
Abstract
The present report examined the effects of essential oils (EOs) from Satureja hortensis L. and Salvia fruticosa M. on the viability and outer membrane permeability of the periodontopathogen Fusobacterium nucleatum, a key bacteria in oral biofilms, as well as the inhibition of matrix metalloproteinase (MMP-2 and MMP-9) activities in epithelial cells exposed to such bacteria. Membrane permeability was tested by measuring the N-phenyl-1-naphthylamine uptake and bacterial viability by using the commercially available Live/Dead BacLight kit. In addition, gelatin zymography was performed to analyze the inhibition of F. nucleatum-induced MMP-2 and MMP-9 activities in HaCaT cells. We showed that 5, 10, and 25 μL/mL of Sat. hortensis L. EO decreased the ratio of live/dead bacteria and increased the outer membrane permeability in a range of time from 0 to 5 min. Treatments with 10 and 25 μL/mL of Sal. fruticosa M. also increased the membrane permeability and 5, 10, and 25 μL/mL of both EOs inhibited MMP-2 and MMP-9 activities in keratinocytes induced after exposure of 24 h to F. nucleatum. We conclude that antibacterial and antigelatinolytic activities of Sat. hortensis L. EO have potential for the treatment of periodontal inflammation.
P
Aerial parts of Satureja hortensis L. are frequently used as a food additive and also as a traditional remedy. 8 We have recently reported strong growth inhibitory characteristics of its essential oil (EO) against common periodontitis-associated bacteria. 9 In our study, in addition to F. nucleatum, other common periodontopathogens were used, including strains of Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis, Parvimonas micra, Tannerella forsythia, Prevotella intermedia, and Prevotella nigrescens. Sat. hortensis L. EO inhibited the growth of periodontal bacteria in a concentration that had no obvious deleterious effects on the growth or viability of human skin keratinocytes. Further, our group has also described antigelatinolytic activities of a number of EOs. 10 This short communication reports the effects of two EOs from the Lamiaceae (Labiatae) family, Sat. hortensis L. and Salvia fruticosa M., on the outer membrane permeability and viability of F. nucleatum. Second, the inhibition of F. nucleatum-induced MMP-2 and MMP-9 activities in epithelial cells by Sat. hortensis L. and Sal. fruticosa M. EOs is demonstrated.
Leaves from Sat. hortensis L. and Sal. fruticosa M. were collected in the West-Mediterranean region of Turkey from May to July 2006 and the major constituents of the EOs utilized in this report were previously described by our group: carvacrol (86.77%), cymene (4.39%), α-pinene (1.50%), terpeniol (1.02%), thymol (0.74%), β-pinene (0.55%), linalool (0.49%), borneol (0.47%), and myrcene (0.01%) for Sat. hortensis L.; and 1,8-cineole/eucalyptol (49.5%), camphor (13.3%), β-pinene (7.2%), α-pinene (5.8%), and camphene (5.1%) for Sal. fruticosa M. 9,10
The F. nucleatum American Type Culture Collection (ATCC) 25586 strain was used for the experiments. The strain was revived from a frozen (−70°C) stock and subcultured for purity on brucella blood agar, enriched with hemin, and incubated at 37°C in an anaerobic chamber. Afterward, cultures were grown in a Brain Heart Infusion broth supplemented with 1% sucrose, at 37°C in an anaerobic chamber. After 48 h of incubation, bacteria were washed twice by centrifugation and adjusted to an optical density (OD) at 660 nm of 1.0 with phosphate-buffered saline (PBS).
To determine the bacterial outer membrane permeability, an adjusted bacterial solution was resuspended in 10 μM of N-phenyl-1-naphthylamine (NPN; Sigma-Aldrich, 104043, Saint Louis, MO, USA) together with 5, 10, and 25 μL/mL of EOs. The NPN uptake was fluorometrically measured (excitation: 360 nm; emission: 460 nm) in a range of time from 0 to 5 min (T=0–5 min). For elucidating the bacterial viability, a commercial kit was used (Live/Dead BacLight bacterial viability kit; Molecular Probes Inc., Eugene, OR, USA). Briefly, an adjusted bacterial solution was resuspended and stained with SYTO9/propidium iodide (PI) together in the absence and presence of 5, 10, and 25 μL/mL of EOs for 15 min. After incubation, fluorometric readings were performed for SYTO9 (excitation: 485 nm, emission: 528 nm) and PI (excitation: 485 nm, emission: 645 nm) in a range of time from 0 to 5 min (T=0–5 min).
