedly attenuated, indicating that the antifibrotic effect of CPs may be attributed, at least in part, to the protective effect on hepatocytes against inflammatory responses and/or oxidative stress.
In hepatocytes, TAA is metabolized into TAA sulfoxide and TAA-S,S-dioxide by the enzyme CYP2E1. These products covalently bind other intracellular molecules and cause oxidative stress. 27 Oxidative stress has been well recognized as a vital contributor to the pathophysiologic process of hepatic fibrogenesis. 28 Inverse correlations between antioxidant enzymes and lipid peroxidation have been found in rats with TAA-induced liver fibrosis. 29 Thus, antioxidants such as tocopherol, silymarin, phosphatidylcholine, Sho-saiko-to and Ipomoea aquatica extract have been tested to verify the capacity to attenuate liver fibrosis in experimental models. 30,31 CPs have a potent antioxidant activity, suggesting potential hepatoprotective effects on acute chemical liver injury in mice. 15 In this study, the antifibrotic effect of CPs might be associated with their antioxidative capability.
Antioxidants can decrease hepatic stellate cell (HSC) activation in culture. 32 It is generally accepted that HSC is an important target cell for the treatment of hepatic fibrosis. 33 TGF-β is the only cytokine that can stimulate HSC product matrix components. 34 TGF-β has three different isomers, but the most important one is TGF-β1. Downregulation of TGF-β1 expression and modulation of TGF-β/Smad signalling may be effective in preventing liver fibrosis. 35 Our experimental results revealed that the secretion of TGF-β1 was suppressed in rats receiving CPs, which may have contributed the prevention of fibrosis.
The increased synthesis and decreased degradation of the ECM is a characteristic of hepatic fibrosis. The hepatic collagen content reflects the overall load in the liver, an important marker for fibrosis. 36 HYP, rare amino acids in other proteins, is a major component of the collagen and plays a crucial role in collagen stability. CPs can decrease the serum levels of PC-III, IV-C, LN, as well as the HYP content. Thus, preventing the synthesis of collagen or enhancing collagenolysis could be an important therapeutic mechanism by which CPs remedy liver fibrosis.
When chronic liver injury occurs, intracellular components released from necrotic cells can activate immune cells and provoke the process of ROS-mediated cell death, which triggers more oncotic necrosis and amplifies inflammation. 37 Sustained inflammation leads to overactivation of HSC and subsequent hepatic fibrosis. The histological and morphological analysis indicated that inflammation and fibril connective tissue were significantly decreased in rats treated with CPs, further confirming the antifibrotic effect of the CPs. Furthermore, hydrolysate was reported to be used for clinical treatment of liver cirrhosis, with an F-value (a molar ratio of branched-chain amino acid and aromatic amino acid) of higher than 20 and a Tyr and Phe content of less than 2% of the total amino acid residues. 38 Abundant in branched-chain amino acids, the corn protein can be used to prepare high F-value oligopeptides. 39 The high F-value hexapeptide PVVLID was identified in the CPs and Leu, a branched amino acid in all the identified peptides. However, the relationship of antifibrotic effects and closely connected Leu needs further study.
Conclusion
To the best of our knowledge, this study presents the first experimental evidence that CPs exert antifibrotic effects on TAA-induced liver fibrosis as evidenced by significantly decreased levels of ALT/AST, LN, IV-C, and PC-III in serum and increased ALB levels and T-AOC. In addition, the levels of MDA, NO, HYP, TGF-β1, and LDH activity were markedly decreased by CPs, while the SOD activity was significantly increased in the liver. The mechanism of CP action might be associated with the protective effect on hepatocytes due to the antioxidative activity as well as the inhibition of TGF-β1 secretion. Our results suggest the possibility that CPs may be useful as a preventive agent and adjuvant therapy for treating liver fibrosis.
Footnotes
Acknowledgments
The authors gratefully thank the National Natural Science Foundation of China (Grant No. 30972043) and the National 863 Program of China (Grant No. 2008AA10Z314) for financial support of this research.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.