
Abstract
A sapogenin-rich preparation from Jamaican bitter yam (Dioscorea polygonoides) has been shown to reduce blood cholesterol concentrations in hypercholesterolemic rats and mice. Also, diosgenin supplementation has been reported to have antilipemic effects in several animal species. We investigated potential mechanisms of the lipid-lowering actions of bitter yam and also whether the actions were mediated by diosgenin. Sprague-Dawley rats were fed a hypercholesterolemic diet (4% cholesterol) alone or with 5% bitter yam or 1% diosgenin supplementation for 6 weeks. The control group was fed normal rat chow. The serum lipid profile, fecal cholesterol concentration, and serum lipase activity were assessed at the end of the period. The induction of hypercholesterolemia was inhibited by coadministration of 5% bitter yam or 1% diosgenin in the diet. Serum lipid profiles were similar in rats fed bitter yam or diosgenin. The fecal cholesterol concentration was significantly (P<.01) higher in rats fed diosgenin compared to the cholesterol group. However, there was no corresponding elevation in the group fed bitter yam. Administration of bitter yam or diosgenin supplement significantly increased (P<.01) the serum lipase activity compared to the normal control and cholesterol groups. The cholesterol-supplemented diet inhibited normal gain in body weight over the period. This action was potentiated by diosgenin. The effects of the respective supplements on body weight were not completely explained by food consumption. Supplementation of the diet with Jamaican bitter yam may be therapeutically beneficial in the management of hypercholesterolemia.
Background
Y
Diosgenin is the main sapogenin in yam and is common to all members of the Dioscoreaceae family, which contain steroidal saponins. McAnuff et al. 3 reported the presence of Δ 3 diosgenin, β-sitosterol, stigmasterol, diosgenin, and pennogenin in crude bitter yam sapogenin extract, which has some beneficial effects in the management of diabetes mellitus. The consumption of sapogenin extract from bitter yam has been demonstrated to lower blood glucose and lipid concentrations in streptozotocin-induced diabetic rats. 4 Similarly, diosgenin and other sapogenins have been reported to produce lipid-lowering effects in several hypercholesterolemic animal models. 5 –9
It has been proposed that diosgenin produces its antilipemic action through inhibition of gastrointestinal absorption of cholesterol. 5,8,9 Mechanisms suggested for the inhibition of cholesterol absorption by diosgenin include formation of insoluble complexes with cholesterol or changes in the permeability or structure of the intestinal mucosa. 6,10 –12 McAnuff et al. 2 also reported morphological changes in the gastrointestinal tract, including reductions in the intestinal crypt depth, of diabetic rats fed 1% diosgenin for 3 weeks.
There have also been reports of antiobesity effects of diosgenin. 8,13 Reduction in food intake has been suggested as the basis of the inhibitory effects of diosgenin on weight gain in animal models. 9 Another study reported reduced nutrient absorption in rats fed diosgenin, for which the proposed explanation was inhibition of pancreatic lipase. 14 Saponins such as diosgenin-related compounds also reportedly slow the rate of gastric emptying, possibly through reduced activity of gastrointestinal enzymes. 15
We previously reported that a sapogenin-rich preparation from Jamaican bitter yam (D. polygonoides) significantly reduced blood cholesterol in hypercholesterolemic Sprague-Dawley rats. 16 However, the mechanism by which bitter yam lowers blood cholesterol is not known. This study was designed to compare the effects of bitter yam and diosgenin supplements on blood cholesterol levels in a rat model of diet-induced hypercholesterolemia. 16 We investigated the likely mechanisms by which bitter yam produced its antilipemic effects and whether these effects were mediated by diosgenin.
