
Abstract
Previous works on staghorn sumac (Rhus hirta) were mostly dedicated to its phytochemical profiles, antioxidant properties, and antidiabetic potentials. This study explored the potential of staghorn-sumac-derived functional ingredients for food and pharmacological applications. Sumac may have other biological functions, such as inhibitory effect on cancerous cells independent of its antioxidant properties. We characterized sumac and raspberry interactions, and their antioxidant capacities (ACs) and their inhibitory effect on both normal and cancerous cells. Mixing sumac and raspberry extracts yielded significantly higher ACs than the sum of sumac and raspberry as evaluated by three in vitro AC assays. However, the potential use of staghorn sumac as a natural source of dietary antioxidant supplement for oxidative-stress-related disorders might be challenged by its cytotoxicity in culturing normal cells. Remarkably, mixing sumac and raspberry showed maximal inhibition of the growth of both rat colon and human breast cancer cells with relatively low cytotoxicity toward normal rat colon and human breast epithelial cells, as compared with sumac or raspberry treatment alone. Sumac-derived products and their synergistic interactions with other food ingredients have great promise as functional food or nutraceutical products that would target cancer cells with minimal toxic effects to normal cells.
Introduction
A
Sumac belongs to the genus Rhus of the Anacardiaceae family. Approximately 250 species of flowering sumac shrubs and small trees are growing worldwide. 5 Diverse health beneficial activities have been demonstrated for sumac extracts, including antioxidant, antitumorigenic, antimutagenic, antimicrobial, cytotoxic, and anti-inflammatory effects. 6 –9 Crude ethanol extracts from Rhus verniciflua exhibited strong antioxidant and cytotoxic effects on mouse thymocytes. 10 A chloroform–methanol fraction from a crude acetone extract of the same sumac species exhibited selective growth inhibition and apoptosis-inducing effects in mouse tumorigenic hepatic cells. 11 These previous works have focused only on a few members of this large family of sumac. Most of the previously published works on other species, in particular, the staghorn sumac (Rhus hirta), were related to phytochemical profile, their antioxidant properties, and antidiabetic potentials. 5,12 –14 Most recently, LC-ESI-MS analysis revealed that staghorn sumac fruits process five unique anthocyanins with aglycones (anthocyanidins), which were structurally different from those of commercially available anthocyanin-rich fruits like raspberry (Rubus idaeus L.), namely, peonidin 3-O-galloyl-hexose, (449 Da)-glucoside-3-O-galloyl-hexose, (419 Da)-hexose-3-O-galloyl-hexose, (433 Da)-glucoside-3-O-galloyl-hexose, and (433 Da)-3-O-galloyl-hexose. 13 As generally recognized as safe (GRAS), multitude of health benefits and functionalities of raspberry and its associated anthocyanins have been inventively reported. 15 –17 Due to possible compensatory and multisite targeting role of these structurally diverse anthocyanins, a mixture of staghorn sumac and raspberry might be valuable ingredients of high quality and biological significance than staghorn sumac or raspberry alone. The interactions between staghorn sumac and raspberries, however, have not yet extensively been investigated.
There are increasing interests in developing effective prevention/treatment strategies using natural food sources to circumvent some challenges encountered with chemotherapeutic drugs in cancer treatment. Some natural polyphenolic components or mixtures of polyphenolics were found to be detrimental to certain types of cancerous cells, but marginally cytostatic toward normal cells. 18 Little information is available regarding specific combinations that have beneficial combinatorial effects in selective cytotoxicity against cancerous cells. Here we were practically interested in the combinatorial effects derived from sumac and raspberry mixture.
To explore the potentials of sumac and sumac/raspberry mixture as functional ingredients for food and pharmaceutical applications, we quantified the total phenolic content (TPC) and ACs of staghorn sumac and raspberry individually and in combination by using an in vitro chemical model system, comprising TPC assays and three different AC assays (ferric-reducing antioxidant power [FRAP], 2,2-diphenyl-1-picrylhydrazyl [DPPH] radical-scavenging capacity, and oxygen radical absorbance capacity [ORAC]). Plant extract and mixtures with synergistic effects observed in this in vitro model system have been confirmed to have protective property against reactive-oxygen-species-induced cell death in biological systems. 3 We also evaluated the selective antiproliferative activity individually and in combination of extracts in two paired cell lines. One pair was normal rat colonic epithelial cells (4D/WT) and transformed colonic epithelial cells (D/V-Src). Another pair was human spontaneously immortalized breast epithelial cell line (MCF-10A) and breast cancer cell line (MCF-7). The cytotoxicity of sumac was investigated in the cultured RAT-2 and C2C12 cell lines. Most of the investigations on the effect of polyphenols on cancer cell proliferation have been conducted with pure or isolated polyphenolic compounds (i.e., flavonoid). This is one of a few studies where crude extract containing complex mixture of polyphenols and other phytochemical species was used to treat the cells for evaluating cell survival and cell death. This study may contribute to the development of a low-cost and clinically applicable dietary combination(s) with enhanced antioxidant and/or cancer-fighting properties.
