
Abstract
The identification of the active compounds of herbal medicines and the molecular targets of those compounds is an attractive therapeutic objective. Reynoutria elliptica has been used for the treatment of various inflammatory diseases as a Korean folk remedy. Based on the evidence that anti-inflammatory agents frequently exert antiproliferative activity, we tested two sesquiterpene derivatives, 8-hydrocalamenene (HC) and 8,14-dihydrocalamenene (DHC), for their ability to induce apoptosis and suppress signal transducer and activator of transcription 3 (STAT3) activation in multiple myeloma (MM) U266 cells. We found that HC inhibited cell viability in U266, but not in peripheral blood mononuclear cells. HC exerted significant cytotoxicity and induced substantial subG1-phase arrest and apoptosis as compared with DHC. HC inhibited the expression of gene products involved in antiapoptosis (Bcl-2 and Bcl-xL), proliferation (cyclin D1), and invasion (MMP-9), all of which are known to be regulated by STAT3. Furthermore, HC up-regulated cyclin-dependent kinase inhibitor p21 and induced apoptosis through the activation of caspase-8, -9, and -3 in U266 cells. Interestingly, HC blocked constitutive STAT3 activation through the inhibition of activation of upstream kinases Janus-like kinase 1 (JAK1), JAK2, and c-Src and up-regulated PIAS3. Deletion of STAT3 reversed cytotoxic effects and the down-regulation of cyclin D1 and c-myc by HC in MM cells. Finally, this sesquiterpene significantly synergized the cytotoxic and apoptotic effects of bortezomib in U266 cells. Taken together, these results suggest that HC is a novel blocker of STAT3 activation which may have a potential in the prevention and treatment of MM.
Introduction
H
Recently, the root of the plant has been reported to exhibit several beneficial biological effects such as antineuraminidase 1 and anti-inflammatory 2 activities. Specifically, these studies have shown that four active compounds, including emodin, physcion, omega-hydroxyemodin, and trans-resveratrol, derived from the root, exhibit high neuraminidase inhibitory activity 1 and the hexane fraction of the plant inhibits LPS-induced production of inflammatory markers by blocking nuclear factor-kappaB (NF-κB) and MAPKs signaling in RAW 264.7 cells. 2 Here, we have purified two compounds designated HC and DHC from Reynoutriae Radix to evaluate their potential antiproliferative effects, because chronic inflammation has been found to be closely linked with cancer. Chronic inflammation increases the risk for various cancers, indicating that eliminating inflammation may represent a valid strategy for cancer prevention and therapy. 3 Moreover, the molecular mechanisms underlying the anti-inflammatory and antiproliferative effects of HC and DHC are not yet fully understood.
Since the signal transducer and activator of transcription (STAT) pathways have been closely associated with inflammation, survival, and apoptosis, it is possible that the agents from the root mediate their effects through modulating the STAT pathways. The STAT proteins were identified in the last decade as the transcription factors that were critical in mediating all cytokine-driven signaling. 4 Seven known mammalian STAT proteins, STAT1, 2, 3, 4, 5a, 5b, and 6, are involved in cell proliferation, differentiation, and apoptosis. 5 One STAT family member, STAT3, is often constitutively active in various human cancer cells, including multiple myeloma (MM), leukemia, lymphoma, and prostate tumors. 6 –8 Once activated, the phosphorylated STAT3 is subsequently translocated into the nucleus, where active STAT3 regulates the transcription of its target genes. The phosphorylation of STATs is mediated through the activation of non-receptor protein tyrosine kinases called Janus-like kinase (JAK). JAK1, JAK2, JAK3, and TYK2 have been linked to the activation of STAT3. 9 In addition, the role of c-Src kinase has been demonstrated in STAT3 phosphorylation. 10 STAT3 has been known to regulate the expression of genes that participate in oncogenesis, such as apoptosis inhibitors (Bcl-xL and Bcl-2) and cell-cycle regulators (cyclin D1). 7,11 Therefore, agents that suppress constitutive STAT3 activation have implications for the prevention and treatment of cancer. 12,13
Due to the critical role of STAT3 activation in tumorigenesis, including survival and proliferation, we hypothesized that HC and DHC mediate their effects, in part, through the suppression of STAT3 signaling. In the present report, human MM U266 cells were used for most experiments, because these cells express constitutively active STAT3. Whether HC and DHC can modulate the constitutive STAT3 activation in these MM cells, was investigated. The results that follow indicate that HC but not DHC suppressed constitutive STAT3 activation and down-regulated the expression of cell survival and proliferative gene products, leading to the suppression of the proliferation and induction of apoptosis in human MM U266 cells.