Materials and Methods
Freshly harvested tubers of bitter yam (D. polygonoides) were obtained from the parish of Trelawny, Jamaica. The yam variety was verified (voucher numbers 35620, 35621, and 35622) at the Herbarium, University of the West Indies, Mona, Jamaica. 17 The sapogenin content of the bitter yam was 4.1 mg/g dry weight, of which diosgenin contributed 80%, while pennogenin, Δ 3 diosgenin, stigmasterol, and β-sitosterol accounted for the remainder. 3,16 The tubers were thoroughly washed, sliced, dried under the sun, and pulverized. The bitter yam and diosgenin were supplemented at a level of 5.0 g of bitter yam or 1.0 g of diosgenin/100 g of normal rat chow. 5,16 Cholesterol and diosgenin were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Adult Sprague-Dawley rats (16), obtained from the University of the West Indies Animal House, were divided into four groups by weight (4 rats per group, average body weight 326.6±25.1 g) for a 6-week study excluding a1-week period of acclimatization, during which rats in the four groups were fed with normal rat chow. The experimental groups were as follows: normal rats fed normal diet (normal control), normal rats fed normal diet plus 4% cholesterol (cholesterol supplement), normal rats fed normal diet plus 4% cholesterol plus 5% bitter yam (bitter yam supplement), and normal rats fed normal diet plus 4% cholesterol plus 1% diosgenin (diosgenin supplement). Rat chow (Lab diet 5008; obtained from PMI Nutrition Intl. Inc., St. Louis, MO, USA) was crushed in a laboratory mill for ease of mixing with the other dietary components. The diets were prepared by incorporating the required quantity of cholesterol, bitter yam, or diosgenin with the rat chow to achieve the desired proportions.
Rats were housed in stainless steel cages in a room kept on a 12-h light–12-h dark cycle and allowed to have access to their respective diets and water ad libitum. The cages were cleaned daily. The body weight and total food intake were recorded weekly, and fecal output for each group was collected and weighed daily. The rats were fed their respective diets for 6 weeks and sacrificed by intraperitoneal injection of sodium pentobarbitone (120 mg/kg) at the end of the study. Ethical approval for the study was obtained from the University Hospital of the West Indies/University of the West Indies/Faculty of Medical Sciences Ethics Committee.
Blood samples were collected from the tail veins of rats after an overnight fast at the end of the 6-week treatment period. The lipase activity and concentrations of total cholesterol, high-density lipoprotein (HDL)-cholesterol, and triglycerides (TG) in serum were measured using an automated analyzer (Architect C8000) and Abbott reagents in the Department of Pathology at the University Hospital of the West Indies. The concentration of the combined very low-density lipoprotein (VLDL)+low-density lipoprotein (LDL)-cholesterol component was determined mathematically from the following relationship: VLDL+LDL-cholesterol=total cholesterol−HDL-cholesterol.
Lipids were extracted from feces by a modified version of the method of Folch et al. 18 Samples (2 g) of dried feces were homogenized in a 2:1 ratio of chloroform and methanol. The homogenate was made up to 40 mL with the chloroform–methanol mixture and filtered through a Büchner funnel into a glass vessel. Each sample of crude homogenate was mixed with distilled water (8 mL), and the stirring rod rinsed with a small volume of solvent comprising chloroform, methanol, and water (86:14:1). The mixture was allowed to stand, after which the lower phase containing the lipids was collected and stored in glass tubes. The concentration of total cholesterol in each sample was quantified using the autoanalyzer as was done for serum cholesterol.
Statistical analysis
Data are presented as mean±SEM; n=4. Analysis of variance was used to test for differences among the groups. Tukey's post hoc test was used to locate statistical differences in group means (P<.05).
Results
Figure 1 shows the effects of cholesterol, bitter yam, and diosgenin supplements on the body weight of rats. A steady increase in body weight was noted in the control rats maintained on normal rat chow. However, in general, the body weight of rats fed cholesterol, bitter yam, or diosgenin supplement demonstrated a slight increase up to week 3, at which point there was a significant decline up to week 5. The decrease in body weight was greatest in the group fed diosgenin supplement. Food consumption was highest in the group fed cholesterol supplement and lowest in the groups fed bitter yam or diosgenin supplement; however, the differences were not statistically significant (Table 1).

