
Abstract
Antimetabolites have proven successful as therapeutics for advanced-stage breast cancers, but are often accompanied by severe side effects that can limit treatment regimens. 5-Fluorouracil (5-FU), an antimetabolite that inhibits cell proliferation, has served an important role in standard chemotherapy protocols for a variety of solid tumors. Although reasonable response rates have been reported for 5-FU, continued exploration is necessary to improve clinical outcomes and reduce cytotoxic side effects that are an inherent problem for chemotherapeutic interventions. Because of its diverse anticancer properties, we explored whether by combining the natural product curcumin with 5-FU, synergistic improvements in preventing breast cancer cell proliferation and/or provide protection against 5-FU-induced cytotoxicity could be achieved. Indeed both curcumin and 5-FU inhibit DNA synthesis in MDA-MB-231 cells using BrdU incorporation assays; however, combined treatment showed no synergistic improvement. We next established the cytotoxicity profile for 5-FU in MDA-MB-231 cells using a tetrazolium-based cell viability assay and obtained an LD50 value of 28 μM. When 5-FU incubations were repeated with the addition of curcumin, the LD50 value increased to 200–300 μM, representing a 7–10-fold protection by curcumin against 5-FU cytotoxicity. These findings suggest that the addition of curcumin as an adjuvant therapy during 5-FU treatment might enhance the chemotherapeutic effectiveness of 5-FU by protecting normal cells from reduced viability and thus permitting higher dosing or longer treatment times. This would be especially important to those individuals who are plagued with severe cytotoxicities and require frequent interruptions, or even early termination of their treatment regimens.
Introduction
I
Current treatment options are largely dependent on estrogen receptor (ER) status and pathological staging of the tumor. 2 Approximately 50% of patients with ER-positive tumors are responsive to antiestrogen therapy, 3 while more advanced, ER-negative breast tumors require a more aggressive approach, including radiation treatment and chemotherapy. 4 The most commonly used chemotherapeutic agents include anthracyclines, taxanes, alkylating agents, alkaloids, nucleotide analogs, and topoisomerase inhibitors. 4 These compounds impair cancer growth and progression by a variety of mechanisms, including intercalation between base pairs of DNA/RNA strands, disruption of microtubule polymerization/depolymerization, alkylation of DNA, and suppression of proper coiling and uncoiling of DNA. 4 Antimetabolites have also been used extensively to inhibit cell proliferation by inhibiting DNA replication. One such compound, 5-fluorouracil (5-FU), has provided significant clinical benefit as an anticancer therapeutic for over 30 years, 5,6 including as an adjuvant therapy 7 and in the treatment of metastatic disease. 8 5-FU is a nucleotide analog that prevents chromosomal replication by inhibiting thymidylate synthase activity, which is required for conversion of 2′-deoxyuridine-5′-monophosphate (dUMP) to 2′-deoxythymidine-5′-monophosphate (dTMP). This inhibition reduces the amount of available dTTP for DNA synthesis thereby preventing cell growth. 5-FU has served an important role in standard chemotherapy protocols for a range of solid tumors, including breast and colorectal cancers. 9,10 Response rates of 25–35% have been reported when 5-FU was administered as a single-agent bolus injection. 5,8,11 To increase its therapeutic efficacy, administration of 5-FU by repetitive intravenous injections or continuous infusion for extended times has been investigated. 12,13 These approaches have shown some success until toxicity supervenes, which necessitates a planned break between treatment cycles. 5 Cytotoxicity resulting from 5-FU treatment can be substantial; for example, 31% to 34% of patients treated with 5-FU experience dose-limiting toxicities. 14 The primary mechanism of cytotoxicity of 5-FU has been attributed to incorporation of fluoronucleotides into RNA and DNA and to the direct inhibition of thymidylate synthase. 15 Clinical side effects resulting from 5-FU cytotoxicity include hematologic (anemia and thrombocytopenia), as well as gastrointestinal (oral and intestinal mucositis, stomatitis, diarrhea, nausea, and vomiting) complications. 16