Materials and Methods
Chemicals and reagents
All chemicals were obtained from Sigma-Aldrich (Oakville, ON, Canada) except ferric chloride and sodium acetate (Aldrich Chemical Co. Milwaukee, WI, USA), and ascorbic acid, 2,4,6-tripyridyl-s-triazine (TPTZ), and the Folin-Ciocalteu reagent (Sigma Chemical Co., St. Louis, MO, USA). Alpha-minimal essential medium (alpha-MEM) and Dulbecco's modified Eagle's medium (DMEM) were from Cedarlane Laboratories (Burlington, ON, Canada); fetal bovine serum (FBS) was from Thermo Scientific Hyclone (Logan, UT, USA). Penicillin and streptomycin were from Invitrogen (Burlington, ON, Canada) and insulin from Calbiochem (La Jolla, Canada). All solvents were HPLC grade and purchased from Caledon Laboratories Ltd. (Georgetown, ON, Canada).
Sample collection and extraction
Fresh R. hirta fruits were harvested at two different growth periods in September and November, 2011 (Guelph, ON, Canada). Fresh, commercially available raspberry (Rubus strigosus) (600 g/kg moisture content) was purchased from a local grocery store (Metro, Guelph, ON, Canada; May–August, 2011). Preparation, extraction procedures, and moisture content determinations were similar to those reported in our previous publication. 1 All extracted samples were stored at 4°C before analysis within 2 days.
TPC and AC evaluation
Methanol (800 mL/L) extracts of sumac and raspberry were evaluated individually and in combination using four in vitro assays, as reported previously. 1 They are TPC, 19 FRAP, 20 DPPH radical-scavenging capacity, 21 and ORAC. 22 Fresh samples were used in this particular study. Data are expressed in fresh weight of raspberry and sumac. The TPC values were expressed as micrograms of gallic acid equivalent per gram of sample. The FRAP value of the samples was calculated on the basis of 500 μM ascorbic acid being equivalent to a 1000 μM FRAP value. The radical-scavenging activity of each sample was expressed as EC50 (concentration of sample extract necessary to scavenge initial concentration of free-radical DPPH 50%), which is defined as the concentration of sample that decreased the initial DPPH radical concentration by 50%. The ORAC values were expressed as micromoles of trolox equivalent per gram of fresh samples. Eighty percent methanol was selected by preliminary studies for ensuring the yields of highest antioxidant potential. The methanol extracts of sumac and raspberry were mixed in pairs at 1:1 (v/v) ratio. For assays of all individual extracts, they were diluted twofold with 800 mL/L methanol before testing, so that their final concentrations were the same as those in their respective paired mixtures. The observed and expected ACs are defined and compared as in our previous report. 1 Specifically, the observed AC of each mixture was compared with the expected value, which is the mathematical sum of the AC derived from the individual extracts analyzed at twofold dilution. Therefore, all comparisons were based on the same total weight of the pair; for example, the AC (observed value) of a 1 g mixture of sumac and raspberry (0.5 g each when mixed at 1:1 v/v ratio) was compared with the mathematical sum of the AC (expected value) of 0.5 g of sumac and that of 0.5 g of raspberry (twofold dilution). If the observed values were significantly higher than the expected value derived from of the same pair of individual foods (P<.05), then a synergistic interaction was considered to have occurred in the mixture. The opposite result, meaning that the observed value was significantly lower than the expected value, was defined as an antagonistic interaction. No significant difference between the two values indicated an additive interaction. The approach was used for all assays except for the DPPH assay. If the observed DPPH values were the same, lower, or higher than the expected DPPH values, then they were considered as additive, synergistic, and antagonistic interactions, respectively, because a higher DPPH value (EC50) represents a lower AC. All tests were done in triplicate.
