
Abstract
Diabetic encephalopathy (DE) is one of the severe complications in patients with diabetes mellitus. Paeonol, an active compound isolated from the root bark of Paeonia suffruticosa, has significant antidiabetic activity in vivo. However, its underlying beneficial effects on DE were unclear. In the present study, the protective activity of paeonol on DE was evaluated in streptozotocin (STZ)-induced diabetic rats. Paeonol at 50 and 100 mg/kg significantly increased body weight and decreased blood glucose levels, glycosylated serum proteins, and serum advanced glycation end products (AGEs) levels. Immunohistochemistry assays and Western blot analysis revealed a significant decrease in expressions on receptor for advanced glycation end products (RAGE) and nuclear factor kappa B (NF-κB) in hippocampus and cerebral cortical neurons after paeonol treatment. Furthermore, paeonol significantly increased glutathione content and remarkedly decreased induced nitric oxide synthase activity in hippocampus tissue. Our findings indicated that paeonol could improve the pathological damage of DE in STZ-induced diabetic rats. It might be associated with the modulating AGEs/RAGE/NF-κB pathway. This study suggested that paeonol might be a promising candidate for the prevention and treatment of DE.
Introduction
D
The increased morbility and mortality of encephalopathy in patients with type 2 diabetes is closely associated with hyperinsulinemia, hyperglycemia, hypertension, and hypercholesterolemia, and commonly accompanying attributes, such as obesity. 4 Advanced glycation end products (AGEs), generated by nonenzymatic glycation and oxidation of protein and reducing sugars, are known to initiate and aggravate the pathological damage in DE. 5 The accumulation of AGEs can damage brain organs by eliciting oxidative stress and inflammatory responses which have been implicated in the damage of the structure and function of cerebral cortical neuron and hippocampus. 6 AGEs activates the receptor for advanced glycation end products (RAGE) and mediates the reactive oxygen species (ROS)-dependent activation of transcription factor nuclear factor kappa B (NF-κB) and other regulatory cytokines. 7 Therefore, interfering with the AGEs/RAGE/NF-κB signaling pathway might have the potential to attenuate hyperglycemia-induced brain damage.
Paeonol [1-(2-hydroxy-4-methoxyphenyl)-ethanone] (Fig. 1) is a major component of Paeonia suffruticosa which has been used as a Chinese herbal medicine for thousands of years. Accumulating evidence in vitro and in vivo showed that paeonol has a beneficial neuroprotective activity on oxygen–glucose deprivation-induced injury in ischemia–reperfusion injured rats.
8,9
It can attenuate the neurotoxicity for the primary microglia and cortical neurons in cerebral infarct and neuro-deficit in rat, as well as hippocampus and temporal cortex in

The chemical stucture of paeonol (C9H10O3, MW=166.18).
The aim of the present investigation was to evaluate the hypoglycemic and antioxidant activities of paeonol on DE in STZ-induced rats.
Materials and Methods
Animals and treatment
Male Sprague-Dawley rats weighing 180–220 g were purchased from the Experimental Animal Center in Nantong University. Rats were randomly housed five per cage for 1 week to adapt the environment before being used for further experiments The animals were kept in a controlled environment, with a temperature of 22±1°C and a 12 h light/12 h dark cycle. Standard food and water were given ad libitum. All experiments and animal care were performed strictly in accordance with the Provision and General Recommendation of Chinese Experimental Animals Administration Legislation and approved by the Science and Technology Department of Jiangsu Province. All animals were fed high fat/high sucrose feed for 50 days and then injected intraperitoneally with a single dose of 1% STZ (Sigma Co.) 40 mg/kg 13 except the control group. The self-made feed consisted of 15% lard, 10% sucrose, 3% cholesterol, 3% NaCl, and 69% commercial feed. Rats in Group A were fed with non-high fat/high sucrose standard feed and received intraperitoneal (i.p.) injection of 0.9% saline for blank control. After STZ injection for 7 days, 12 h fasting blood glucose levels were determined by a blood sugar kit. Rats with blood glucose levels greater than 13.9 mmol/L were considered as diabetic and used for the study. Ten days after the induction of diabetes, STZ-treated rats were divided into six groups of eight animals. Group B was left without treatment and served as the model group (Model). Group C, D and E were treated with paeonol (Nanjing Zelang Pharmaceutical Co., Ltd.) at a daily dose of 25, 50, and 100 mg/kg, respectively. The daily dosages of paeonol were used in this experiment were estimated from published literature. 11 Groups included low-paeonol group (L-pae), moderate-paeonol group (M-pae), and high-paeonol group (H-pae). Rats in Group F received aminoguanidine (AG group) at a daily dose of 50 mg/kg for the positive control. 13 Each drug was administrated intragastrically for 30 days.