To improve upon the efficacy of 5-FU as an anticancer therapeutic, it has been administered in combination with other compounds with the anticipation that such combinations might enhance the antiproliferative activity of 5-FU. One compound tested in this manner is the natural product curcumin, which by itself has been shown to inhibit prosurvival signals in certain cancer cell lines, including colorectal, gastric, and prostate cancers, 17,18 as well as significantly enhance apoptosis. 19 When the human colon cancer cell line HT-29 was treated with both curcumin and 5-FU, a synergistic inhibition of growth was observed. 18 Similar results were found when dual treatment was applied to a human gastric carcinoma cell line. 20 In a human esophageal squamous cell carcinoma (ESCC) xenograft model, curcumin or 5-FU individually reduced tumor volume; however, their combination had the strongest anticancer effects with reduced cell proliferation and higher levels of apoptosis than measured for 5-FU treatment alone. 21
Based on these findings, we examined whether curcumin provided similar synergistic antiproliferative effects with 5-FU in breast cancer cells. Combined therapy with these two compounds as a potential therapeutic intervention for breast cancer has not been explored, and if found effective, then it may offer a novel treatment option for inhibiting late-stage, ER-negative tumor growth.
Materials and Methods
Cell culture
MDA-MB-231 human breast cancer cells (American Type Culture Collection, Manassas, VA, USA) were cultured in Dulbecco's modified Eagle's medium (DMEM; Life Technologies/Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal calf serum (Irvine Scientific, Santa Ana, CA, USA), 0.1 mM non-essential amino acids, 1 mM sodium pyruvate, 10 μg/mL bovine insulin, and 100 units/mL penicillin G. Cells were grown in a humidified incubator at 37°C with 5% CO2/95% air and passaged twice a week.
Wst-1 and BrdU assays
For Wst-1 and BrdU assays, cells (100 μL) were added to wells of a 96-well culture plate diluted to a final concentration of 50,000 cells/well and 2000 cells/well, respectively, and incubated for 24 h at 37°C with 5% CO2/95% air. Complete DMEM phenol red-free media containing the indicated concentrations of curcumin or 5-FU were added to cells and cells were incubated for the indicated times. For cell viability assays, Wst-1 reagent (Roche Diagnostics, Indianapolis, IN, USA) was added to media in each well at a final dilution of 1:10 from manufacturer stock solution and the cultures were further incubated for 1 h at 37°C. Absorbance was then measured at 460 nm with a reference wavelength of 650 nm. For cell proliferation assays, BrdU (Roche Diagnostics) was added to each well at final concentration of 10 μM and cells were incubated for 16 h at 37°C. Unincorporated BrdU was then removed by rinsing cells three times with phosphate-buffered saline (PBS) and cells were fixed for 30 min at 25°C with 100 μL FixDenat. Anti-BrdU horseradish peroxidase conjugated antibody (100 μL of 1:100 dilution of manufacturer stock solution) was added to each well followed by a 90-min incubation at 25°C. Cells were rinsed three times with PBS using a BioTek plate washer followed by an addition of 100 μL/well 3,3′,5,5′-tetramethylbenzidine (0.4 g/L, in 26% N,N-dimethylformamide). After 30 min at 25°C, absorbance was measured at 370 nm.
Statistical analysis
P-values were determined by performing t-tests on replicate samples using Sigma Plot (Version 7.0). Statistical significance was confirmed when P<.05.