Percentage change in body weight of rats fed cholesterol, diosgenin, or bitter yam supplement. Graph showing effect of 4% cholesterol diet alone or supplemented with bitter yam (5%) or diosgenin (1%) on the change in body weight of rats over 6 weeks. Values represent mean±SEM; n=4 rats for each group. The symbol “*” represents a significant difference from the control group, whereas “#” represents a significant change in body weight within a group during the 6-week feeding period. Color images available online at
Table showing the average daily food intake relative to body weight, and the weight of feces excreted within 24 h by rats fed cholesterol, bitter yam or diosgenin supplement, or normal chow. Values represent mean±SEM; n=4.
Figure 2 shows the effects of cholesterol, bitter yam, and diosgenin supplements on serum lipid profile in rats. The administration of cholesterol supplement for 6 weeks significantly elevated serum total cholesterol and VLDL+LDL-cholesterol above normal (P<.01). The serum total cholesterol and VLDL+LDL-cholesterol levels were significantly lower in the groups administered bitter yam or diosgenin supplement compared to the group fed cholesterol supplement (P<.05).
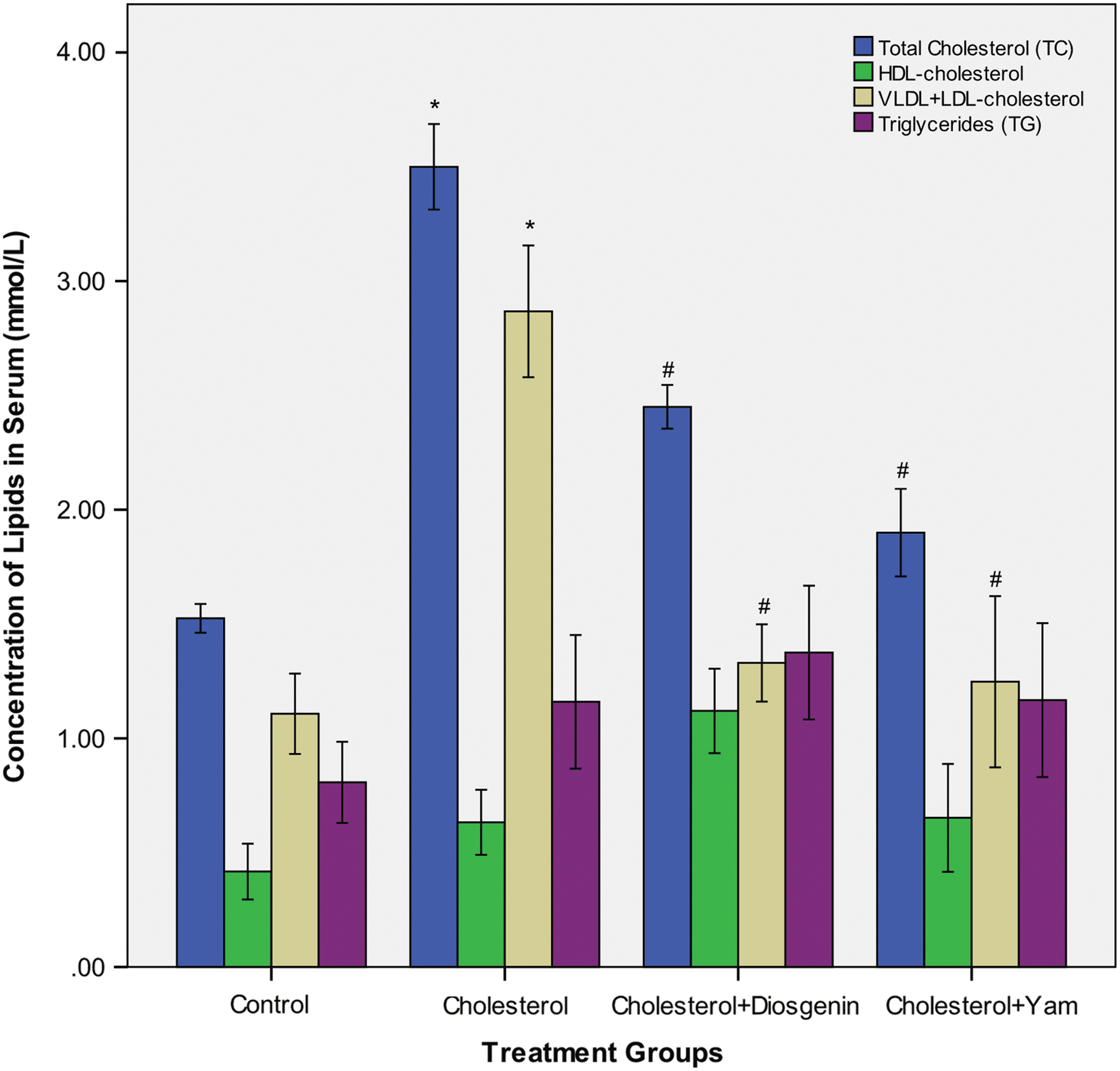
Serum lipid profiles of rats fed cholesterol, diosgenin, or bitter yam supplement for 6 weeks. Graph showing effects of diosgenin and bitter yam on the induction of hypercholesterolemia with dietary cholesterol. The symbol “*” indicates that the mean lipid concentration differs significantly from the control group. The symbol “#” indicates a significant difference in the serum lipid concentration between the groups administered the 4% cholesterol diet supplemented with 5% bitter yam or 1% diosgenin in comparison to the group administered the cholesterol diet alone for 6 weeks. Values represent mean±SEM; n=4. Color images available online at
Figure 3 shows the effects of cholesterol, bitter yam, and diosgenin supplements on fecal cholesterol concentrations in rats. Fecal cholesterol levels were at least 100% higher in the groups fed cholesterol, bitter yam, or diosgenin supplement compared to the normal control rats. The fecal cholesterol level was almost twice as high in the group fed diosgenin supplement compared to