Measurements of body weight and blood glucose
The rats were weighed before being sacrificed and then blood samples were collected from the ophthalmic venous plexus. Serum was separated by centrifugation at 3000 g for 5 min and stored at −80°C until determination within 12 h. Rats were sacrificed by cervical dislocation. Brains were removed carefully and then the hippocampi were immediately dissected on a cold plate, weighed and homogenized with ice-cold saline. The homogenate was centrifuged at 3000 g for 10 min at 4°C. Finally, the supernatant was taken for use.
Fasting blood glucose levels were determined by a blood sugar kit (Nanjing Jiancheng Bioengineering Institute) based on the glucose oxidase method 14 according to the manufacturer's instructions. The absorbance of which is measured at 505 nm with a microplate reader (SPECTRAmax 190; Molecular Devices).
Measurement for glycosylated serum proteins in serum
Serum concentrations of glycosylated serum proteins (GSP) were detected using a commercial kit (Nanjing Jiancheng Bioengineering Institute). In brief, an aliquot of serum (100 μL) was mixed with 2 mL of nitroblue tetrazolium maintained at 37°C for 15 min. At the end of the reaction, a stabilizer was added to the mixture. Finally, the absorbance of the mixture was read at 530 nm with a spectrophotometer.
Enzyme-linked immunosorbent assay for AGEs in serum
Enzyme-linked immunosorbent assay was performed to quantity AGEs concentration in serum according to the reported method. 15 Briefly, 96-well plate was incubated with diluted serum sample (1:5) and standard AGE-bovine serum albumin (40, 80, 160, 320 and 480 ng/L) for 30 min at 37°C. An aliquot of phosphate buffered saline (PBS) was used for the blank control and then the wells were washed with a wash buffer for five times. Horseradish peroxidase (HRP)-labeled AGEs antibody (50 μL) was added to maintain for another 30 min at 37°C. After being washed five more times, the plate was incubated with 100 μL of chromogenic agent tetramethyl benzidine with a dark environment for 30 min at 37°C. The blue reaction was changed to yellow by an addition of 50 μL acidic stopping solution. Finally, the absorbance of the samples was measured with a spectrophotometer at 450 nm.
Preparation of test samples of hippocampus
The hippocampus tissue of rats was taken quickly after being sacrificed and was excised quickly under sterile conditions. The tissue was cut into pieces and then added with cold saline containing 1% Triton X-100 to prepare the homogenate of hippocampus tissue. The samples were then collected and centrifuged at 14,000 g at 4°C. The supernatant was used for the glutathione (GSH) content and induced nitric oxide synthase (iNOS) activity assays with in 6 h.
Assay of GSH content in hippocampus tissue
The GSH content in hippocampus homogenate was determined as described previously 16,17 and expressed as μmol/L.
Assay for iNOS activity in hippocampus tissue
The pathogenesis of DE is associated with oxidative stress in the body; therefore, the activity iNOS in the serum was measured. The activity of iNOS in the serum was measured by an iNOS activity assay kit according to the manufacturer's instructions (Beyotime Institute of Biotechnology) using a fluorescence microplate reader with excitation/emission wavelength of 495/515 nm.
Immunohistochemistry assays for RAGE and NF-κB
Rats were sacrificed by transcardial perfusion of 0.1 M PBS (pH 7.4). Brains of rats were fixed quickly in 4% paraformaldehyde in 0.1 M PBS. After 24 h fixation, the brains were dehydrated by serial gradient concentrations of alcohol and then embedded in paraffin blocks. Two sections (4 μm in thickness) of the following brain regions were chosen for immunohistochemistry study: −2.3 mm to −3.8 mm relative to bregma, including cerebral cortex and hippocampus. Tissue slides were stored at 4°C before being stained.
Immunohistochemistry was performed on the paraffin-embedded tissue sectons using a standard avidin-biotin complex-peroidase method. 18 Briefly, xylene was used for the deparaffinization of sections and serial gradient concentrations of alcohol for the dehydration. Then, endogenous peroxidase was blocked by the incubation with 3% hydrogen peroxide for 5 min at room temperature. After being washed with PBS (pH=7.2), the sections were blocked with 10% goat serum for 30 min at 37°C. Rabbit anti-rat antibodies specific against RAGE (Wuhan Boster Bio-Engineering Ltd.) and NF-κB p65 (Bioworld Technology, Inc.) was added and maintained at 4°C for overnight. An aliquot of PBS was used to replace the primary antibodies as the negative control. Sections were then incubated with secondary antibody biotinylated anti-rabbit immunoglobulin G and maintained with ABC Elite (HRP) reagent. 19 Finally, 3,3′-diaminobenzidine was used for chromogen. For staining for β-amyloid (Aβ) expression, Congo Red in methanol was used for the staining of the sections for 3 min. Finally, the sections were cleared in xylene before mounting with neutral gum under a cover slip and then photographed under light microscopy using Leica Application suite V3.1.0 software.
All sections were visualized and photographed under a light microscope (Olympus IX71). The images were analyzed with Image-Pro Plus software.
