
Abstract
Glycerol-3-phosphate acyltransferase (GPAT) catalyzes the first committed step in triacylglycerol (TAG) and phospholipid biosynthesis, and has been considered as one of the drug targets for treating hepatic steatosis, insulin resistance, and other metabolic disorders. The aim of this study was to investigate the GPAT inhibitors from natural products and to evaluate their effects. The methanol extract of Aralia cordata roots showed a strong inhibitory effect on the human GPAT1 activity. A further bioactivity-guided approach led to the isolation of ent-pimara-8(14),15-dien-19-oic acid, (PA), one of the major compounds of A. cordata, which suppressed the GPAT1 activity with IC50 value of 60.5 μM. PA markedly reduced de novo lysophosphatidic acid synthesis through inhibition of GPAT activity and therefore significantly decreased synthesis of TAG in the HepG2 cells. These results suggest that PA as well as A. cordata root extract could be beneficial in controlling lipid metabolism.
Introduction
B
There are two major pathways for TAG synthesis; the glycerol phosphate pathway involved in de novo lipogenesis and the monoacylglycerol pathway, which plays a major role in lipid absorption. The first committed rate-limiting step in the glycerol phosphate pathway is catalyzed by glycerol-3-phosphate acyltransferase (GPAT). Currently, four mammalian GPAT isoforms have been identified—two mitochondrial GPAT (GPAT1 and GPAT2) 4,5 and two microsomal GPAT (GPAT3 and GPAT4), each encoded by separate genes. 6 –8 The activity of GPAT1 can be differentiated from other isoforms by their resistance to sulfhydryl group reactive reagents such as N-ethylmaleimide (NEM). 9 GPAT1 is most highly expressed in the liver and adipose, and accounts for 30–50% of the total GPAT activity in the liver. 10 GPAT1 is regulated at both the transcriptional and post-transcriptional levels. GPAT1 mRNA is upregulated by a high carbohydrate, insulin, and sterol regulatory element binding protein-1 (SREBP-1) 11 and downregulated by AMP-activated protein kinase (AMPK). 12 The activation of GPAT1 is closely related with the requirements for de novo TAG synthesis, suggesting a role for this enzyme in regulating TAG biosynthesis.
TAG synthesis is catalyzed sequentially by enzymes that have multiple isoforms, including GPAT, 1-acylglycerol-3-phosphate acyltransferase (AGPAT), and diacylglycerol acyltransferase (DGAT). The inhibition of these enzymes using genetic alterations results in decreased TAG in various tissues. 13 –17 Noteworthy, the previous studies have reported that mice with genetically modified GPAT in the liver 14,18,19 are closely developing hepatic steatosis, suggesting that the GPAT enzyme can be the therapeutic target. Recently, a couple of GPAT inhibitors have been reported. 20,21 However, the pharmacological validation of their use in cells and animal models remains to be examined.
Aralia cordata Thunb. belongs to the family Araliaceae, which has long been recognized in Korea, China, and Japan as therapeutic herbs with antinociceptive, 22 antidementia, 23 antioxidant, 24 anticancer, 25,26 and anti-inflammatory 27 activities. The root of this plant has been used to treat rheumatism, lumbago, common cold, migraines, and lameness clinically. A previous study showed that the dichloromethane fraction of the root contains essential oil, saponins, sesquiterpenes and diterpene acids, diterpenes, polyacetylenes, and sterols. 28 During the screening of human GPAT1 inhibitors from natural sources, we found that the MeOH extract from the root of A. cordata strongly inhibited the GPAT1 enzymatic activity. The further bioactivity-guided approach led to the isolation of the diterpene compound, ent-pimara-8(14),15-dien-19-oic acid (PA). In this study, we isolated and identified a bioactive molecule from A. cordata, which affected the GPAT1 activity and TAG metabolism.