the group fed cholesterol supplement (P<.05). However, no significant difference was noted between the groups fed cholesterol or bitter yam supplement. In addition, there was no significant difference in the amount of feces excreted daily by rats among the groups (Table 1).
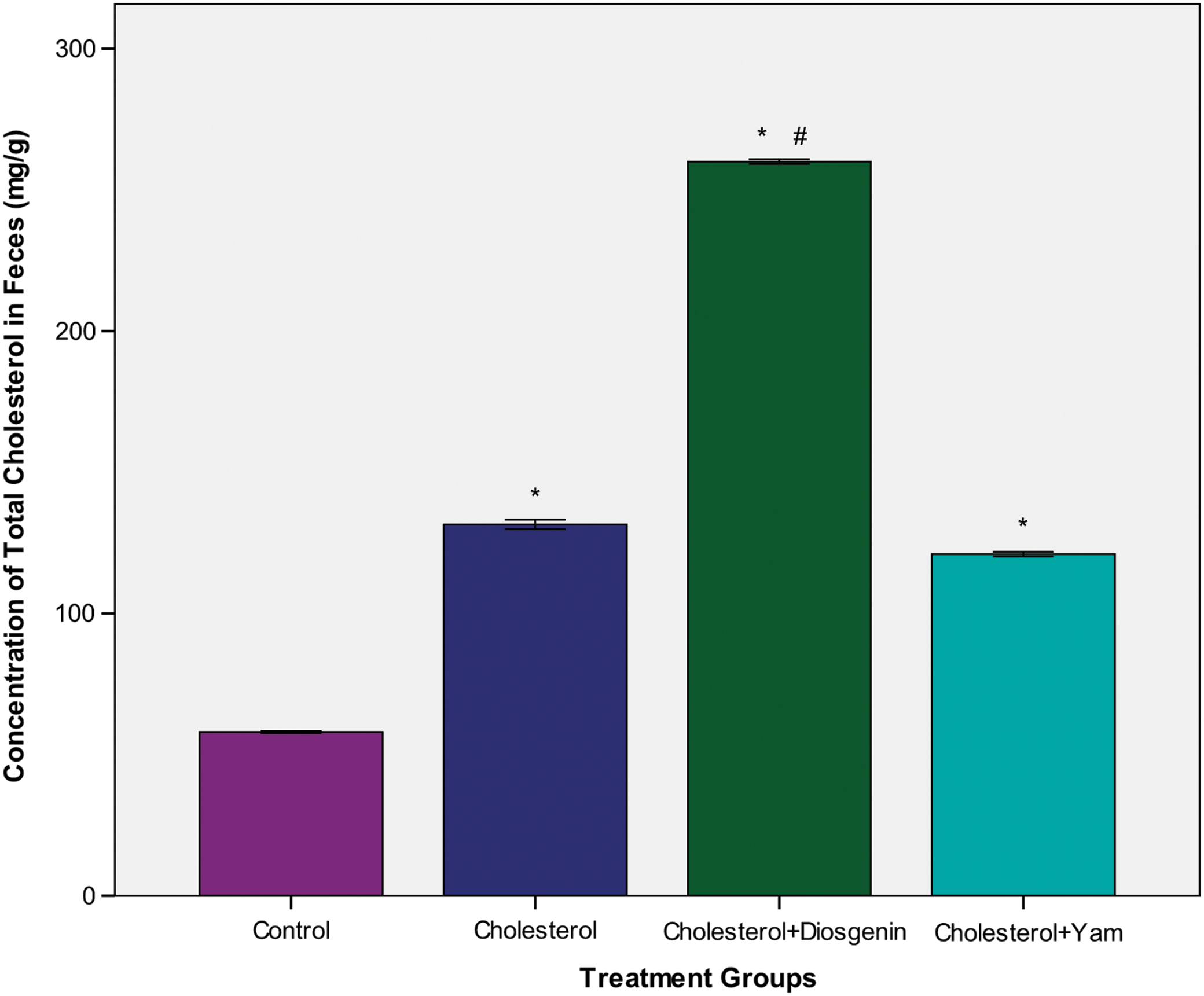
Fecal cholesterol concentration of rats fed cholesterol, diosgenin, or bitter yam supplement for 6 weeks. Graph showing effects of dietary cholesterol, diosgenin, and bitter yam on fecal cholesterol concentration. Values represent means for pooled samples from each group of rats (n=4). The symbol “*” indicates a significant difference from normal control groups. A significant difference between the diosgenin supplement group and the cholesterol group is represented by “#.” Color images available online at
The serum lipase activity was significantly lower (P<.05) in rats fed cholesterol supplement compared to normal control rats. We noted a significantly higher (P<.05) serum lipase activity in the groups fed bitter yam or diosgenin supplement compared to the normal control and cholesterol supplement groups (Fig. 4).