Biochemical assays for glutamic-oxaloacetic transaminase, glutamic-pyruvate transaminase, blood urea nitrogen and creatinine
Serum levels of glutamic-oxaloacetic transaminase, glutamic-pyruvate transaminase, blood urea nitrogen, and creatinine were carried out according to the method as described previously. 20,21
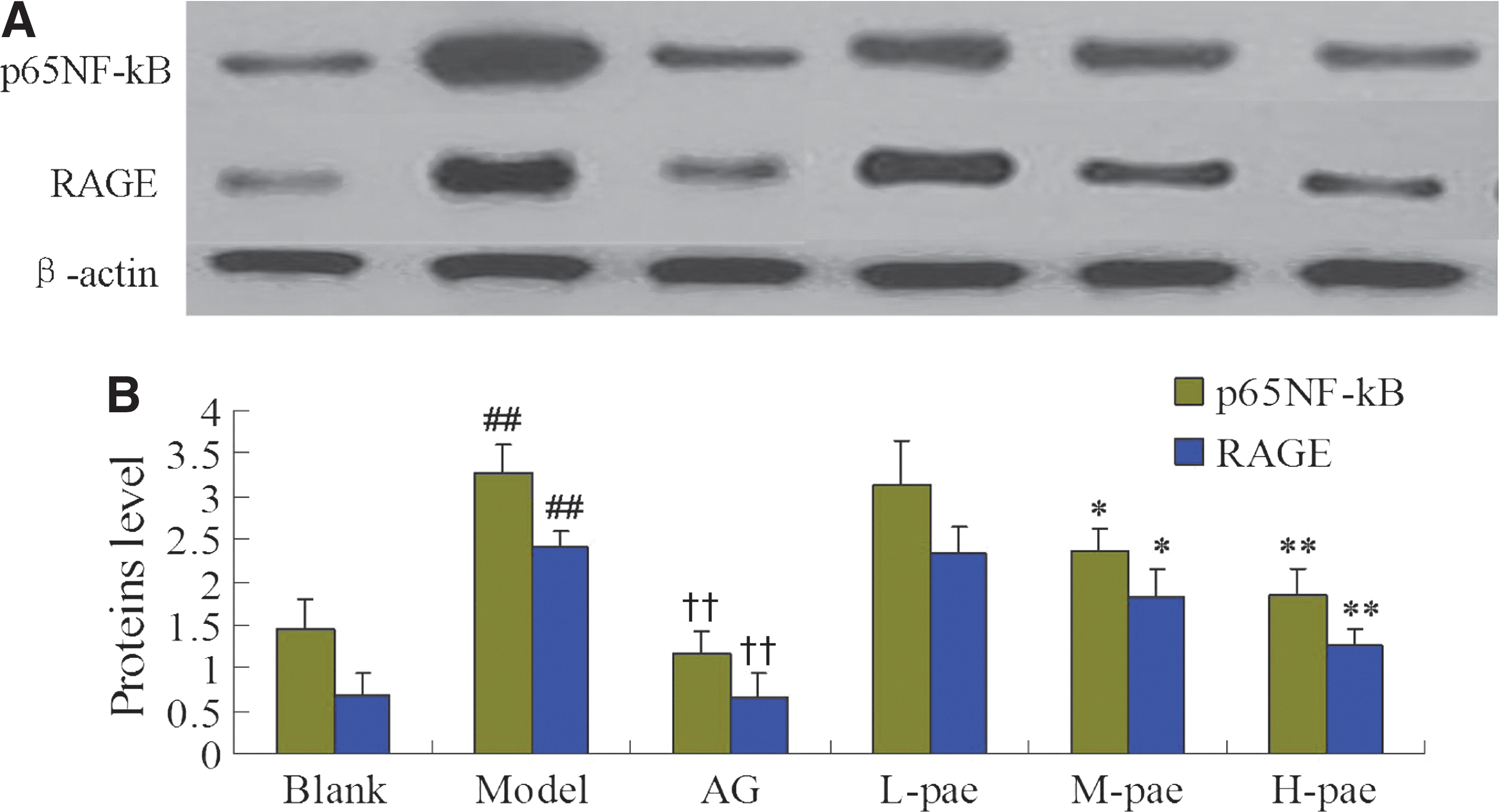
Western blotting for RAGE and NF-κB
After extraction with lysis buffer, equal amounts of supernatant protein (50 μg) was denaturated with sodium dodecyl sulfate buffer at 95°C for 10 min and electrophoresed on 12.5% Tris–HCl gels. Sequentially, the separated proteins were electrophoretically transblotted onto polyvinylidene difluoride membranes. Specific primary antibodies against NF-κB p65 (Santa Cruz; 1:200) and RAGE (Santa Cruz; 1:1000) were used for Western blot analysis of brain tissue lysates. β-Actin (1:1000) was used for the internal reference. Membranes were washed thrice with tris buffered saline and incubated with a HRP-conjugated species-specific secondary antibody (Santa Cruz). Finally, enhanced chemiluminescence solution and proteins was added for visualization.