Materials and Methods
Reagents
8-Hydrocalamenene (HC) and 8,14-dihydrocalamenene (DHC) with purities greater than 95% were kindly provided by Natural Research Center in KIST, Republic of Korea. HC and DHC were dissolved in dimethylsulfoxide as a 100 mM stock solution and stored at −20°C. Further dilution was done in cell culture medium. 3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenyltetrazolium bromide (MTT), dimethyl formamide, Bortezomib, bovine serum albumin, Tris, glycine, NaCl, and sodium dodecyl sulfate (SDS) were purchased from Sigma-Aldrich (St. Louis, MO, USA). RPMI 1640 and fetal bovine serum (FBS) were purchased from Invitrogen (Carlsbad, CA, USA). Polyclonal antibodies to STAT3 and phospho-STAT3 (Tyr705) and Bcl-2, Bcl-xL, caspase-3, survivin, MMP-9, cdc2, p21, multi-drug resistance (MDR), IAP-1, poly(ADP ribose) polymerase (PARP), protein inhibitors of activated STAT1 (PIAS1), protein inhibitors of activated STAT3 (PIAS3), c-myc, and goat anti-rabbit and goat anti-mouse HRP were purchased from Santa Cruz Biotechnology (Santacruz, CA, USA). Antibodies of phospho-JAK2 (Tyr1007/1008), JAK2, cyclin D, caspase-8, cleaved caspase-8, caspase-9, and cleaved caspase-9 were purchased from Cell Signaling Technology (Danvers, MA, USA).
Cell lines
Human MM U266 cells was obtained from American Type Culture Collection (ATCC, Manassas, VA, USA) and cells were grown in RPMI 1640 medium that was supplemented with 10% FBS, penicillin (100 units/mL), and streptomycin (100 μg/mL) at 37°C in a humidified atmosphere with 5% CO2.
Isolation of human peripheral blood mononuclear cells
Human peripheral blood mononuclear cells (PBMC) were isolated from the blood of healthy adult volunteer donors by density gradient centrifugation on Lymphoprep (Axis-Shield PoC AS, Oslo, Norway).
MTT assay
We used different numbers of cells for the exposure of HC or DHC in U266 cells (2000 cells/well) or PBMC (5000 cells/well), because optimal absorbance depends on cell size and cell mass during the growth of each cell types. Cell viability was measured by an MTT assay to detect NADH-dependent dehydrogenase activity. Fifty microliters of MTT solution (5 mg/mL) in 1× phosphate-buffered saline (PBS) was directly added to the cells, which were then incubated for 4 h to enable MTT to metabolize to formazan. Absorbance was measured with an automated spectrophotometric plate reader at a wavelength of 570 nm. Cell viability was normalized as relative percentages in comparison with nontreated controls.
Cell cycle analysis
U266 cells were seeded into six-well plates at a density of 1×106 cells per well and incubated for 2 h. After treatment with 0, 20, or 40 μM of HC and DHC for 24 h, the cells were collected and washed with 1× PBS. Cell pellets were fixed in 70% cold ethanol overnight at −20°C. The fixed cells were resuspended in 1× PBS containing 1 mg/mL RNase A and incubated for 1 h at 37°C, and the cells were stained by adding 50 μg/mL PI for 30 min at room temperature in the dark. The DNA contents of the stained cells were analyzed using CellQuest Pro software with an FACS Calibur (Becton Dickinson, Heidelberg, Germany) flow cytometer.
Annexin V assay
U266 cells (1×106 cells per well) were treated with 0, 20, or 40 μM of HC and DHC for 24 h, stained by annexin V conjugated to fluorescein isothiocyanate (FITC). The cells were washed with cold PBS, resuspended in 100 μL of 1× binding buffer, and 1.25 μL of annexin V was added for 30 min at 37°C and 10 μL of PI was added for 10 min in the dark. Then, 400 μL of 1× binding buffer was added to each tube and analyzed with a flow cytometer.