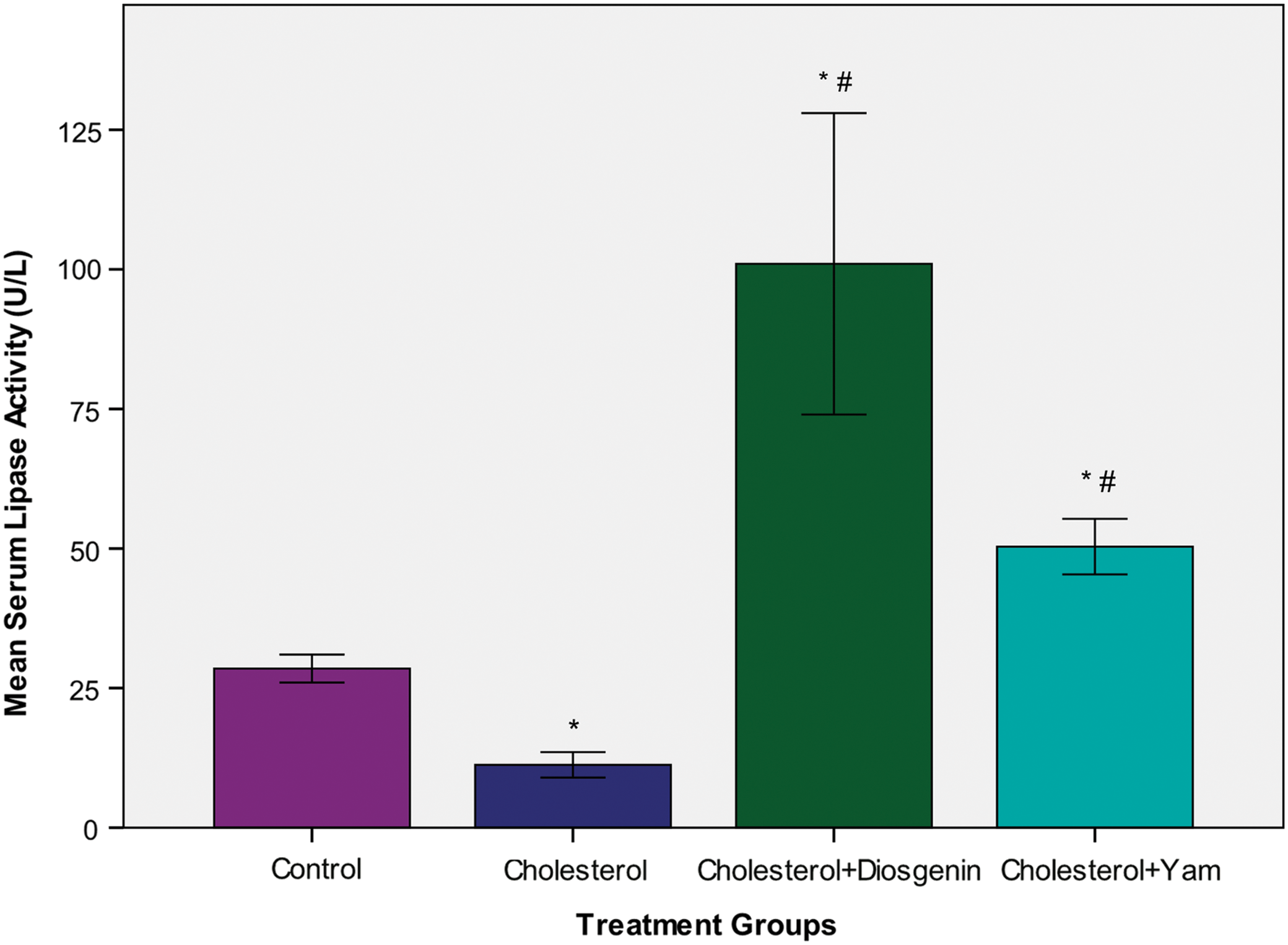
Serum lipase activity of rats fed cholesterol, diosgenin, or bitter yam supplement for 6 weeks. Graph showing effect of 4% dietary cholesterol alone or with 5% bitter yam or 1% diosgenin on activity of serum lipase. Values are group mean±SEM; n=4. The symbol “*” indicates a significant difference from the control group, whereas “#” indicates a significant difference between the groups administered cholesterol with bitter yam or diosgenin as compared to cholesterol alone for 6 weeks. Color images available online at
Discussion
The decrease in body weight observed in Sprague-Dawley rats, fed the high cholesterol diet in the present study, is supported by comparable findings in Wistar and Fischer rats fed hypercholesterolemic diets. 19,20 It was suggested that the higher caloric value of the hypercholesterolemic diets may have reduced food intake or that the absorption of nutrients may have been impaired by the higher fat content of hypercholesterolemic diets. Inhibition of nutrient absorption may be a reasonable explanation for the effects of dietary cholesterol on body weight of rats in the present study.