Results
Curcumin inhibits DNA synthesis at concentrations below its cytotoxic threshold
Before determining whether curcumin is able to synergize with 5-FU in preventing breast cancer cell proliferation, we performed a titration study with curcumin alone to identify its LD50 for inhibiting DNA synthesis. First, it was necessary to distinguish direct inhibition of DNA synthesis from potential cytotoxicity to avoid using concentrations of curcumin that would present false positive data in our antiproliferative assay. For this, MDA-MB-231 breast cancer cells were incubated with various concentrations of curcumin for 24 h, after which cell viability was assessed by measuring functional dehydrogenase activity using a Wst-1 assay. A small, but measurable amount of cytotoxicity was seen beginning with incubations of curcumin at 10 μM (Fig. 1). The LD50 for curcumin-mediated cytotoxicity in MDA-MB-231 cells was calculated as 70 μM.
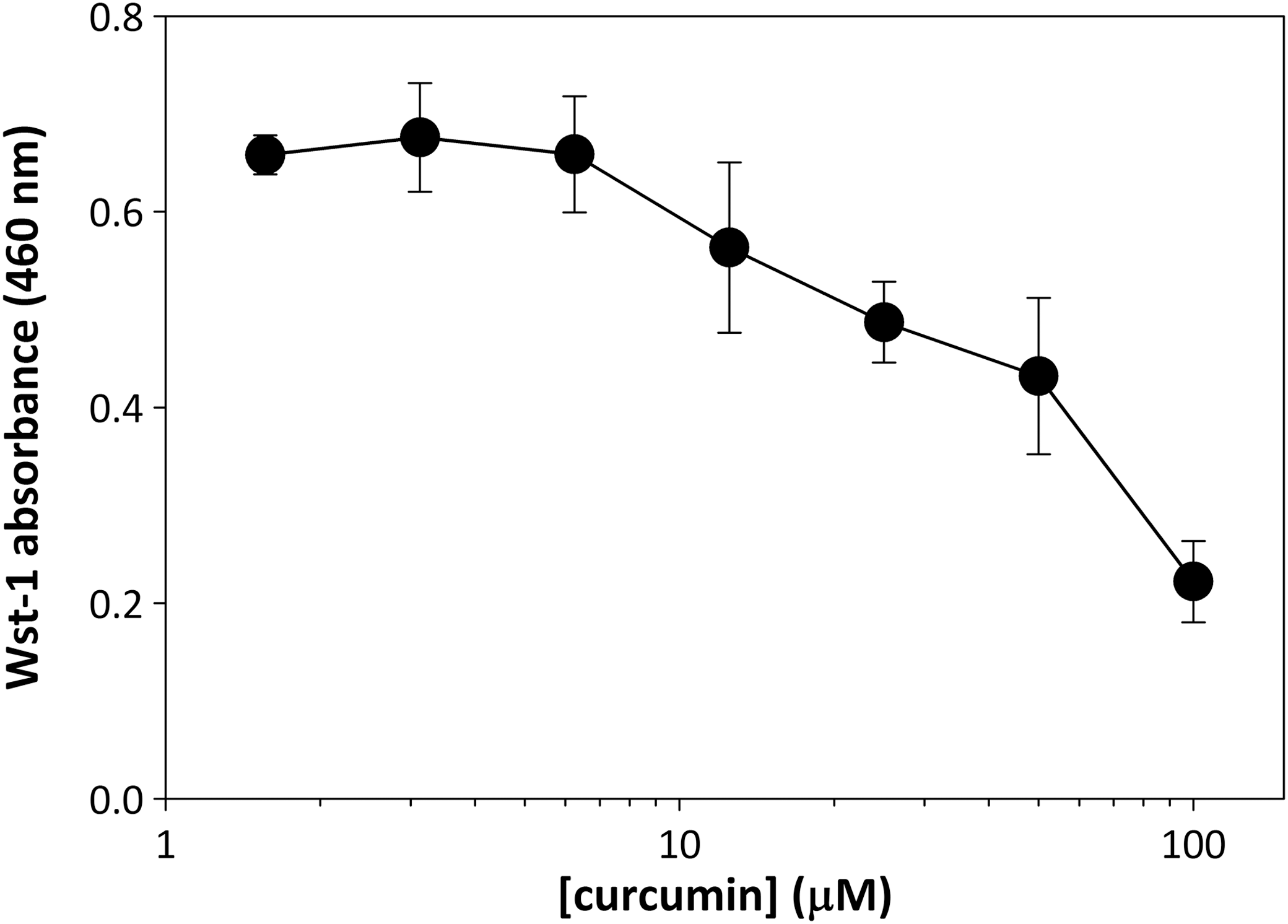
Cytotoxicity of curcumin in MDA-MB-231 cells. MDA-MB-231 cells were incubated with curcumin at concentrations of 0, 1.56, 3.125, 6.25, 12.5, 25, 50, or 100 μM. Following a 24-h incubation, Wst-1 reagent was added to media and cells were incubated for 1 h at 37°C. Absorbance was measured at 460 nm. Error bars represent standard deviation of triplicate points. LD50 calculated as 70 μM.