Cell lines and culture conditions
Rat fibroblast cell line (RAT-2), mouse myoblast cell line (C2C12), human spontaneously immortalized breast epithelial cell line (MCF-10A), and breast cancer cell line (MCF-7) were from the American Type Culture Collection (ATCC; Cedarlane Laboratories). Rat normal colonic epithelial cells (4D/WT) and transformed colonic epithelial cells (D/V-Src) were generous gifts from Dr. Kelly Meckling (University of Guelph, Canada). Nontumorigenic MCF-10A and 4D/WT cells are defined as normal cells in this particular study, as described previously. 23 –25
The RAT-2 and C2C12 cells were cultured separately in DMEM with 10% FBS. The MCF-10A cells were cultured in 50:50 DMEM and HAM F12 nutrient mixture supplemented with 5% horse serum, 10 μg/mL insulin, 0.5 μg/mL hydrocortisone, 20 ng/mL epidermal growth factor, and 100 ng/mL choleratoxin. The MCF-7 breast cancer cells were cultured in alpha-MEM. The medium was supplemented with 10% FBS, 1 mM sodium pyruvate, 50 U/mL penicillin, 50 ng/mL streptomycin, and 10 mg/mL insulin. The 4D/WT and D/V-Src cells were cultured separately in DMEM with 10% FBS, 0.02 μg/mL hydrocortisone, 0.25 μg/mL insulin, 0.12 μg/mL transferrin, 1 μg/mL polybrene, and 50 U/mL penicillin-streptomycin.
All cells were grown at 37°C in a relative humidity of 95% in an atmosphere of 5% CO2 in air, except 4D/WT and D/V-Src cells that were grown in a humidified environment at 37°C in the presence of 10% CO2. All cells were maintained by trypsinization and transferring into fresh medium at appropriate intervals. Growth, division, and morphology of the cells were monitored periodically. Before each experiment, cells were seeded in 96-well plates at a density of 6×103–7×103 cells/well. All extracts were added with concentrations ranging from 0.1 to 100 mg/mL as previously reported 25 to ensure that the comparison among individual studies was meaningful. All extracts were diluted with cell culture medium to give a final methanol concentration of <5 mL/L.
Cell viability and cellular gelatinase activity
For cell viability, equal numbers of myoblast RAT-2 and fibroblast C2C12 were loaded onto a 60-mm culture plate and cultured in DMEM with 10% FBS under 5% CO2 overnight. Different concentrations of the sumac extract were then added and the cells were cultured for an additional 24 h. The cell viability was counted with a hemocytometer. Cellular gelatinase activity was measured as previously described. 26 RAT-2 and fibroblast C2C12 and fibroblast cells were loaded onto the glass cover slip and grown in a 60-mm culture plate overnight. The cells were then grown for an additional 24 h with or without 5 mL of the crude extract. The highly quenched gelatine was diluted in 1:10 in 0.5% agarose and loaded onto the cells growing on the glass slip and then covered with glass slide. The slide was then incubated in a Tris incubation buffer (Molecular Probe) at 37°C overnight. The cellular gelatinase activity and cell morphology were examined under a fluorescence microscope.
Cell proliferation assays by sulforhodamine dye binding method
Cell proliferation assays were performed by the sulforhodamine dye binding (SRB) method as previously described. 27 The highest concentration of methanol used did not exceed 0.5% (v/v) of the total assay volume and a vehicle blank was performed and found to be nontoxic to the cells. Serial dilutions of extracts and their combinations were made in 96-well plates with preseeded cells as previously described. The plates were incubated at 37°C for 1 day. Cells were fixed by gently layering 25 μL of cold 50% trichloroacetic acid (TCA) (4°C) on top of the growth medium in each well. Plates were incubated at 4°C for 1 h and fixed cells were washed five times with deionized water to remove TCA, growth medium, low-molecular-weight metabolites, and unprecipitated proteins. Plates were air-dried and 50 μL of 0.4% sulforhodamine B in 1% acetic acid was added to each well. Plates were incubated for 30 min at room temperature, washed six times with 1% acetic acid, and air-dried. The dye was solubilized by the addition of 100 μL of 10 mM unbuffered tris base [tris(hydroxymethyl)aminomethane] (∼pH 10.5) and shaking plates on a rotary shaker. Absorbance at 570 nm was recorded on Powerwave XS2 plate reader (Biotek, Winooski, VT, USA).
Statistical analysis
All analyses were in triplicate. The data were expressed as mean±standard deviation. All statistical analyses were performed with Statistix for Windows version 9.0 (Analytical Software, Tallahassee, FL, USA). The effect of treatment was determined by a one-way ANOVA and Student's t-test was used for comparing the means of two samples with a significance level of P<.05.