Materials and Methods
Materials
Optical rotations were determined on a JASCO P-1020 Polarimeter using a 100-mm glass microcell. 1 H-NMR (300 MHz) and 13 C-NMR (100 MHz) spectra were obtained on a Varian Unity Inova 400 spectrometer using CDCl3 as the solvent. Electrospray ionization (ESI) mass spectrometry (MS) was measured on a Finnigan navigator spectrometer. Stationary phases for column chromatography (Kieselgel 60, 70–230, and 230–400 mesh, and Lichroprep RP-18 gel, 40–63 μm; Merck) and TLC plates (Kieselgel 60 F254 and RP-18 F254; Merck) were purchased from Merck. Bovine serum albumin and palmitotyl-CoA were obtained from Sigma Chemical Co. [1- 14 C] acetic acid (250 μCi) was purchased from Amersham Biosciences. [1- 14 C] glycerol (50 μCi) and [1- 14 C] oleate (50 μCi) were purchased from PerkinElmer.
Extraction and isolation
The dried roots of A. cordata (3 kg) were purchased from a herbalist in Daejeon (Korea) and extracted with MeOH by marceration for 15 days at room temperature to give 373 g of dried MeOH extract, which was suspended in 1 L of water and then extracted with an equal volume of CHCl3. A part of the CHCl3-soluble fraction (50 g) was separated by silica gel column chromatography (9.5 cm diameter×35.0 cm, 2 kg, 70–230 mesh; Merck) using a stepwise gradient of hexane: EtOAc (10:1–1:1), which afforded 11 subfractions (C1–C11). The fraction C3 (3 g) was rechromatographed over a silica column chromatography (5 cm diameter×120 cm; 230–400 mesh; eluting solvent: 100% hexane) to yield crude PA, which was recrystallized from MeOH yielding a diterpene compound, PA (1.3 g). The compound was sealed and stored in a dark place at −20°C.
ent-pimara-8(14),15-diene-19-oic acid: C20H30O2, colorless needle (MeOH), ESI-MS: m/z 301 [M-H]+, 357 [M+Na]+; UV (MeOH); λmax 203 nm, mp: 165–166 C;
Human GPAT1 expression in Sf-9 cells and membrane preparation
Human GPAT1 genes were overexpressed in Sf-9 insect cells using the Bac-to-Bac Baculovirus Expression System (Invitrogen) according to the manufacturer's instructions. hGPAT1 (gene accession number: BC030783) cDNA clone is a gift from 21C Human Gene Bank, Genome Research Center, KRIBB, Korea. The cDNA with an in-frame C-terminal FLAG tag was cloned into the pFastBac1 donor vector (Invitrogen) to obtain the recombinant bacmid. Sf-9 cells infected with the resultant baculovirus at 10 MOI for 56 h were harvested in ice-cold PBS and homogenized using a sonicator (Fisher Scientific) in the sucrose solution (250 mM sucrose, 10 mM Tris, pH 7.4, 1 mM EDTA, and 1 mM DTT). Nuclei were removed by centrifuging at 600 g for 15 min at 4°C. The resulting supernatants were further centrifuged at 8000 g for 15 min at 4°C to collect crude mitochondria. The pellets were resuspended and the protein concentration was quantified using the Bradford protein assay method.
The GPAT activity was measured according to the method of Hammond et al. 4 The assay buffer contained 75 mM Tris, pH 7.5, 4 mM MgCl2, 8 mM NaF, 100 μM palmitoyl-CoA, 1.8 μM [ 14 C] glycerol-3-phosphate, and 2 mg/mL BSA. To measure the NEM-resistant GPAT activity, protein samples were mixed with NEM (2 mM final concentration) for 15 min at 4°C before starting the reaction. The reaction was started by mixing 2 μg of mitochondrial membrane fraction with the assay buffer in a final volume of 200 μL and shaking the samples for 20 min at 26°C. The reaction was stopped by adding 1 mL of water-saturated butanol and then adding 0.5 mL of butanol-saturated water to extract lysophosphatidic acid (LPA). After vortexing, 0.8 mL of the organic phase was transferred to a tube and washed once with 0.8 mL of butanol-saturated water. The radioactivity of the top phase was determined by liquid scintillation counting (Tri-Carb 2900TR Liquid Scintillation Analyzer; PerkinElmer).
Cell culture and viability test
Human hepatocyte HepG2 cells were obtained from the American Type Culture Collection. Cells were maintained in the DMEM (Invitrogen) supplemented with 10% heat-inactivated FBS and 1% penicillin–streptomycin (Sigma) in 5% CO2 at 37°C. To test whether the PA has cytotoxic effects on HepG2 cells, we determined viability by the MTT assay. Cells were seeded in 96-well plates (2×104 cells/well) and treated with compounds for 24 h. The absorbance of the formazan dye was read using the ELISA plate reader at 560 nm.