Next, to measure the effectiveness of curcumin as an antiproliferative agent for breast cancer cells, we performed a titration study using a BrdU incorporation assay to examine whether curcumin is capable of directly inhibiting DNA synthesis. As before, MDA-MB-231 breast cancer cells were incubated with various concentrations of curcumin for 24 h, followed by a measurement of BrdU incorporation to directly quantify DNA synthesis. At concentrations of curcumin > 12.5 μM, DNA synthesis was impaired and fully inhibited with ≥25 μM (Fig. 2). The IC50 value for inhibition of DNA replication by curcumin was calculated as 18 μM, which is well below the LD50 value for cytotoxicity, indicating that curcumin provides antiproliferative activity that is distinct from cytotoxicity in human breast cancer cells.

Effect of curcumin on DNA synthesis in MDA-MB-231 cells. Cells were incubated with curcumin at concentrations listed in Figure 1 for 24 h. Cells were then incubated with BrdU for an additional 16 h in the continued presence of curcumin. BrdU incorporated into DNA was quantified by ELISA. Error bars represent standard deviation of triplicate points. IC50 calculated as 18 μM.
Curcumin does not synergistically inhibit DNA synthesis with 5-FU
A similar titration study was performed for 5-FU as done for curcumin to quantify the antiproliferative activity of 5-FU in human breast cancer cells. As anticipated, incubation of MDA-MB-231 cells with various concentrations of 5-FU was found to inhibit DNA synthesis in a dose-dependent manner (Fig. 3). The IC50 value of 5-FU as measured by the BrdU assay is 16 μM (Fig. 3), indicating that 5-FU and curcumin (IC50, 18 μM) share similar potencies as antiproliferative compounds in MDA-MB-231 cells. To determine whether curcumin and 5-FU function synergistically in preventing DNA synthesis, cells were once again incubated with various concentrations of 5-FU but with 6 μM curcumin included at each point measured. A concentration of 6 μM curcumin was chosen for this comparison to minimize cytotoxic effects seen at concentrations ≥10 μM. In addition, a previous study showed a significant synergistic inhibitor effect in AGS gastric carcinoma cells when 5 μM curcumin was selected for these conditions. 20 However, in contrast to results seen for esophageal, 21 gastric, 20 or colon cancer, 18 the IC50 value for 5-FU remained the same at 16 μM, demonstrating that no synergistic enhancement of antiproliferative activity by curcumin was seen in breast cancer cells (Fig. 3).