TdT-mediated dUTP nick end labeling assay
Individual apoptotic cell death was observed using a TdT-mediated dUTP nick end labeling (TUNEL) assay kit. For a flow cytometer assay, U266 cells were seeded onto six-well plates at a density of 1×106 cells per well, incubated overnight, and then treated with 0, 20, and 40 μM of HC and DHC for 24 h. The cells were collected and washed with 1× PBS. Cell pellets were fixed with 4% paraformaldehyde for 30 min at room temperature. The fixed cells were permeabilized by 0.2% Triton X-100 in 1× PBS for 15 min, washed, resuspended in 1× PBS containing TUNEL reaction solution, and incubated for 1 h at 37°C in the dark.
Cell transfection and siRNA knockdown
To knock down STAT3 expression in U266 cells by RNA interference, U266 cells (1×106 cells per well) were transfected with 100 nM scrambled siRNA (sc-37007; Santa Cruz Biotechnology) and STAT3 siRNA (sc-29493; Santa Cruz Biotechnology) using NEON Transfection System (Invitrogen) according to the manufacturer's protocol.
Reverse transcriptional polymerase chain reaction
The total RNA was isolated using the TRIZOL Reagent (Invitrogen). Total RNA was reverse transcribed into cDNA using an oligo (dT)15 premix (Intron, Seongnam, Korea). The following forward and reverse primer sequences were used for STAT3, 5′-ACC GTA AGT GGC TTC CTT TC-3′ and 5′-CAT CGG CAG GTC AAT GGT AT-3′. Polymerase chain reaction (PCR) conditions were as follows: 5 min at 94°C, followed by 35 cycles of 1 min denaturation at 94°C, 1 min annealing at 60°C, and 1 min extension at 72°C; and for GAPDH, 5′-TCA CCA TCT TCC AGG AGC GA-3′ and 5′-CAC AAT GCC GAA GTG GTC GT-3′. PCR conditions were as follows: 5 min at 94°C, followed by 30 cycles of 30 s denaturation at 95°C, 30 s annealing at 58°C, and 30 s extension at 72°C. PCR products were confirmed on a 1% agarose gel and visualized under UV light after ethidium bromide staining.
Western blot analysis
Cells were lysed in lysis buffer (20 mM Tris [pH 7.4], 250 mM NaCl, 2 mM EDTA [pH 8.0], 0.1% Triton X-100, 0.01 mg/mL aprotinin, 0.003 mg/mL leupeptin, 0.4 mM phenylmethylsulfonyl fluoride, and 4 mM NaVO4). Lysates were then centrifuged at 14,000 g at 4°C for 10 min, and supernatants were in parallel lanes on 10% SDS gels. After electrophoresis, the proteins were electro-transferred to a nitrocellulose membrane, blocked with 5% nonfat milk, and probed with antibodies (1:1000) overnight at 4°C. The blot was washed, exposed to HRP-conjugated secondary antibodies for 1 h, and examined by enhanced chemiluminescence (ECL; Amersham, Piscataway, NJ, USA).
Combination studies
U266 cells were seeded and cotreated with HC (30 μM) and bortezomib (10, 25, and 50 nM) for a 24 h. After treatment, cells were collected and washed with chilled PBS, and the treated samples were examined using an MTT assay and Western blotting.
Statistical analysis
The results were expressed as means±SD, and an analysis of variance (ANOVA) with Bonferroni's test was used for the statistical analysis of multiple comparisons of data.
Results
HC induces apoptosis in human MM cells
We first investigated the cytotoxicity of HC and DHC (Fig. 1A) against U266 and PBMCs using the MTT assay. Under similar conditions, HC exerted significant cytotoxic effects as compared with DHC in U266 cells, but not PBMC (Fig. 1B). We next examined apoptosis inducing effects of HC and DHC using the cell cycle analysis, which detects accumulation of the subG1 portion. U266 cells were treated with 0, 20, and 40 μM of HC and DHC for 24 h. When compared with DHC, HC induced substantial accumulation of subG1 in a concentration-dependent manner (Fig. 1C). To determine whether the HC induces early and late apoptosis, we also performed annexin V and TUNEL assays. All of these results indicated that HC treatment can clearly induce apoptosis more significantly than DHC (Fig. 1D, E).