Statistical analysis
Data from the three groups were expressed as means±standard deviation. The statistical analysis was performed by one-way analysis of variance using GraphPad Prism™ 5.0 statistical software. Differences between treatment groups were analyzed by Tukey's test. P<.05 was considered statistically significant.
Results
Paeonol significantly increased body weight in STZ-induced DE rats
STZ induced diabetic rats had significantly lower body weights (247.8±32.0 g) compared with the Blank control (396.5±34.9 g). However, the treatment with 50 mg/kg AG prevented STZ-decreased body weight in DE rats (305.2±19.3 g). Interestingly, the treatment with M-pae (50 mg/kg) and H-pae (100 mg/kg) also significantly increased body weight in DE rats (281.5±17.5 g for M-pae, 284.8±15.0 g for H-pae, vs. Model). Furthermore, paeonol increased the body weight in a dose-dependent manner (Table 1). The results indicate that paeonol could improve the body weight in DE rats.
50 mg/kg of aminoguanidine (AG), 5 mg/kg of rosiglitazone, and 3 mg/kg of donepezil were chosen as the positive control. Values are means±standard deviation. n=8.
P<.01, Model vs. Blank control.
P<.05, M-pae vs. Model; ** P<.01, H-pae vs. Model.
P<.01, AG vs. Model, rosiglitazone vs. Model.
AGEs, advanced glycation end products; GSP, glycosylated serum proteins; iNOS, induced nitric oxide synthase; GSH, glutathione.
Paeonol significantly decreased blood glucose levels
Compared with control (5.15±0.49 mmol/L), blood glucoses level in DE rats were increased significantly in this study (23.88±1.10 mmol/L). However, a significant decrease in blood glucose level was observed after treatment with AG (18.11±1.06 mmol/L, vs. Model group). More importantly, the administration of 50 or 100 mg/kg paenol markedly normalized blood glucose levels (21.10±2.87 mmol/L, M-pae vs. Model; 18.95±3.03 mmol/L, H-pae vs. Model). Although there was an insignificant decrease in the L-pae group (22.43±2.09 mmol/L), the mean blood glucose was lower than Model group (Table 1). The results show that paeonol decreases blood glucose level in STZ-induced DE rats.
Effects of paeonol on safety parameters
As shown in Table 2, no statistically significant differences in serum glutamic-oxaloacetic transaminase, serum glutamic-pyruvate transaminase, blood urea nitrogen, or creatinine in serum were found between Blank control and paeonol (100 mg/kg). These data suggest that the improvement of paeonol on DE did not induce side effects.
Values are means±standard deviation. n=8.
P<.01, Model vs. Blank control.
SGOT, serum glutamic-oxaloacetic transaminase; SGPT, serum glutamic-pyruvate transaminase; BUN, blood urea nitrogen.
Paeonol significantly decreased serum AGEs
The accumulation of AGEs has been implicated in the pathogenesis of DE. 22 In this study, the AGEs level in serum in the Model group was elevated significantly as compared with the Control (269.9±14.4 ng/L for Control, 1364.6±199.1 ng/L for Model). As shown in Table 1, AG, a well-known AGE inhibitor, significantly inhibited AGE formation in serum (55.93% inhibition). L-pae, M-pae, and H-pae significantly inhibited AGE formation up to 12.31%, 24.38%, and 32.79% at doses of 25, 50, and 100 mg/kg, respectively (M-pae vs. Model; H-pae vs. Model; Table 1). The findings demonstrate that paeonol is able to inhibit AGE formation in vivo.
Effects of paeonol on GSP
As shown in Table 1, there was a significant increase in GSP level between the Model group (2.52±0.10 mmol/L) and Blank control (1.45±0.13 mmol/L) by ∼1.74-fold. The 25, 50, and 100 mg/mL doses of paeonol decreased GSP levels up to 6.75% (2.35±0.29 mmol/L), 19.05% (2.04±0.53 mmol/L), and 26.98% (1.84±0.39 mmol/L), respectively. Compared with the Model group, AG also inhibited GSP levels up to 30.16% (1.76±0.28 mmol/L).
iNOS enzyme activity in hippocampus tissue
The NOS isoforms play an important role in hippocampus-dependent forms of learning in rats. 23 As shown in Table 1, STZ markedly enhanced the iNOS level in hippocampus of DE rats as compared with the Blank control (2.140±0.215 U/mg protein for Model and 0.406±0.033 U/mg protein for Blank control). Interestingly, the STZ-increased iNOS level in the hippocampus was decreased significantly by the treatment with paeonol of 50 and 100 mg/kg (1.791±0.336 U/mg protein for 50 mg/kg paeonol; 1.764±0.219 U/mg protein for 100 mg/kg paeonol). However, the AchE inhibitor donepezil did not significantly affect the STZ-increased iNOS levels, whereas the hypoglycemic drug rosiglitazone significantly prevented increases in iNOS levels. The results show that hippocampal iNOS levels of DE rats were correlated negatively with performance. These results suggest that the effect of paeonol on the STZ-enhanced hippocampal iNOS levels might contribute to the amelioration of memory deficits in DE rats.