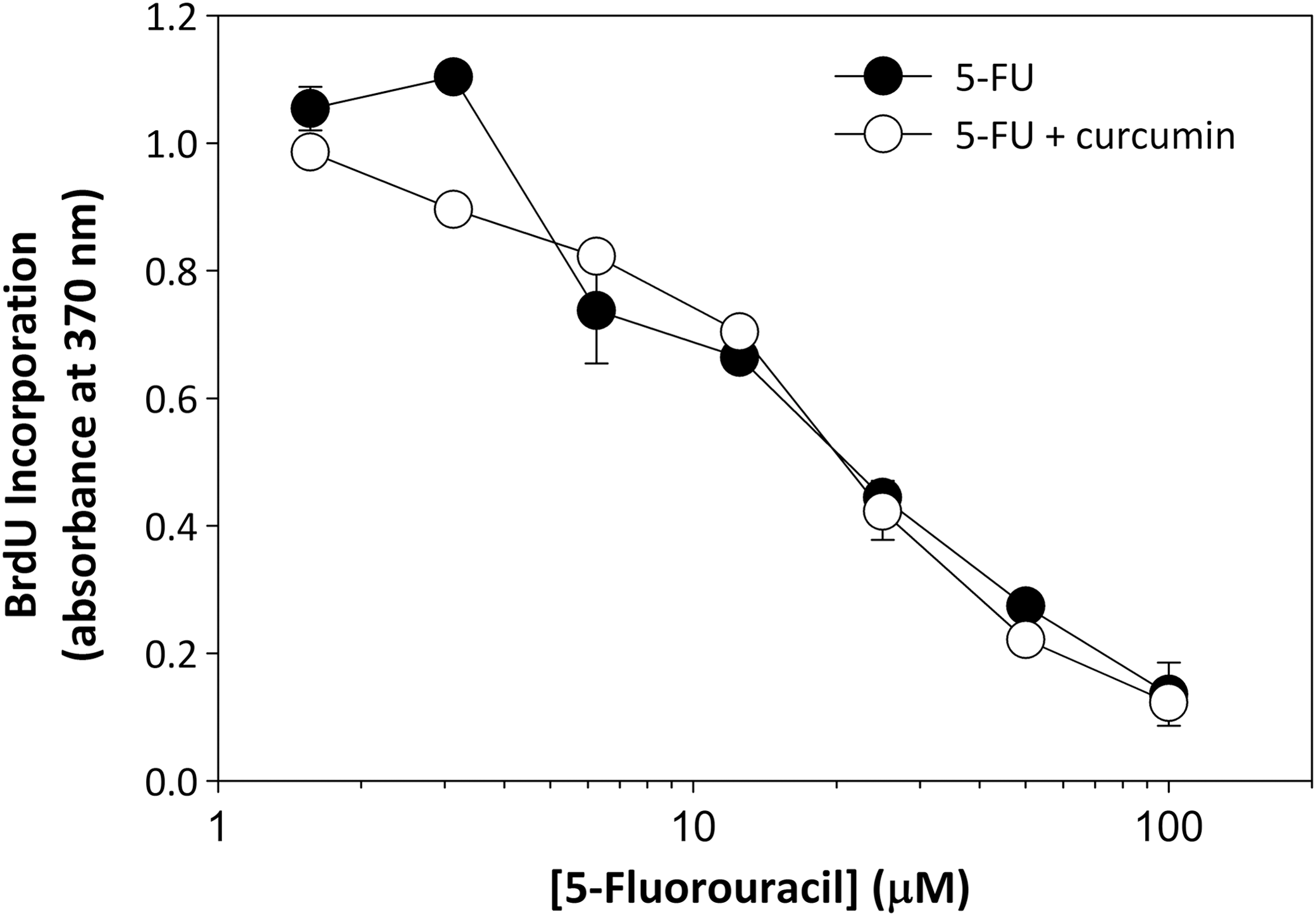
Effect of combined treatment of curcumin and 5-fluorouracil (5-FU) on DNA synthesis. MDA-MB-231 cells were plated at a density of 2000 cells/well into a 96-well plate to stimulate cell division. 5-FU was added to media at concentrations of 1.56, 3.125, 6.25, 12.5, 25, 50, and 100 μM in the presence or absence of 6 μM curcumin. Following a 24-h incubation, BrdU incorporation was quantified by ELISA as in Figure 2. Error bars represent standard deviation of triplicate points. IC50 for 5-FU calculated as 16 μM.