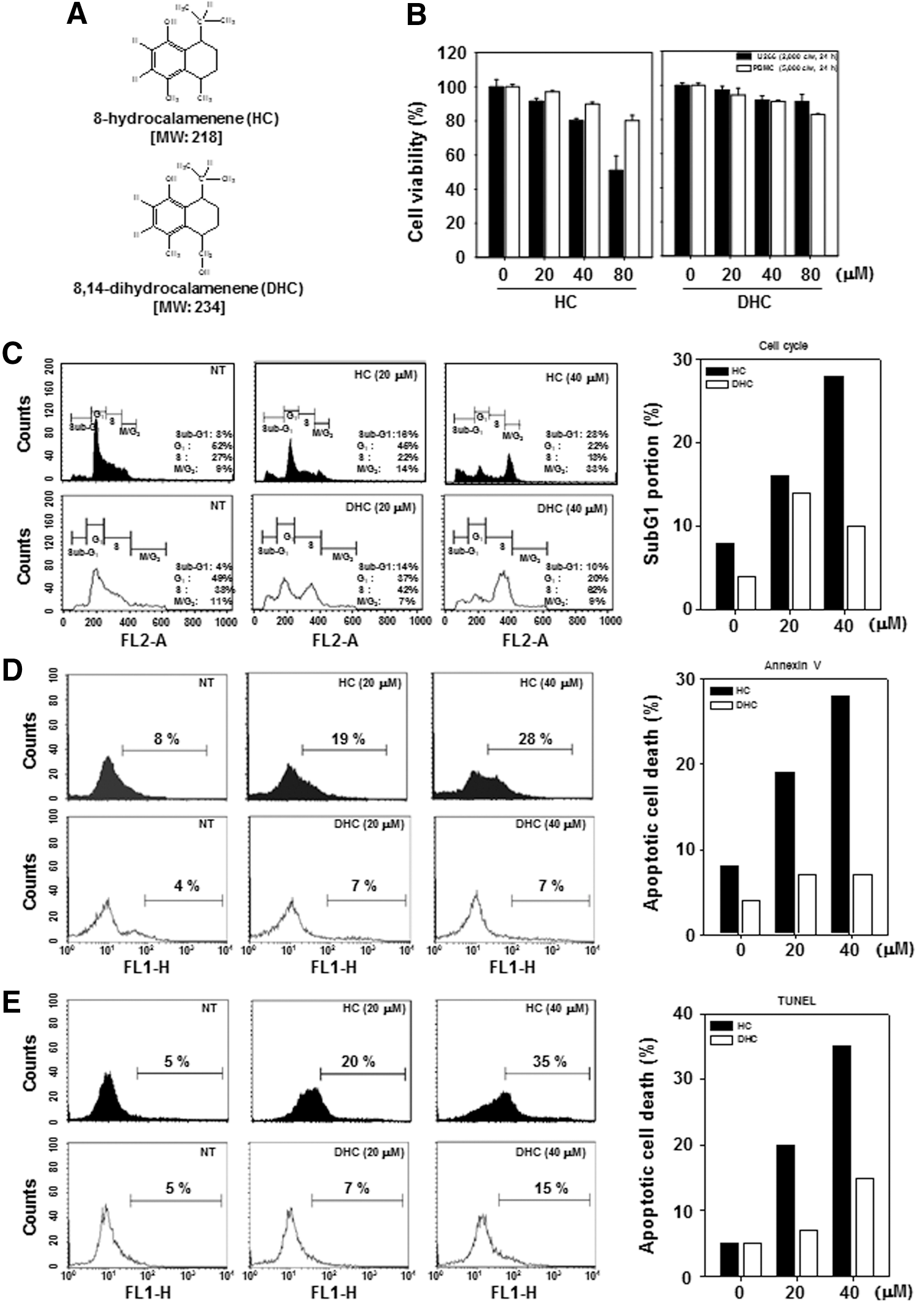
HC induces apoptosis in multiple myeloma cells but not DHC.
HC induces PARP cleavage, but not DHC
In order to compare the apoptotic effects of two sesquiterpene derivatives, HC induced substantial PARP cleavage in comparison to DHC (Fig. 2A). As a result, HC clearly exhibited more significant apoptosis-inducing activities as compared with DHC. Thus, we focused on HC to elucidate the detailed molecular mechanism underlying its antiproliferative and apoptotic effects in the MM cells.