GSH content in hippocampus tissue
Oxidative stress plays a crucial role in the development and progression of DE. GSH, a small molecule peptide consisting of three amino acids, has been regarded as an important factor in the antioxidant and scavenging of free radicals in vivo. After being induced by 40 mg/kg STZ, the hippocampal GSH content was lower compared with Blank control (33.63±4.09 μmol/L vs. 51.98±5.37 μmol/L). More importantly, the GSH content in the hippocampus increased significantly to 39.06±3.51 and 46.05±4.49 μmol/L after the administration of 50 and 100 mg/kg paeonol (M-pae vs. Model; H-pae vs. Model). For the positive drugs, there was no significant difference between donepezil and Model, whereas a significant increase between rosiglitazone and Model was observed under our experimental conditions (Table 1). Our data support the premise that paeonol has antioxidant activity on the attenuation of memory deficits in DE rats.
Amelioration by paeonol of RAGE and NF-κB expression in hippocampus and cerebral cortical neuron
Studies suggest that RAGE was markedly expressed in neurons and mediated a neuroinflammatory response associated with the pathogenesis of primary DE. 24 The overexpression of RAGE resulted in some significant structural changes in the hippocampus and adversely affected cognitive development in diabetes. Compared to the Blank control, a significant upregulation on the brown positive expression of RAGE protein was observed in Model rats. A comparison between Model and paeonol depicted that paeonol down-regulated the over expression of RAGE in a dose-dependent manner (Figs. 2A–C and 3A, B).
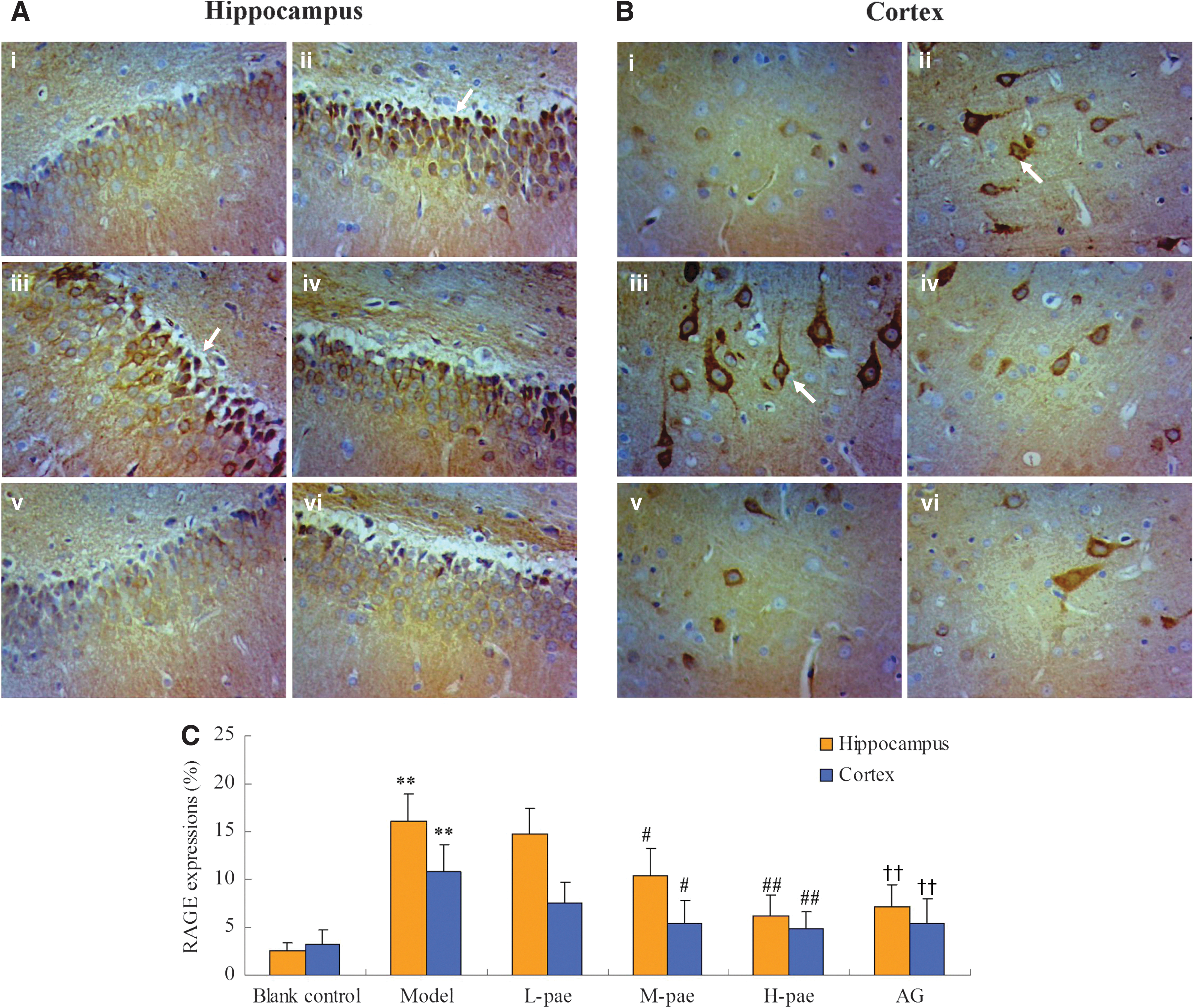
RAGE protein expression in the hippocampus
It has been confirmed that long term hyperglycemia can elicit and accelerate the degeneration of neurons and the up-regulation of RAGE and NF-κB p65 in the brain of diabetic rats. 19 Herein, an obvious brown positive expression of NF-κB was observed in hippocampus and cerebral cortical neuron, compared with the blank control. After 30 days of paeonol treatment, the upregulation of NF-κB was ameliorated significantly in a dose-dependent manner (Figs. 3A, B and 4A–C).