Curcumin is protective against 5-FU cytotoxicity
Besides its antiproliferative activity, curcumin also provides a protective function against both oxidative stress 22 and activation of tumor cell prosurvival signaling pathways. 23 Because of these protective functions, we next examined whether curcumin could reduce the cytotoxic effects of 5-FU, 24,25 such as occurs in individuals with dihydropyrimidine dehydrogenase (DPD) deficiency. 16 We first established the cytotoxicity profile for 5-FU in MDA-MB-231 cells by incubating cells with various concentrations of 5-FU, followed by a measurement of cell viability using the Wst-1 assay. Under these conditions, 5-FU demonstrated an LD50 of 28 μM (Fig. 4). However, when these incubations were repeated with the addition of 6 μM curcumin to each concentration of 5-FU examined, the LD50 value increased substantial to >100 μM. Extrapolation of these data suggests an LD50 value of 200–300 μM, representing a 7–10-fold protection by curcumin against 5-FU cytotoxicity.
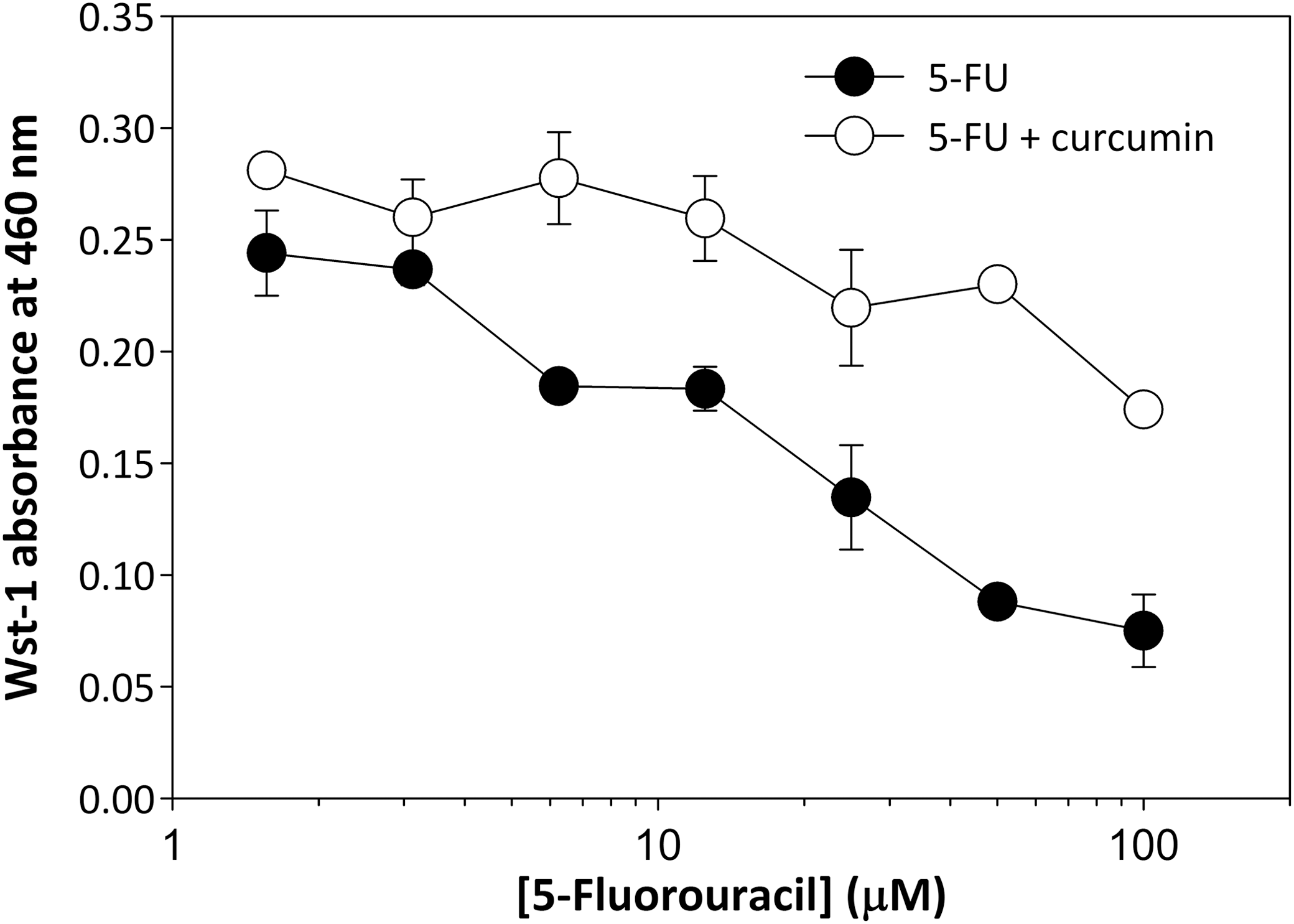
Curcumin protects against 5-FU-induced cellular toxicity. MDA-MB-231 cells were plated at a density of 50,000 cells/well into a 96-well plate. 5-FU was added to media at concentrations listed in Figure 3 in the presence or absence of 6 μM curcumin. Following a 24-h incubation, Wst-1 reagent was added to cells and incubated for 1 h at 37°C. Absorbance was then measured at 460 nm. Error bars represent standard deviation of triplicate points. LD50 for 5-FU in the absence of curcumin, 28 μM; LD50 for 5-FU in the presence of curcumin, 200–300 μM.
Discussion
5-FU has proven to be an effective anticancer agent by functioning as a potent inhibitor of DNA synthesis through its inhibition of thymidylate synthase. Thymidylate synthase catalyzes the conversion of dUMP to dTMP using 5,10-methylene tetrahydrofolate as the methyl donor. 5-FU is converted to its active metabolite, 5-fluoro-dUMP, which binds to the nucleotide-binding site of thymidylate synthase and forms a stable ternary complex with the enzyme and 5,10-methylene tetrahydrofolate. This complex prevents entrance of dUMP into the nucleotide-binding site of the enzyme and inhibits dTMP synthesis resulting in deoxynucleotide pool imbalances inhibiting DNA synthesis.