HC down-regulates the expression of various gene products involved in antiapoptosis, metastasis, proliferation, and induces apoptosis through the activation of caspase-3, -8, and -9.
HC down-regulates the expression of antiapoptotic, metastatic, and proliferative proteins
We investigated the effect of HC on expression of the antiapoptotic proteins Bcl-2 and Bcl-xL. In addition, we examined whether suppression of metastasis and proliferation of U266 cells by HC is due to down-regulation of proteins involved in cell metastasis and proliferation, such as MMP-9, cyclin D1, and cdc2. Results indicate that HC can inhibit the expression of these proteins in U266 cells in a time-dependent manner (Fig. 2B).
HC induces the expression of p21 in U266 cells
The expression of p21 is controlled by the tumor suppressor protein p53. 14 The cyclin-dependent kinase inhibitor p21 is a prototypical member of the Cip/Kip family of cyclin-dependent kinase inhibitors. It negatively modulates cell cycle progression by inhibiting the activities of cyclin E/CDK2 and cyclin D/CDK4 complexes and can block DNA replication by binding to proliferating cell nuclear antigen. 15 We found that HC induced the expression of p21 in a concentration-dependent manner in MM cells (Fig. 2B).
HC induces caspases activation and PARP cleavage
Activation of caspases is a hallmark of apoptosis. While caspase-8 activation constitutes the extrinsic pathway, the intrinsic pathway also requires caspase-9 activation. Activated caspase-8 and -9 activate caspase-3, which, in turn, induces PARP cleavage. HC induced activation of caspase-3, -8, and -9 and PARP cleavage in U266 cells. These results indicate that HC can induce apoptosis in U266 cells through the activation of both the extrinsic and intrinsic pathways (Fig. 2C, D).
HC inhibits STAT3 signaling pathway
The modulatory effect of HC in constitutive STAT3 in U266 cells was investigated. First, U266 cells were incubated with 40 μM of HC and DHC for 8 h. The phosphorylation of STAT3 was then examined by Western blot analysis. HC inhibited the constitutive activation of STAT3 in U266 cells in comparison with DHC (Fig. 3A). HC suppressed the binding of STAT3 to DNA in a concentration-dependent manner in U266 cells by electrophoretic mobility shift assay (Fig. 3B). In addition, tyrosine kinases of the JAK family and Src have been associated with STAT3 activation. We also examined the effects of HC on the phosphorylation of JAK1, JAK2, and Src in U266 cells. The results showed that HC inhibited constitutive STAT3 activation through blocking upstream JAK1, JAK2, and Src (Fig. 3C–E).

HC suppresses STAT3 signaling pathway.
HC up-regulates the expression of PIAS3
To elucidate the mechanism by which HC inhibits STAT3 phosphorylation, we investigated the effect of HC on the negative regulators of STAT3. PIAS3 functions as a negative regulator of STAT3 signaling by interfering with its ability to bind to DNA. PIAS1 and PIAS3 protein levels were examined by Western blotting in the MM cells. Treatment of U266 cells with HC for 8 h up-regulated the expression of PIAS3 protein. However, the expression of PIAS was not affected by HC (Fig. 4A).