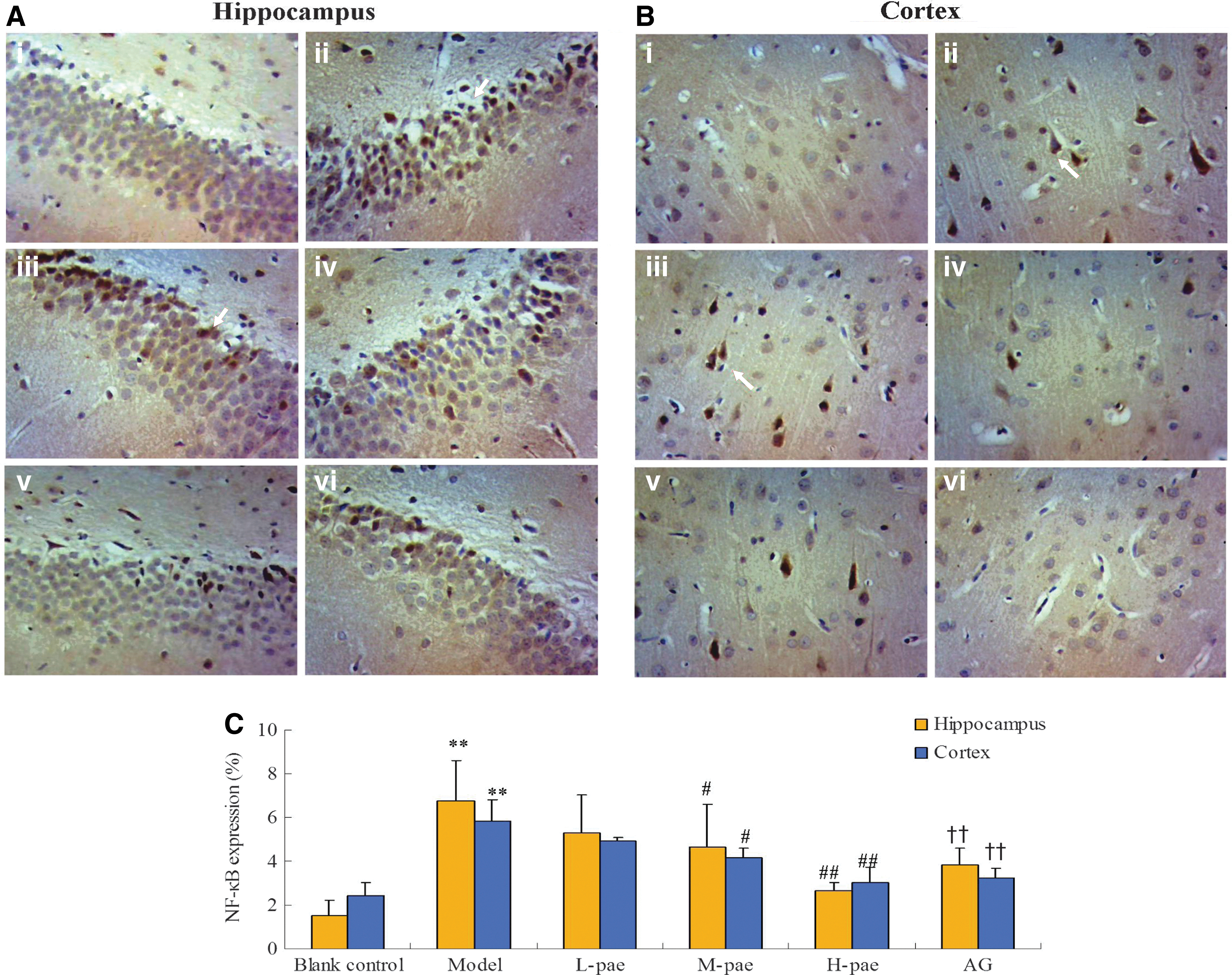
NF-κB expression in the hippocampus
Discussion
DE, one of the common diabetic complications, causes chronic cognitive disturbance in long-standing diabetes. 25 Accumulating evidence showed that it might be related to regional tissue pathological changes in the brain. 26 Aβ deposit are an established index of cognitive deficits. Therefore, in this study, Aβ deposit expression increased significantly following treatment with high fat and sucrose diet and STZ (Fig. 5A, B). A poor performance in Morris water maze (data not shown), confirmed that the rats with DE had cognitive deficits. Furthermore, it is known that consistent hyperglycemia contributes to the pathogenesis and the development of DE. The accumulation of GSP levels (GSP) and AGEs, derived from the nonenzymatic glycation of reducing sugars and proteins, contribute to the development of DE. 19 The daily intake of 50 and 100 mg/kg paeonol significantly increased body weight and decreased blood glucose, GSP levels, and as serum AGEs levels in STZ-induced DE rats. The findings indicate that paeonol has a beneficial effect on DE.