Results and Discussion
Antioxidant synergism derived from sumac and raspberry mixture
Dietary plant materials are touted as being good sources of dietary antioxidants. Complex interactions might occur among the different dietary antioxidants when two plant materials are mixed. Total AC of the mixture might be increased or decreased in synergistic or antagonistic manner as compared with the individuals. 1 The health benefits derived from these antioxidant-rich foods could therefore depend on the nature of these interactions. 1
In this study, 80% methanol extracts of raspberry and sumac were evaluated for their TPC and ACs individually and then in combination. As presented in Figure 1, the mixture of sumac and raspberry had a much greater TPC value and AC than the mathematical sum of the TPC values of the sumac and raspberry extracts analyzed at twofold dilution. Such synergistic phenomena (as defined in “Materials and Methods” section of this study) could be due to the fact that the addition of raspberry extract to sumac extract increases the solubility of phenolic that was either in raspberry extract or sumac extract. 28,29 It should also be noted that the synergistic antioxidant response in the mixtures may not necessarily only arise from the phenolic compounds interacting with each other, 1,30 but possibly from interactions with other phytochemicals. It has been reported that no synergistic phenomena were found in a combination of pure phenolic compounds commonly found in fruits and vegetables. 30 Further studies on the mechanisms of the synergistic interaction of raspberry and sumac will need to be carried out, particularly on other phytochemical species. Additional chemical assays will be expected to provide some insights into the nature of sumac and raspberry interactions. These types of assays will help answer the following questions: (1) Are the observed synergistic interactions related to phytochemical composition of sumac and raspberry? This would then permit the formulation of sumac- and raspberry-derived phytochemical cocktails. (2) Are there other measurable indicators that will predict interaction between sumac and raspberry? Oxidation-reduction (redox) potential could be one such indicator (e.g., redox potential is a measure of the tendency of system to either gain or lose electrons). If appropriate indicators are selected, then this would permit the development of assay to rapidly detect important synergisms.
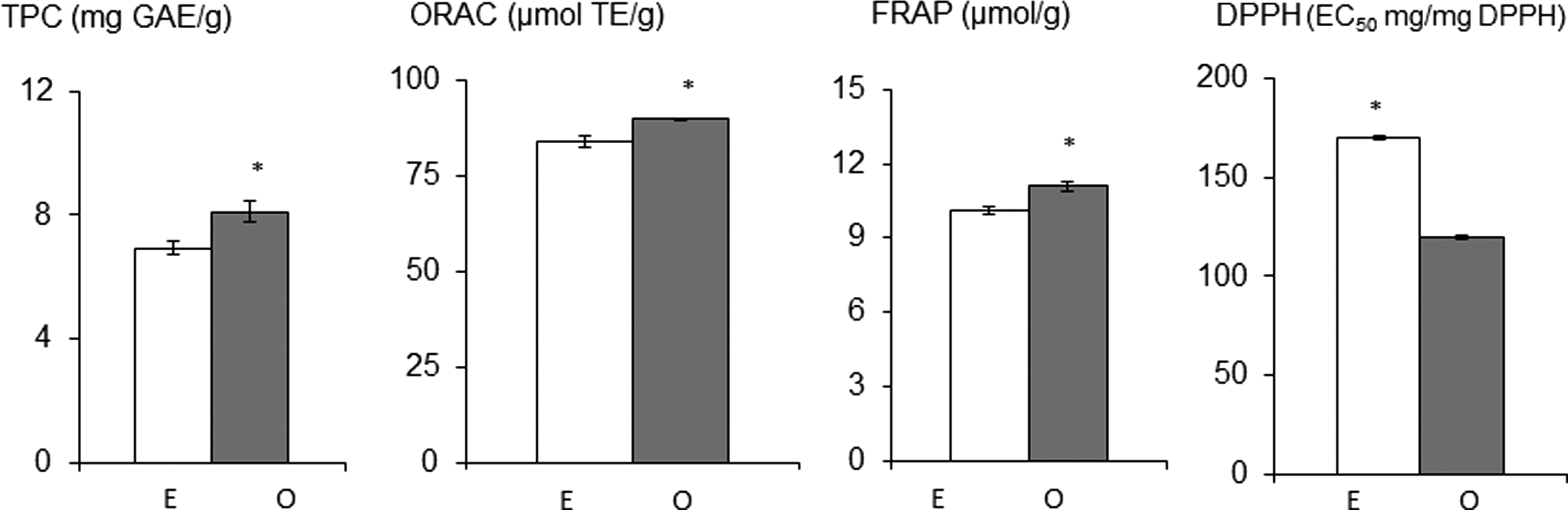
Comparison of phenolic content and antioxidant capacities (ACs) derived from the mixture of sumac and raspberry and a mathematical sum of values of sumac and raspberry. E, expected value: a mathematical sum of the ACs derived from the individual foods in the mixtures at twofold dilution, and O, observed value. In each assay, means followed by * indicate significant difference between expected and observed values (P<.05). TPC, total phenolic content assay; ORAC, oxygen radical absorbance capacity assay; FRAP, ferric-reducing antioxidant power assay; DPPH, DPPH radical absorbance capacity assay. For DPPH assay, lower values indicate higher AC.