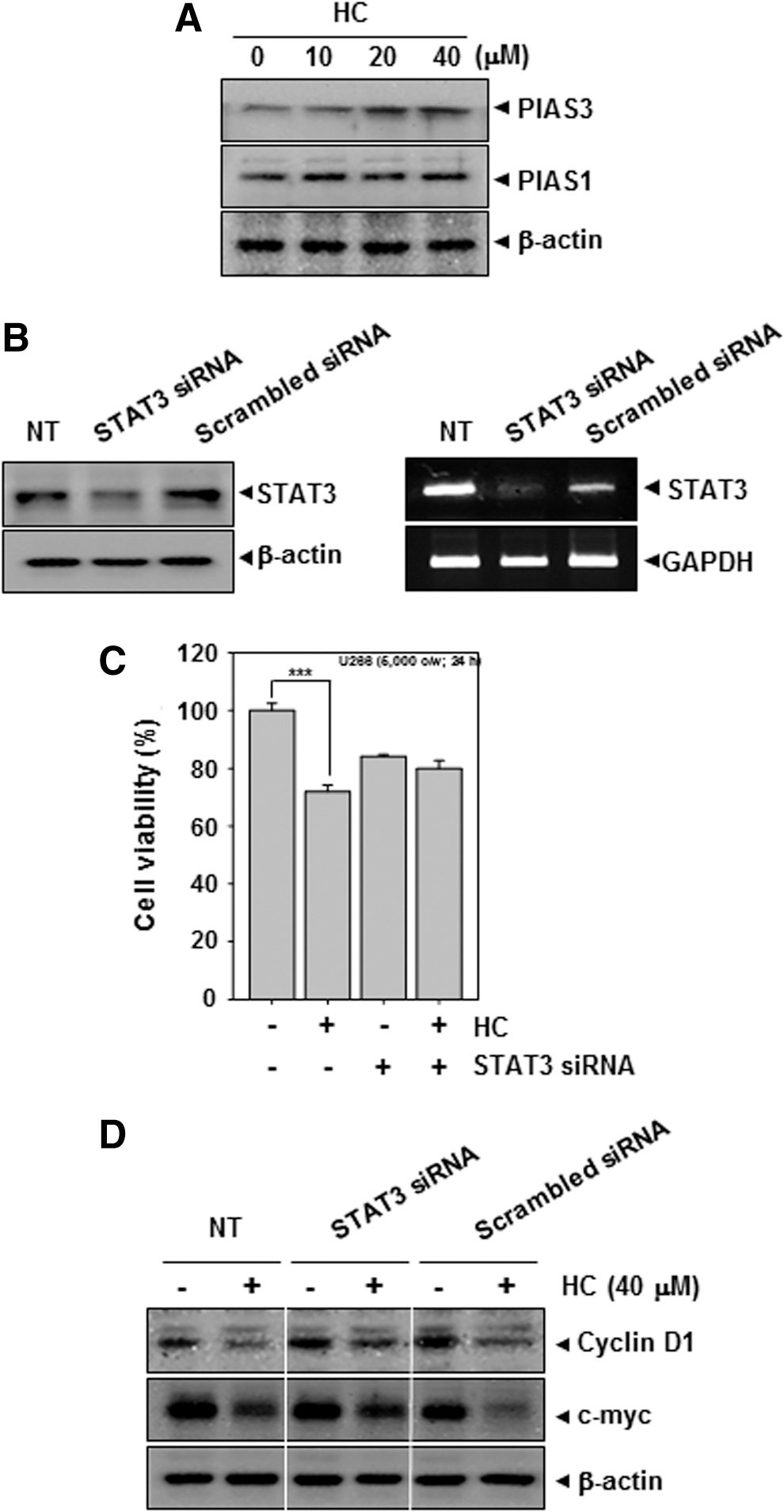
HC up-regulates PIAS3 and deletion of STAT3 does not reduce the antiproliferative effect of HC in U266 cells.
Knockdown of STAT3 by RNAi reverses the observed antiproliferative effect of HC in U266 cells
To determine whether deletion of STAT3 protein reverses the observed antiproliferative effect of HC in the cells, we knocked down STAT3 expression using RNA interference. We found that STAT3 protein and its mRNA expression were knocked down by STAT3 siRNA (Fig. 4B). However, in cells depleted of STAT3, HC failed to induce cytotoxicity (Fig. 4C) and caused the down-regulation of cyclin D1 and c-myc as compared with only HC-treated cells (Fig. 4D).
HC synergizes the cytotoxic and apoptotic effects of chemotherapeutic agents
Bortezomib has been approved for the treatment of patients with MM. We next evaluated whether HC can synergize the effects of bortezomib in MM cells. HC and bortezomib synergistically acted in inducing cytotoxicity (Fig. 5A), inhibition of STAT3 activation in U266 cells compared with each individual treatment (Fig. 5B), substantial cleavage of PARP, suppression of both antiapoptotic (IAP-1) and antiproliferative (cyclin D1) proteins, and inhibition of MDR protein as compared with individual drug treatment (Fig. 5C).