For the analysis of F. nucleatum-induced MMP-2 and MMP-9 activities, HaCaT human skin keratinocytes (generously provided by Hubert Fusenig, German Cancer Center, Heidelberg, Germany) were used. Cells were grown in 96-well plates to 80% of confluence. For these experiments, F. nucleatum stocks were prepared by adjusting to an OD at 490 nm of 0.5 with PBS. Before adding 30 μL of F. nucleatum suspension in PBS into the test wells, an aliquot of 270 μL of fresh medium was placed to each well. Each EO was added into the culture medium (to final volumes of 5, 10, and 25 μL/mL). PBS was used as a negative control. After 24 h of incubation, 20 μL of medium from the wells were collected and used for gelatin zymography analyses by subjecting the samples to sodium dodecyl sulfate–polyacrylamide gel electrophoresis and using 8% gels containing 1 mg of gelatin/mL, as previously described. 10 Gelatin zymography experiments were repeated twice in triplicates.
Statistical analyses were performed by using the Student's t-test. Results were considered significant with P values<.05 and are representative of three independent experiments.
According to the results, the outer membrane permeability of F. nucleatum was increased (P value<.01) after 5 min of incubation with 5, 10, and 25 μL/mL of Sat. hortensis L. EO, while Sal. fruticosa M. EO was able to increase the permeability (P value<.05) only after treatments with 10 and 25 μL/mL (Fig. 1A). The ratio of live/dead bacteria was decreased (P value<.01) by all the treatments with Sat. hortensis L. EO (5, 10, and 25 μL/mL) in a range of time from 0 to 5 min (Fig. 1B). In a similar interval of exposure, treatments with Sal. fruticosa M. EO failed to decrease the bacterial viability (Fig. 1B). In addition, both Sat. hortensis L. and Sal. fruticosa M. EOs, at concentrations of 5, 10, and 25 μL/mL, inhibited F. nucleatum-induced MMP-9 (92 kDa band) and MMP-2 (72 kDa band) activities (Fig. 2).

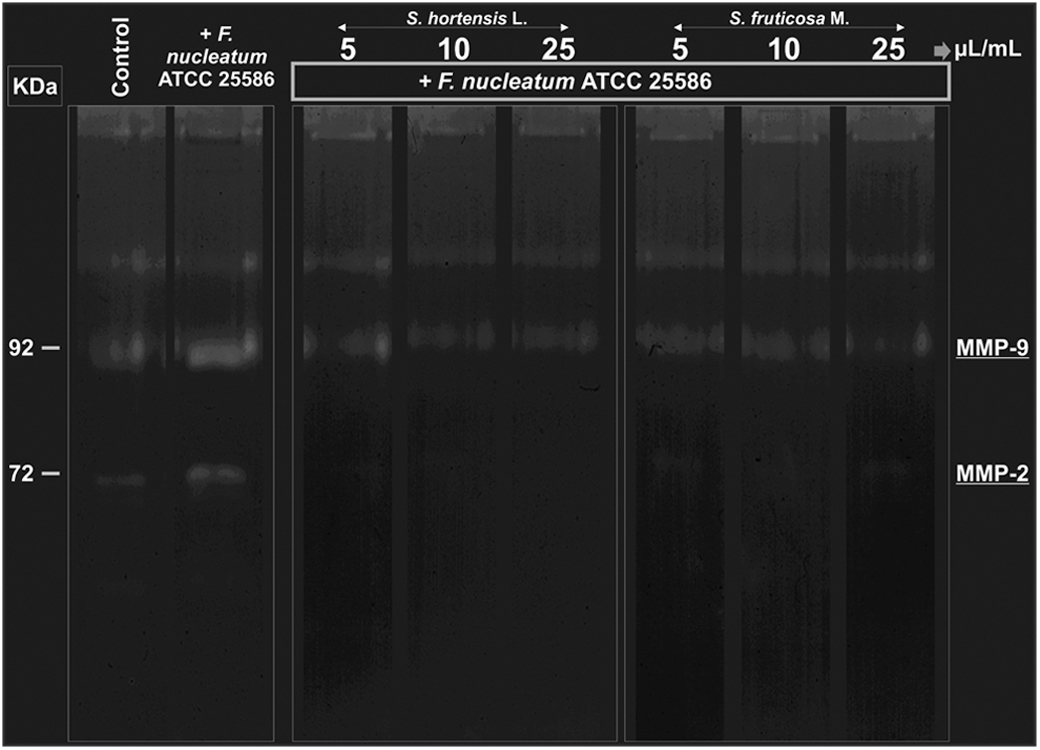
Antigelatinolytic effects of Sat. hortensis L. and Sal. fruticosa M. EOs (5, 10, and 25 μL/mL) on human keratinocytes (HaCaT cells) after 24 h of bacterial exposure (F. nucleatum).