Cellular TAG and LPA formation assay using HepG2 cells
The cell-based assay for confirmation of GPAT inhibition was performed in HepG2 cells. To determine the de novo synthesis of LPA and TAG, the cells were cotreated with PA or vehicle at the indicated concentrations and [ 14 C] glycerol (0.5 μCi) or [ 14 C] acetate (0.5 μCi) or [ 14 C] acetate (0.5 μCi). At the end of the incubation, intracellular lipids were extracted with a mixture of hexane/isopropanol (3:2, v/v). Cellular lipids were resolved on silica plates by thin layer chromatography (Kieselgel 60 F254 plates; Merck) using either a solvent system consisting of hexane/diethyl ether/acetic acid (80:20:1, v/v) for TAG or chloroform/ethanol/water/triethylamine (30:35:6:35, v/v/v/v) for LPA. The isotope-labeled lipids were detected and quantified with a bioimage analyzer (FLA-7000; Fuji).
Statistical analysis
All data are presented as mean±standard deviation (SD). Statistical analysis was performed using the Student's t-test (unpaired) with GraphPad Prism 4 software. A P value of<.05 was considered to be significant.
Results
MeOH extract and its fractions of A. cordata inhibit human GPAT1 activity and intracellular TAG synthesis
The crude MeOH extract (AC-M) showed considerable inhibition of the human GPAT1 activity with an IC50 value of 19.7 μg/mL by confirming the in vitro enzymatic activity assay. The extract was separated into two portions with a CHCl3-soluble part (AC-C) and remaining water residue (AC-H). The AC-C exhibited more potent GPAT1 inhibitory effects with an IC50 value of 19.4 μg/mL, suggesting that putative bioactive molecules were transferred into the CHCl3 phase (Fig. 1A). To examine whether AC-M and AC-C can inhibit newly synthesized TAG through the inhibition of GPAT1, we analyzed isotope-labeled TAG contents after treatment of [ 14 C] acetate or [ 14 C] glycerol as a substrate in the presence or absence of compounds in HepG2 cells. First, cell viability assay was performed to avoid any toxic effects on cells using the MTT assay. The treatment of both fractions (AC-M and AC-C) up to 20 μg/mL