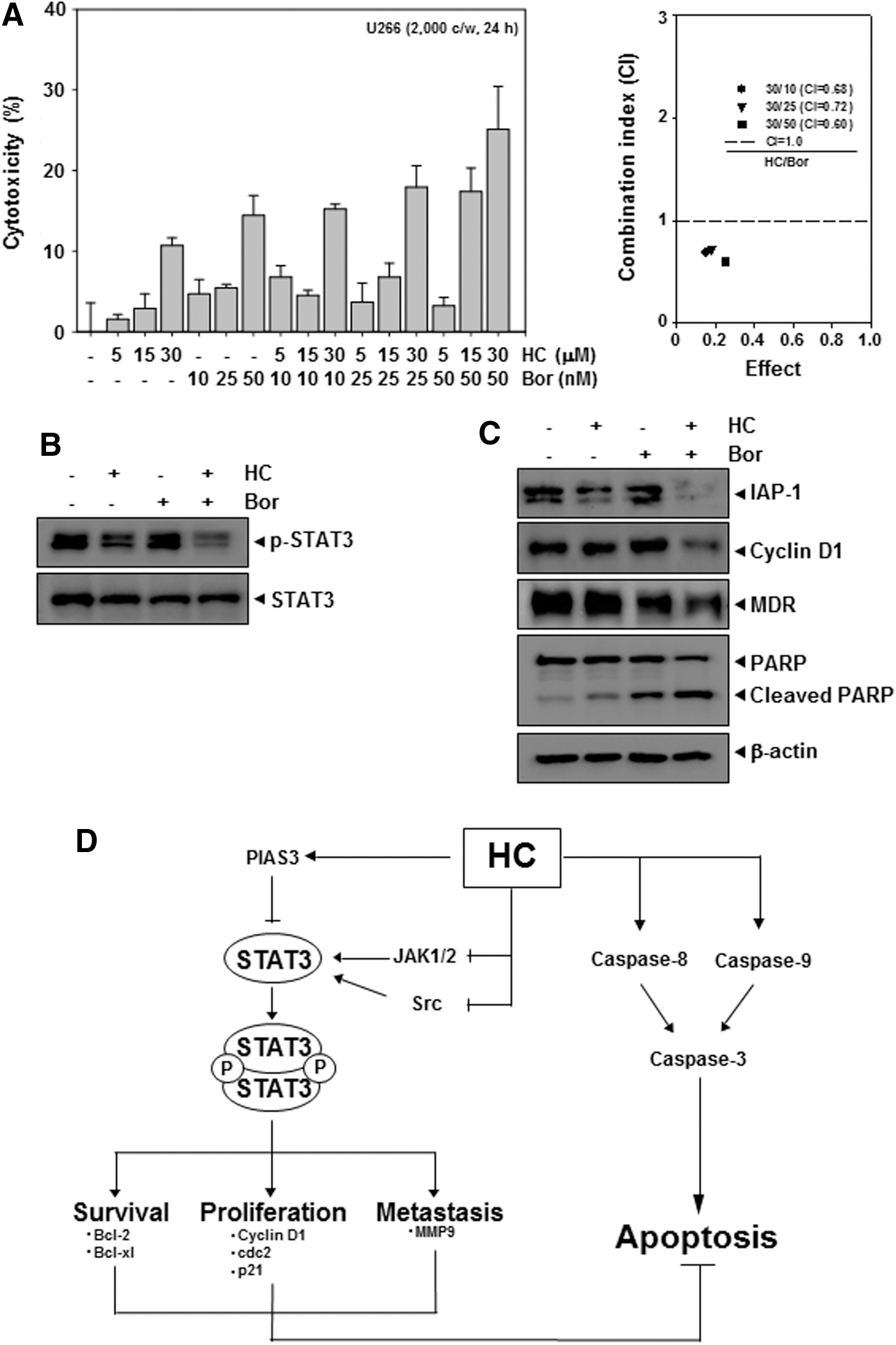
HC synergizes the cytotoxic and apoptotic effects of chemotherapeutic agents in U266 cells.