The addition of bitter yam or diosgenin to the hypercholesterolemic diet produced further inhibitory effects on body weight. The reduction in food intake is, however, not entirely supported by the results, as the food intake was almost identical in the groups administered bitter yam or diosgenin although the inhibitory effect on body weight was more pronounced in the diosgenin group. It was previously reported that dietary diosgenin reduced intestinal crypt depth in diabetic rats. 2 It is suggested that the reduction in the intestinal surface area may have decreased nutrient absorption and inhibited weight gain in rats fed diosgenin in the present study. The inhibitory effects of bitter yam on normal body weight may partly be attributable to its diosgenin component.
Bitter yam and diosgenin may also have high satiating and appetite suppressant properties. A blueberry extract was shown to reduce the food intake and body weight in rats by its high satiating effects, although the specific underlying mechanism was not presented. 21 Blueberries also reportedly demonstrated antioxidant effects, reduced the accumulation of fat in the aortas and livers of animal models, and reduced cardiovascular risk in obese individuals. 22 –25 It is possible that the biological effects were produced by phytosterols, phenolics, and other bioactive compounds present in blueberries as in bitter yam. 2,16,26 Saponin-induced delay in gastric emptying 15 or other mechanisms of satiating effect may have suppressed the appetite and slightly reduced food intake in rats fed 5% bitter yam or 1% diosgenin supplement in the present study.
The negative effect of the high cholesterol diet on blood lipid profile was inhibited by the supplementation of the hypercholesterolemic diet with 5% bitter yam or 1% diosgenin. The significantly higher concentration of cholesterol in the feces of rats administered diosgenin with cholesterol, in comparison to those administered the cholesterol diet alone, suggests that diosgenin inhibits the gastrointestinal absorption of cholesterol. This increase in fecal cholesterol excretion was, however, not observed with bitter yam, although the total fecal output was similar in the groups administered cholesterol with 5% bitter yam or 1% diosgenin. It is suggested that diosgenin inhibited the gastrointestinal absorption of dietary cholesterol by reducing the intestinal surface area, consequently increasing fecal elimination of cholesterol. Bitter yam, unlike diosgenin, was not shown to significantly alter intestinal morphology in rats. 2 This could account for the absence of significant differences in fecal cholesterol concentrations between rats administered the 4% cholesterol diet alone or with 5% bitter yam in the present study.
The serum lipase activity was highest in the groups administered 5% bitter yam or 1% diosgenin with the hypercholesterolemic diet and lowest in the group administered the high cholesterol diet alone. It is suggested that the cholesterol diet reduced the serum lipase activity, thereby decreasing hydrolysis of TG. The TG would then be available for incorporation into VLDL and LDL, contributing to higher levels of these lipoproteins in rats administered the 4% cholesterol diet alone. It is further proposed that bitter yam and diosgenin inhibited the dietary cholesterol-induced reduction in serum lipase activity, thus increasing hydrolysis of TG to release fatty acids. This would decrease the serum concentrations of LDL and VLDL as evidenced by the results of a lower combined VLDL+LDL-cholesterol component of blood lipids in rats administered bitter yam or diosgenin with dietary cholesterol.