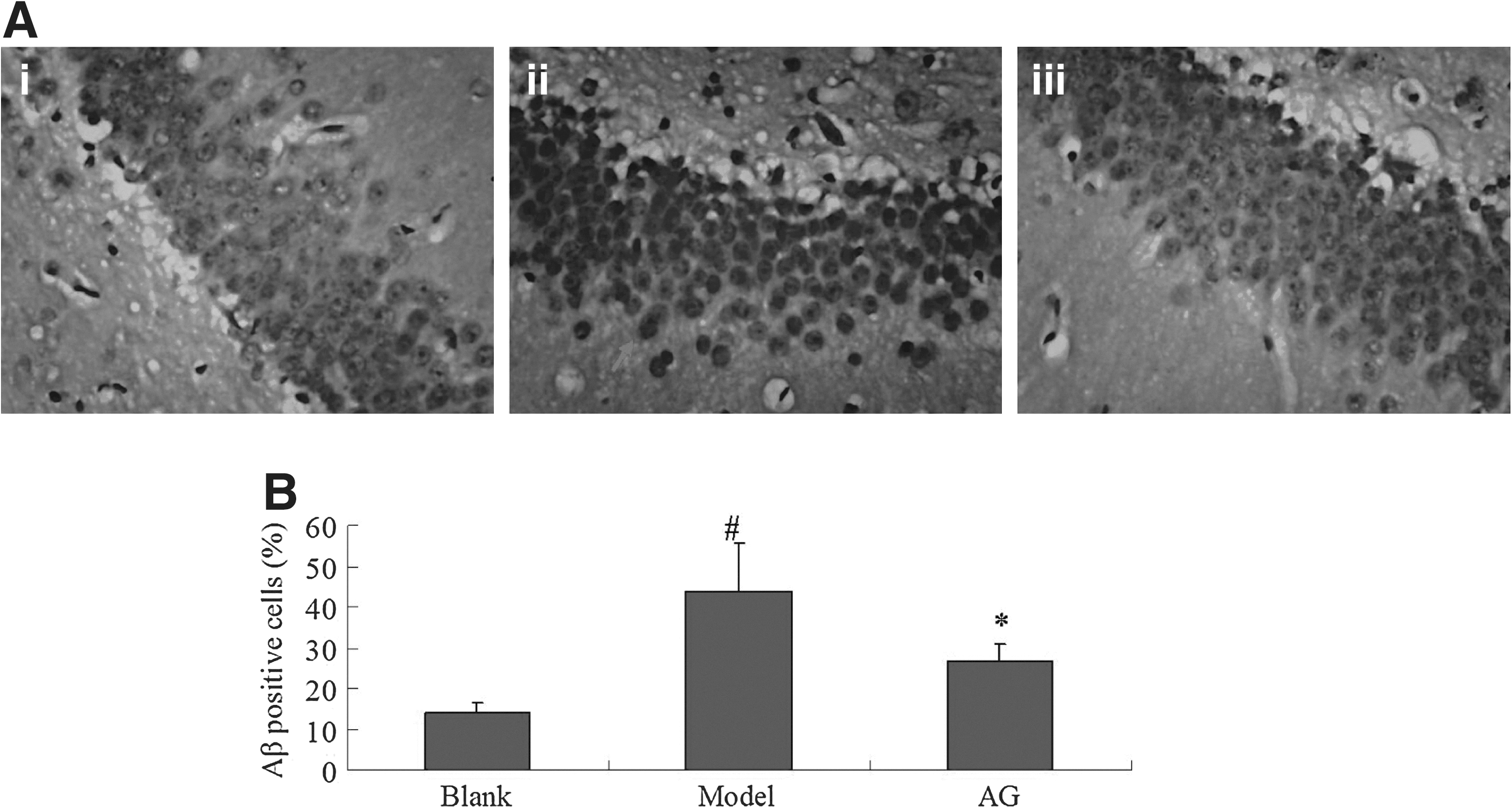
The effect of high fat/high sucrose diet+STZ on the hippocampus of rats. Hippocampus was stained by Congo Red.