for 24 h had no effect on the viability of HepG2 cells (Fig. 1B). TLC analyses revealed that the AC-M and AC-C efficiently inhibit incorporation of TAG from both acetates and glycerol in a dose-dependent manner (Fig. 1C, D). Moreover, these data showed that AC-M and AC-C have similar or better efficacy compared with berberine (BBR, a positive control, 15 μg/mL) at the same concentration. A previous study showed that BBR inhibits TAG synthesis through AMPK activation. 29
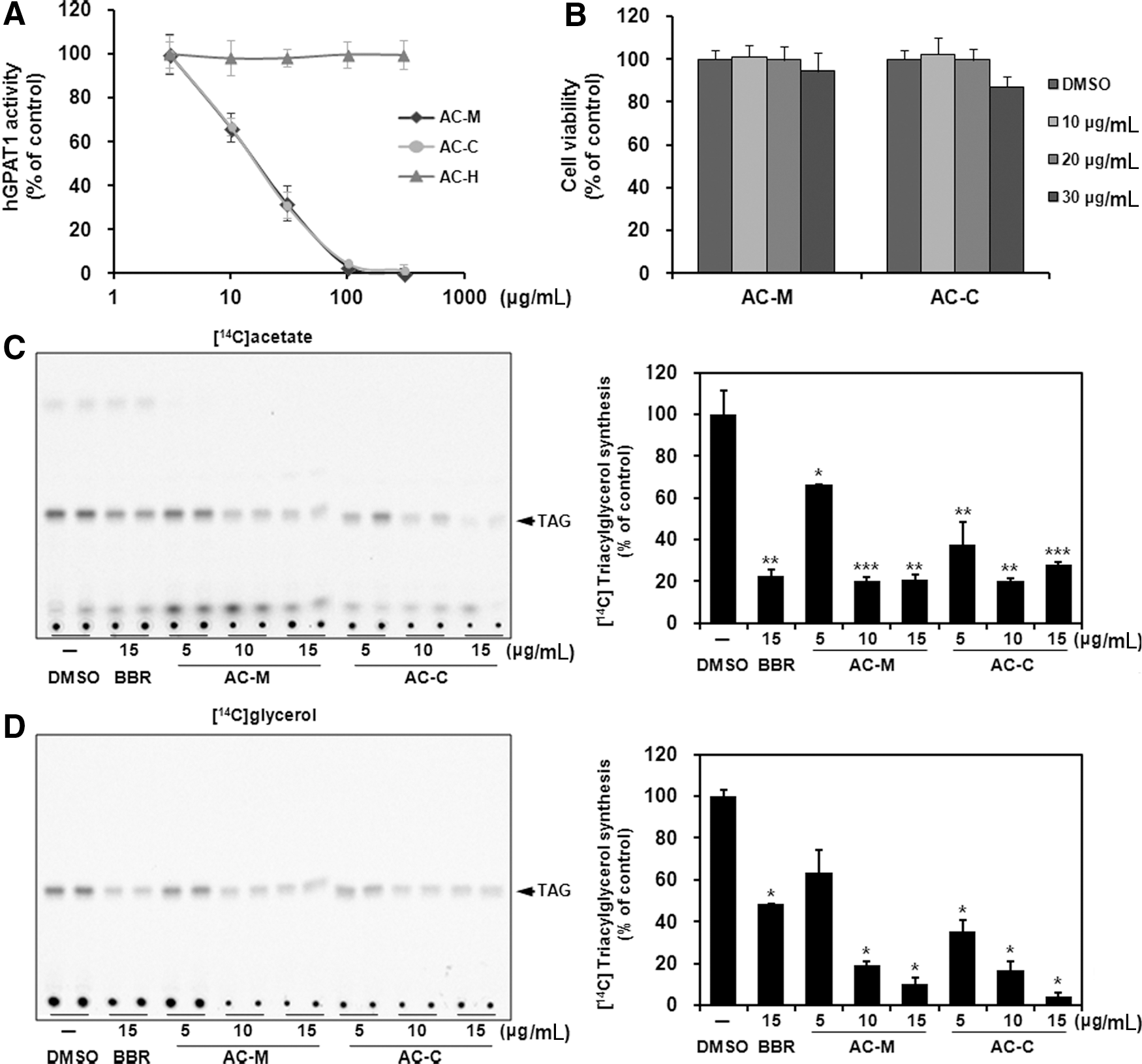
Effect of MeOH extract from Aralia cordata and its CHCl3 and H2O phases on GPAT1 activity and de novo triacylglycerol (TAG) synthesis.
Isolation of PA as an hGPAT1 inhibitory compound
Further bioactivity-guided fractionation of the AC-C led to the isolation of an active substance with appreciable GPAT1 inhibitory activity. The structure of the isolate was identified as one of the pimarane-type diterpenes, PA by direct comparison of its physical and spectral properties (ESI-MS, 1 H-NMR, and 13 C-NMR) with reference data (Fig. 2A). PA was one of the major compounds of the MeOH extract (AC-M) of A. cordata roots with a yield of 0.62% w/w and occupied 2.6% w/w of chloroform fraction (AC-C). To evaluate the activity of PA against GPAT1, we performed in vitro GPAT1 enzymatic assay using the mitochondrial membrane fraction prepared from human GPAT1-overexpressed Sf-9 cells. PA showed considerable inhibitory efficacy against GPAT1 with an IC50 value of 60.5 μM (Fig. 2B).

Activity-guided isolation of ent-pimara-8(14),15-dien-19-oic acid (PA) from the roots of A. cordata and its hGPAT1-inhibitory activity.
To determine whether PA exhibits multifunctional effects on other acyltransferases, we examined its inhibitory effects on human DGAT1 (acyl CoA:diacylglycerol acyltransferase-1), which is well known to catalyze the esterification of sn-1,2-diacylglycerol with long-chain fatty acyl CoA in the final step in TAG synthesis. As a result, PA exhibited weak DGAT1 inhibition with an IC50 value of 193 μM (data not shown), suggesting it was more specific against hGPAT1 than hDGAT1.