Although 5-FU is widely used in the treatment of a range of cancers, including colorectal and breast cancers, it demonstrates only modest response rates as a first-line treatment for advanced cancers. 15 To improve upon these results, 5-FU has been combined with newer chemotherapies, such as Irinotecan and Oxaliplatin, resulting in improved response rates up to 40–50%. 15,26,27 Consistent with this rationale, treatment regimens that combine 5-FU and natural products that have shown potential as anticancer agents may also provide improved response rates while limiting undesirable side effects that are typically not associated with natural product compounds. One such natural product is curcumin, a common spice derived from turmeric. Curcumin is a polyphenolic compound that has been shown to inhibit proliferation of many tumor cell lines. In addition, curcumin was reported to inhibit initiation and progression of tumor growth in several murine tissues. 28 –30 The anticancer properties of curcumin are thought to result from its inhibitory activity of a diverse group of intracellular signaling pathways, some of which include cycloxygenase-2, lipooxygenase, ornithine decarboxylase, 31 –33 c-Jun/AP-1, nuclear factor-kappa B (NF-κB), 34,35 c-Jun N-terminal kinase, and protein kinase C. 36,37 The NF-κB signaling pathway is best known for its contributions to gene activation in the innate inflammatory response 38 and is also known to be a major regulatory pathway of breast cancer proliferation. 39 Genes that are constitutively activated by the NF-κB pathway in advanced-stage breast cancers promote the expression of prosurvival signals, such as expression of proteins responsible for extracelluar matrix degradation and cell motility, 40 increased cyclin expression, 39 and other genes responsible for cell proliferation, antiapoptosis, and metastasis. Based on these findings, it is expected that administration of compounds that inhibit the NF-κB pathway should block or at least reduce prosurvival signals and sensitize breast cancer cells to traditional chemotherapy. 41 To explore this possibility, we examined whether treatment of breast cancer cells with combined 5-FU and curcumin would potentiate the antiproliferative activity of 5-FU and improve the therapeutic response of 5-FU. Using a BrdU assay to measure DNA synthesis, curcumin alone did indeed inhibit replication as anticipated based on previous studies; however, we were unable to measure a synergistic response when 5-FU was titrated against curcumin. This was unexpected since a previous study demonstrated synergistic inhibitory effects of curcumin and 5-FU toward proliferation of AGS human gastric cancer cells. 20 In this study, the authors suggest that curcumin inhibits the G2/M transition phase of the cell cycle and provides additional antiproliferative activity along with thymidylate synthase inhibition from 5-FU. Comparing our findings with those from this study clearly indicates that signaling pathways targeted by curcumin in gastric tumors differ from those activated in breast cancers.