AGEs are the final products of nonenzymatic glycation and oxidation of proteins and lipids, which have been found to deposit in human tissues with aging and brains of diabetic patients. 27 AGEs are believed to play a crucial role in the development and the progression of DE by the damaging the structure and functional properties of the hippocampus and cerebral cortical neurons via binding to cell surface receptors RAGE. 17 The receptor for AGEs (RAGE) is a multiligand receptor protein thought to play an important role in neuronal damage. 28 The interaction between AGEs and RAGE activates a variety of signalling pathways that lead to the pathogenesis of DE, such as neurodegeneration. 29 Our study also provided further confirmation that the accumulation of AGEs activated RAGE and upregulated RAGE protein expression in hippocampus and cerebral cortical neurons. However, paeonol treatment ameliorated the pathological changes associated with DE. These results suggest that the decreased hyperglycemia with paenol consumption might contribute to the amelioration of hyperglycemia-induced encephalopathy damage.
Oxidative stress has been regarded as an important in the pathogenesis of DE. 30 Hyperglycemia-induced oxidative stress accelerated oxidative damage of diabetic brain. Neuronal damage was associated with both nitric oxide (NO) levels and ROS levels, which were higher than the normal control. It has been shown that the imbalance of mitochondrial NOS expression and GSH levels contributes to the impaired cognitive functions and neurochemical and structural abnormalities in diabetes. 31 In the present study, hippocampal iNOS activity and GSH content was determined to explore the regulation effect of paeonol on the biochemical changes in brain of DE rats.
NO has been demonstrated to play an important role in the pathophysiology of learning and memory ability. 32 Several lines of research have shown that NOS activity is associated with cognitive impairment in patients with dementia or DE. 33,34 The clear activity changes of iNOS in the hippocampus and cerebral cortex was shown after treatment with 40 mg/kg STZ. Remarkably, the hippocampal iNOS activity was negatively associated with performance in DE rats. 32 The improved iNOS activity suggested that by regulating iNOS activity, paeonol had potential antioxidant activity for attenuating the cognitive impairment in DE rats.
The overgeneration of ROS or NO upsets the original balance of oxidant/antioxidant system and modulates the endogenous thiol antioxidant GSH with high oxygen consumption. 35 The antioxidant effects of GSH were decreased by an i.p. injection of STZ. 36 A significant elevation in GSH activity was also observed due to the treatment with paeonol. The combined results of hippocampal iNOS activity and GSH content revealed that paeonol attenuated STZ-induced oxidative stress and prevented cognitive impairment by modulating iNOS activity and antioxidant status of the neuronal tissue.
Nuclear translocation of transcription factors, such as NF-κB, a key regulator of immune and inflammatory response, is believed to play a key role in the pathogenesis of DE. 37 Immunostaining for NF-κB showed that NF-κB was mainly expressed in the cytoplasm. In the present study, a significant overexpression of NF-κB in the hippocampus and cerebral cortical neurons of diabetic rats was observed. More importantly, paeonol significantly inhibited hyperglycemia-induced in NF-κB p65 nuclear translocation in the hippocampus and prefrontal cortex of STZ-treated rats. 38,39 These findings suggest that paeonol could ameliorate DE associated with NF-κB activation.
A number of studies have shown that the AGE/RAGE/NF-κB pathway are involved in the pathogenesis of chronic complications of diabetes through activating inflammatory systemic responses. 17 Moreover, the AGE/RAGE/NF-κB pathway plays an important role in the progression of various neurodegenerative disorders, including DE. 40 The above protective effects of paeonol on attenuating STZ-induced brain damage might be associated with the inhibition of the AGE/RAGE/NF-κB pathway. It might also be associated with the blockade of paeonol on AGE and RAGE interaction due to the carbonyl groups of paeonol. Previous reports showed that the ligand of RAGE contained –COOH tails and mediated the binding to RAGE. 41 However, this speculation should be confirmed in further experiments.
In conclusion, paeonol has significant hypoglycemic and antioxidative activites for improving pathological changes in the hippocampus and cortex of STZ-induced DE rats. Therefore, peaonol might be beneficial for the prevention and treatment of DE.
Footnotes
Acknowledgments
The authors are grateful for the financial support of the Education Department of Shaanxi Province (No. 12JK1019), Creative Scientific Research Projects for postgraduate students in Jiangsu General Colleges (CXLX12_0328), National Natural Science Foundation of China (No. 81202906), Natural Science Foundation of Jiangsu Province (BK2012491), and the Priority Academic Development Program of Jiangsu Higher Education Institutions.
Author Disclosure Statement
No competing financial interests exist.