PA inhibits endogenous GPAT
Because PA is the main isolate from the AC-C, we next assessed the ability of PA to inhibit the endogenous GPAT activity by measuring its product LPA in HepG2 cells. In the intracellular condition, the glycerol could be metabolized and sequentially converted into glycerol-3-phosphate, LPA, phosphatidic acid, and TAG through the glycerol phosphate pathway. When incubated with 14 C-labeled glycerol to estimate de novo LPA synthesis, cells are expected to show increased generation of 14 C-labeled LPA, which was attenuated by treatment of PA in a dose-dependent manner (Fig. 3A, B).

Effect of PA on lysophosphatidic acid (LPA) formation in HepG2 cells.
PA blocks intracellular TAG synthesis in HepG2 cells
Finally, we examined whether PA inhibits de novo intracellular TAG synthesis by suppressing the GPAT1 activity. TAG synthesis was determined using [ 14 C] acetate or [ 14 C] glycerol incorporation, whereas TAG assembly was assessed by the incorporation of [ 14 C] oleate in HepG2 cells. Cells were incubated with 0.5 μCi [ 14 C] acetate or [ 14 C] glycerol for 6 h and [ 14 C] oleate for 2 h in the presence of PA (10∼30 μM) or BBR (15 μg/mL [40.3 μM], a positive control). As observed in Figure 4A, 20 and 30 μM PA inhibited [ 14 C] acetate incorporation into TAG by 21.5% and 72.1%, respectively. At the same time, PA suppressed [ 14 C] glycerol incorporation into TAG by 24.1% and 79% at 20 and 30 μM, respectively (Fig. 4B). Moreover, PA abolished incorporation of [ 14 C] oleate to TAG by 57.1% and 39.4% at 20 and 30 μM (Fig. 4C), suggesting that TAG assembly was also affected during the treatment. These findings concluded that PA could inhibit de novo TAG synthesis through the regulation of GPAT1 positioning on the first step in the glycerol phosphate pathway.
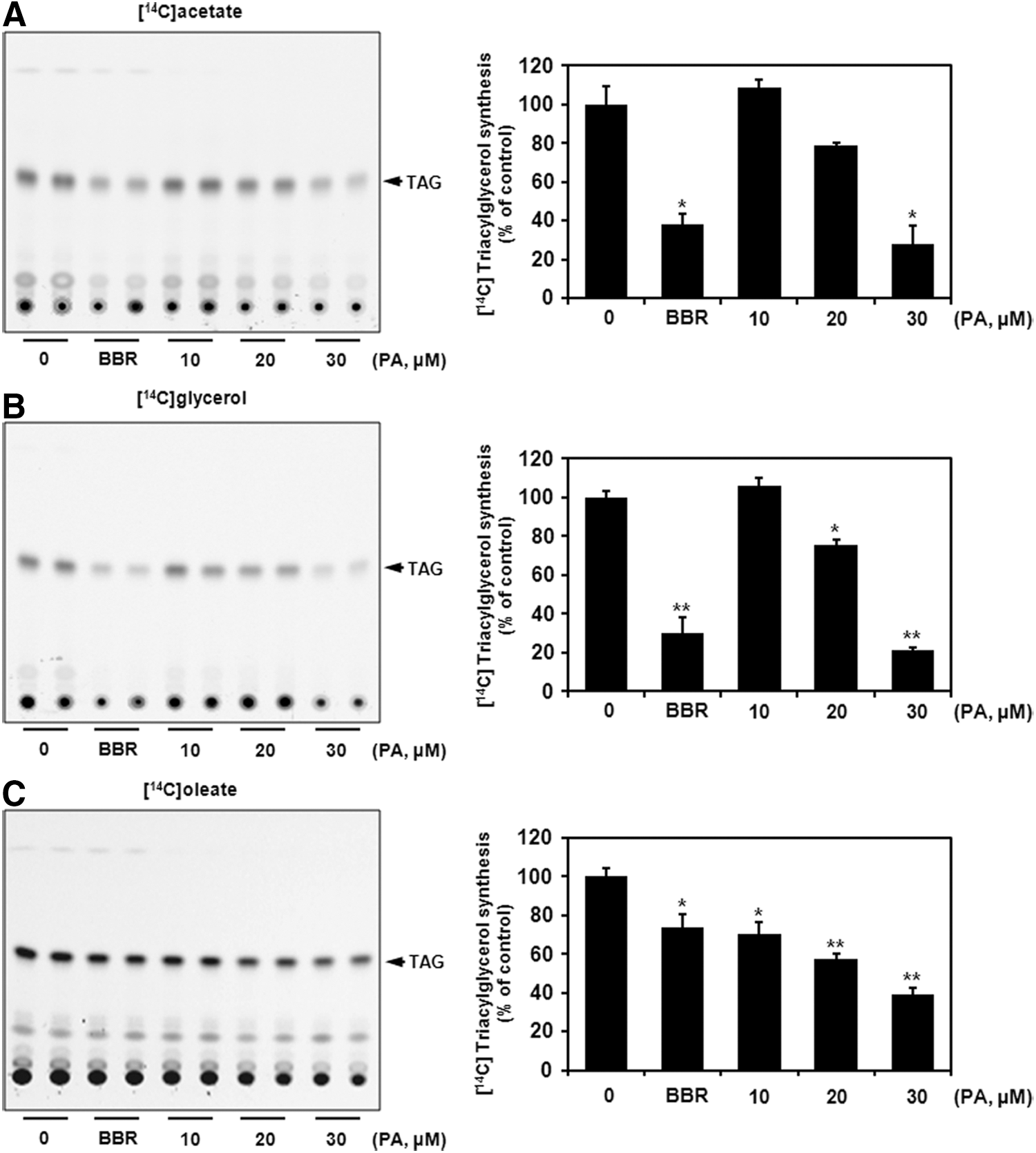
Effect of PA on de novo intracellular TAG synthesis in HepG2 cells. Lipid profile analyzed by TLC using either [14C] acetate
Discussion
Previous studies revealed that overexpression of GPAT1 in hepatocytes increased TAG synthesis rather than phospholipids. 30 In addition, liver-directed overexpression of GPAT1 in lean mice and rats resulted in hepatic steatosis and hepatic insulin resistance through the increase of TAG secretion. 31 Conversely, selective hepatic GPAT1 knockout in ob/ob mice resulted in decreased hepatic steatosis, and improved plasma glucose and cholesterol levels. 13,32 Thus, inhibiting GPAT1 may be an attractive way to interfere with hepatic TAG accumulation and related diseases. 19