Species from the Lamiaceae (Labiatae) family are commonly used plants for their antimicrobial properties. 11,12 In this work, all major constituents of Sat. hortensis L. and Sal. fruticosa M. EOs are terpenes/terpenoids. 9,10 The antimicrobial effect of three monoterpenes like menthol, thymol, and linalyl acetate has been proposed to be due to the perturbation in the lipid fraction of the bacterial plasma membrane resulting in alterations of its permeability and subsequently, the penetration of the EO into the microorganism where it may interact with intracellular sites critical for the antibacterial activity. 13 Terpenes, as lipophilic compounds, interact with the hydrophobic part of the membrane and affect the membrane anisotropy and dipolar organization. 14 In this report, both Sat. hortensis L. and Sal. fruticosa M. EOs increased the permeability of the F. nucleatum outer membrane, which is consistent with the elevated percentage of terpenes/terpenoids present in their composition. The effect was faster and required smaller quantities of EO (μL/mL) for Sat. hortensis L. than those needed and exerted by Sal. fruticosa M. This could be related to the higher number and percentage of terpenes in the composition of Sat. hortensis L. and explain why the treatments with Sal. fruticosa M. EO did not show a decreased ratio of live/dead bacteria, unlike that with Sat. hortensis L. EO. It cannot be excluded that longer incubation periods with Sal. fruticosa M. EO could trigger significant effects on the bacterial viability. In fact, our group has previously demonstrated antibacterial activities of Sat. hortensis L. and Sal. fruticosa M. EOs against F. nucleatum strains and other common periodontopathogens by using the disc diffusion assay. 10,15 Although F. nucleatum is not regarded as a major pathogen in periodontal diseases, it is the main organism that connects early and late colonizers. 3,16,17 This coaggregation ability regulates the maturation of periodontal biofilms. Therefore, F. nucleatum represents an excellent bacterial target for developing therapeutic agents against periodontitis when the aim is to eradicate biofilms. In the present study, we utilized F. nucleatum-induced MMP stimulation in epithelial cells as an in vitro model (bacteria/epithelial cell coculture). The selected ATCC 25586 strain of F. nucleatum was shown to be the best MMP inducer (MMP-2, MMP-9, and MMP-13) according to our previous study. 5 The strain possesses adhering, invading, and β-defensin secretion inducing properties, and it has also been characterized at the genomic level. 4,18,19 In other words, ATCC 25586 is a representative strain to evaluate the virulence characteristics of F. nucleatum.
The regulation of the activation of MMPs by redox status is already known in the literature. 20 Reactive oxygen species (ROS), especially hydrogen peroxide (H2O2), are considered critical interplayers in the regulation of MMPs through the oxidation of the zinc-cysteine switch. ROS also modulate the phosphatase activity, a number of kinase cascades, transcription factor binding, and MMP latency. 20
In a recent study by Anthony and colleagues, 21 423 EOs belonging to 48 different botanical families were evaluated for their antioxidant activities as free radical scavenging agents. They showed that EOs of the botanical families Lamiaceae (Labiatae) and Myrtaceae were the most effective antioxidants. Thus, it would be expected that EOs from plants belonging to the Lamiaceae (Labiatae) family, like Sat. hortensis L. and Sal. fruticosa M. EOs, with a well-known antioxidant activity, could inhibit the bacteria-induced MMP activity in human epithelial cells by neutralizing the pathogenic generation of ROS. 22,23 In fact, it has been shown in silico that MMPs and a number of major components of EOs interconnect through the interaction with hydroxyl radicals, molecular oxygen, and H2O2. 10
In summary, Sat. hortensis L. EO increases the outer membrane permeability and decreases the viability of F. nucleatum as well as inhibits F. nucleatum-induced MMP-2 and MMP-9 activities in human epithelial cells. Therefore, our results require attention and further evaluation since Sat. hortensis L. EO may have the potential, first, as an adjunctive anti-infectious agent able to increase the sensitivity of gram-negative bacteria to hydrophobic antibiotics that alone inefficiently traverse the outer membrane and, second, as a host-modulating agent able to inhibit the bacteria-induced MMP activity in human epithelial cells and, consequently, periodontal tissue destruction.
Footnotes
Acknowledgments
The authors thank the National Council for Scientific and Technological Development (CNPq) of the Federal Republic of Brazil, CAPES (PROCAD 066/2007), FAPERGS (PqG 1008860, PqG 1008857, ARD11/1893-7, and PRONEX 1000274), PROPESQ-UFRGS, IBN-Net No. 01.06.0842-00, and the University of Helsinki (HUS-EVO grant) for supporting this work. Fares Zeidán-Chuliá acknowledges the Marie Curie Early Stage Research Training (EST) program for his previous funding at the University of Helsinki, Finland.
Author Disclosure Statement
No competing financial interests exist.