A major concern of 5-FU therapy is its often severe cytotoxicity toward normal cellular activities, such as that seen with cardiotoxicity following 5-FU treatment. 42 Although the precise etiology and pathophysiology of 5-FU-induced cardiotoxicity is still unknown, its reported incidence related to 5-FU ranges from 1.2% to 18%. 43 –45 A meta-analysis that involves 1219 patients with colorectal cancer showed that grade 3–4 toxicity was encountered in 31–34% of the patients receiving 5-FU, with 0.5% of the patients experiencing lethal toxicity. 14,25 The catabolic pathway is of particular importance for determining a patient's response to 5-FU, since a reduced activity of enzymes involved in 5-FU catabolism may result in a substantially increased half-life of the drug, and thus, an increased risk of dose-dependent severe toxicity. 12,16,46,47 The biochemical basis of severe 5-FU toxicity has been attributed to impaired drug catabolism, resulting in a markedly prolonged 5-FU plasma half-life and almost complete absence of drug catabolites. 16,48
Genetic factors are thought to account for part of the severe to life-threatening toxicities from 5-FU. Most studies that examine pharmacogenetics of 5-FU have focused on DPD, this being the rate-limiting enzyme that converts 5-FU to 5-fluorodihydrouracil. 24,49 It is reported that 71% of 80 patients with severe 5-FU toxicity showed reduced DPD activity, 50 suggesting that impaired DPD activity is a major contributor to 5-FU-related adverse effects. In agreement with this interpretation, several studies reported promising results of reduced rates of severe toxicities and higher treatment efficacy using pharmacokinetically guided dosing of 5-FU. 51,52
Curcumin is known to have chemoprotective effects for many organs, likely due to its antioxidant properties. 53 Because of these chemoprotective effects, we examined whether a combined treatment of curcumin with 5-FU would protect normal cells against the cytotoxic side effects of 5-FU while maintaining its antiproliferative activity toward tumor cells. If this is the case, then the addition of curcumin as an adjuvant therapy to 5-FU treatment in breast cancer may benefit treatment outcomes by accommodating higher doses of 5-FU and/or longer treatment times. Indeed, we found that curcumin significantly reduced 5-FU-induced cytotoxicity by maintaining cell viability. The mechanism by which curcumin provides protection against 5-FU-induced cytotoxicity requires further investigation and may involve its antioxidant properties or its inhibitory activity toward prosurvival signaling pathways in breast tumors. However, at present these novel findings suggest that the addition of curcumin as an adjuvant therapy during 5-FU treatment might enhance the chemotherapeutic effectiveness of 5-FU by protecting normal cells from reduced viability and thus permitting higher dosing or longer treatment times. This would be especially important to those individuals with DPD deficiencies who are plagued with severe cytotoxicities that require frequent interruptions, or even early termination of their treatment regimens.
Footnotes
Acknowledgment
The authors would like to thank Amanda Gonzales for her expert technical assistance with this study.
Author Disclosure Statement
No competing financial interests exist.