Discussion
Although R. elliptica has been demonstrated to possess antineuraminidase and anti-inflammatory activities; what bioactive compounds from the plant can inhibit proliferation and promote apoptosis of cancer cells is not fully understood. The purpose of our study was to investigate the effects of HC and DHC, derived from the plant, on the STAT3 signaling pathway, gene products, and potentiation of bortezomib-induced apoptosis in MM cells. First, we found that HC but not DHC exerted cytotoxicity on U266 cells in a dose-dependent manner, down-regulated the expression of several gene products, including Bcl-2, Bcl-xL, MMP-9, and cyclin D1, and induced apoptosis through the activation of caspase-8, -9, and -3. In addition, HC suppressed constitutive STAT3 activation in parallel with the inhibition of JAK1/2 and Src activation. Moreover, this sesquiterpene was found to synergize bortezomib-induced cytotoxicity and to significantly potentiate the apoptotic effects of bortezomib in U266 cells, suggesting that the combination treatment can be more effective in cancer treatment as compared with single agents.
The hexane fraction of R. elliptica has been previously reported to suppress LPS-induced inflammatory markers by blocking NF-κB activation in murine macrophages. 2 Whether the suppression of STAT3 activation by the plant is associated with the inhibition of NF-κB remains unclear. However, both NF-κB and STAT3 have been found to be activated in response to LPS in macrophages, 16,17 and the p65 subunit of NF-κB has been shown to interact with STAT3. 18 Thus, these findings indicate that this agent can be a good candidate for overcoming chemoresistance due to its potential effect on multiple targets involved in cancer initiation and progression.
Constitutive activation of STAT3 has been reported in a wide variety of tumors such as breast cancer, prostate cancer, 19 lymphomas, and leukemias. 11 We found, for the first time, that HC but not DHC suppressed the phosphorylation of STAT3 and the DNA binding activities of STAT3. This suppression might lead to the exhibition of cytotoxicity and the induction of apoptosis in MM cells, because constitutively active STAT3 plays an important role in the transformation and proliferation of tumor cells. 12 In addition, the effects of HC on STAT3 phosphorylation are linked with the suppression of upstream protein kinases such as JAK1/2 and Src. We found that HC repressed the constitutive activation of JAK1/2 and Src. Based on these findings, we observed for the first time that HC inhibited the constitutive active STAT3 through the suppression of JAK1/2 and Src in MM cells. All Src-transformed cells have persistently activated STAT3 and dominant-negative STAT3 blocks transformation. 20,21
Importantly, we also noticed that the expression of cyclin D1 was down-modulated by HC. Indeed, this agent led to cell cycle arrest in the subG1 phase and apoptosis selectively in U266 cells. We also found that the expression of MMP-9, which has been implicated in metastasis, was also suppressed by HC. We further observed that HC induced the down-regulation of cdc2 and caused the up-regulation of p21 proteins in the cells. Our results strongly suggest that HC can suppress proliferation and metastasis in MM cells.
It has been reported that constitutive STAT3 activation is involved in the growth promotion and apoptosis inhibition functions which can result in chemoresistance in tumor cells. 11,22 The antiapoptotic proteins, Bcl-2 and Bcl-xL, which are regulated by STAT3, are overexpressed in MM cells and can also lead to resistance to apoptosis by various chemotherapeutic agents, in parallel with an increase in chemoresistance. 23 We found, for the first time, the down-regulation of various antiapoptotic gene products, such as Bcl-2 and Bcl-xL by HC. We further observed that HC can significantly synergize the cytotoxic effects of bortezomib and potentiate bortezomib-induced apoptosis via the suppression of STAT3 signaling in human MM cells. When investigated for the mechanism of this potentiation, we observed the down-regulation of various gene products, such as IAP-1, cyclin D1,and MDR by drug combination. Our results clearly suggest that HC is capable of down-regulating the expression of several genes involved in cell survival and chemoresistance. Thus, HC can act as a supplementary therapy to overcome the resistance and limit serious adverse effects encountered frequently in cancer treatment with mono-targeted agents.
Taken together, our data demonstrate that HC is an effective blocker of the STAT3 signaling pathway and, thus, may have potential for the prevention and treatment of MM. However, further animal studies are needed with HC alone and in combination with standard chemotherapeutic agents to demonstrate the potential applications of HC.
Footnotes
Acknowledgments
This work was supported by a grant from Kyung Hee University in 2011 (KHU-20111736). This work was also supported by the Korea Science and Engineering Foundation (KOSEF) grant funded by the Korean Ministry of Education, Science, and Technology (MoEST) (No. 2011-0006220).
Author Disclosure Statement
The authors have no conflicts of interest to disclose.