Pancreatic lipase was reportedly inhibited by the powder from a yam variety (Dioscorea nipponica Makino) grown in Korea. 14 Those researchers proposed that the effects of the yam were mediated by its diosgenin component although only the crude extract of the yam was investigated in the study. The results appear contradictory to those of the present study; however, it should be noted that the yam varieties were different and could contain different constituents. Additionally, the present study used in vivo methods of investigation, whereas in vitro techniques were used by the other researchers. The measurements in the study with Dioscorea nipponica were also reportedly specific for pancreatic lipase, whereas the enzyme specificity may have been lower in the present study. Serum lipase is expected to be predominantly pancreatic lipase 27 ; however, it is suggested that the measurements of serum lipase in the present study may also have included the related lipoprotein lipase, which hydrolyzes protein-bound TG.
The mechanism of action of commercially available fibric acid derivatives includes stimulation of peroxisome proliferator-activated receptor alpha (PPARα) and an increase in the activity of lipoprotein lipase, most significantly in atherogenic dyslipidemias characterized by low blood HDL-cholesterol and elevated TG concentrations. 28 –34 As VLDL+LDL-cholesterol was most significantly elevated in the rat model of hypercholesterolemia used in the present study, it is probable that the elevation in the serum lipase activity and subsequent increase in hydrolysis of TG has a contributory role but may not be the primary antilipemic mechanism of bitter yam and diosgenin. An alternative suggestion is that the lipoprotein lipase activity may have more significant implications for hypercholesterolemia in some animal species. Overexpression of lipoprotein lipase reportedly protected transgenic mice and rabbits from developing dietary hypertriglyceridemia and hypercholesterolemia and also prevented excessive weight gain. 35 –37 It is therefore possible that a higher serum lipoprotein lipase activity prevented the induction of hypercholesterolemia and contributed to inhibition of weight gain in rats fed bitter yam or diosgenin with the high cholesterol diet in the present study.
The effects of bitter yam on serum lipase activity are likely attributable to the diosgenin component of the yam; however, the disparity in results with fecal cholesterol concentrations indicates that the antilipemic effects of bitter yam are not mediated solely by diosgenin. The bitter yam powder used in this study contained other constituents, including pennogenin and the phytosterols, β-sitosterol and stigmasterol. 3 Phytosterols reportedly reduce blood cholesterol concentrations by inhibiting cholesterol absorption from the diet or bile. 38 It is suggested that the phytosterols in bitter yam may have been incorporated in the intestinal mucosa and interacted with dietary cholesterol, 39 thus reducing its gastrointestinal absorption and also its elimination in feces. This likely contributed to the results for serum and fecal cholesterol concentrations in rats administered bitter yam with the hypercholesterolemic diet.
Another possible difference in antilipemic mechanisms of bitter yam and diosgenin may involve bile acids. Diosgenin does not bind bile acids 5 ; however, some saponins reportedly interact with bile acids to form stable species. 40 It is theorized that in contrast to the diosgenin, other saponins in bitter yam may have formed insoluble complexes with bile acids, thus reducing blood cholesterol concentrations in a manner similar to bile acid sequestrants used in the management of hyperlipidemias. 41,42 The concentration of serum cholesterol could thus be regulated by bitter yam without a significant increase in fecal cholesterol concentrations.
In conclusion, the administration of 1% diosgenin or 5% bitter yam demonstrates similar inhibitory potential against dietary-induced hypercholesterolemia; however, the antihyperlipidemic mechanisms are not identical, especially in relation to the elimination of cholesterol in feces. The antilipemic actions of Jamaican bitter yam appear to be mediated by the combined effects of diosgenin and other components of the yam acting by various mechanisms, including the increased serum lipase activity.
Ethical Considerations for Conduct of Research
All institutional and national guidelines for the care and use of laboratory animals were followed.
Footnotes
Acknowledgments
The authors thank Everton Thomas, Jevan Howden, and Kevin Grant for providing technical assistance.
Author Disclosure Statement
Dr. Helen Asemota currently holds a patent on the proprietary preparation of bitter yam for therapeutic purposes. The other authors (Marsha-Lyn McKoy, Peta-Gaye Thomas, Felix Omoruyi, and Oswald Simon) have no competing financial interests.