Recently, the inhibition of lipogenesis was shown to increase fatty acid oxidation. 20,33 Carnitine palmitoyltransferase-1 (CPT-1), the rate-limiting enzyme of the mitochondrial fatty acid β-oxidation, and GPAT1 are both located on the outer mitochondrial membrane and compete for long-chain acyl-CoAs as their substrate. Genetic or pharmacological GPAT inhibition resulted in an increase in oxidation because the acyl-CoA substrates were partitioned toward CPT-1 for β-oxidation rather than the TAG synthetic pathway. 20 Conversely, GPAT1 overexpression in rat hepatocytes resulted in a dramatic decrease in fatty acid oxidation and a significant increase in hepatic diacylglycerol and phospholipid biosynthesis. 30 Therefore, further studies are needed to investigate the effect of PA on fatty acid β-oxidation and alteration of related gene profile.
Previous studies reported that PA has anti-inflammatory 34 and vasorelaxant effects, 35 however, there has been no comprehensive research on the function of PA in TAG metabolism. Therefore, this is the first time report of PA on downregulation of TAG synthesis in human HepG2 cells.
In this study, we showed that the MeOH extract and its fractions of A. cordata roots have strong inhibitory effects on human GPAT1 activity. Also, PA, which was isolated from the MeOH extract of A. cordata roots, markedly decreased new syntheses of LPA and TAG by inhibiting GPAT1 in the HepG2 cells. Therefore, these results suggest that PA as well as the extract of A. cordata roots could be beneficial in regulating lipid metabolism and useful for preventing or treating lipid metabolism disorders and their complications.
Footnotes
Acknowledgments
This work was supported by grants from the Foreign Plant Extract Library Program (FGC1621221) of the Ministry of Education, Science and Technology and from the KRIBB Research Initiative Program, Republic of Korea. This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (NRF-2013R1A1A3009382).
Author Disclosure Statement
No competing financial interests exist.