It has not been reported elsewhere that an antioxidant synergism was derived from a mixture raspberry and sumac. Previous reports showed the ACs of raspberries or sumac alone. 5,13 Mixtures exhibiting synergistic interactions are appreciated. In antioxidant synergisms, different antioxidants might employ complementary mechanisms to promote an antioxidant state in cells. In our previous studies on antioxidant synergism (a mixture of raspberries and adzuki beans), which was selected from 55 plant-food-based combinations, 1 this antioxidant synergism protected the cardiomyocytes from H2O2-induced cell damage significantly better than the raspberry and adzuki bean treatment alone. 3 Exogenous supplementation of synergisms derived from a mixture of sumac and raspberries might be potentially used as an antioxidant supplement for oxidative-stress-related disorders.
In vitro cytotoxic activity of sumac extracts in RAT-2 and C2C12 cells
Although staghorn sumac has been documented to possess antidiabetic and antioxidant potentials, 5,13 unlike GRAS status of raspberries, the toxicity of staghorn sumac needs to be determined prior to its safe use in vivo. Here, cell viability experiments were carried out to determine whether the sumac extract possesses the cytotoxic or cytostatic effects on normal cell growth. RAT-2 and C2C12 cell lines were treated with different concentrations of sumac extracts (0–100 mg/mL fresh weight [FW]). Sumac extract was slightly toxic to these two cell lines (C2C12 and RAT-2) at a concentration ranged from 0 to 10 mg/mL FW (Fig. 2). Different cells had different toxicity tolerance levels and the viability of cells in culture started decreasing significantly at a concentration of 10 mg/mL sumac extract. Treating C2C12 and RAT-2 cells with sumac extracts resulted in a dose-dependent inhibition of cell proliferation, and the relative inhibitory effects of sumac on the two cell lines were different, especially at concentrations between 10 and 100 mg/mL (Fig. 2). Fibroblast cells appeared to be the most susceptible to inhibition, especially at the concentration of 50 mg/mL, where 50% of cell growth was inhibited by sumac treatment (Fig. 2). This finding is in agreement with another investigation, where the C2C12 cells were more susceptible to the cytotoxicity induced by polyphenolic-rich plant extracts than the mouse embryonic fibroblast 3T3-L1 cells. 23

The effects of the 80% sumac extracts on the cell variability of myoblasts and fibroblasts. Data are presented as the mean of three or more independent experiments±standard error. For each concentration, * indicates that the relative inhibitory effects of sumac on the two cell lines are significantly different (P<.05). The 100% cell control represents 7×103 to 8×103 cells.
The nature of growth inhibition observed when cells were treated with the sumac extract was further investigated through the impact of sumac extracts on the activities of total gelatinase (TG) secreted by the cultured fibroblasts and myoblasts. Gelatinase is mostly to play a role in the disruptions of cell homeostasis and cell death in response to the external stimulation. 26 The morphology of the treated RAT-2 and C2C12 cells was shown in the Figure 3. Both cells treated with the extracts showed decrease in cellular gelatinase activity as evidenced by dimmed fluorescence (Fig. 3). As well, treated cells also showed deformed cytoplasm as indicated by the morphological changes, especially in those treated with higher concentrations (Fig. 3). Again, RAT-2 and C2C12 cells show different sensitivity to sumac extracts, as indicated by changes in TG and morphology (Fig. 3). This finding is consistent with another reporter, who pointed out that the TG responded differently to mechanical stimulation in fibroblasts and myoblasts. The mechanisms behind the observed cytotoxicity of the sumac extracts are still to be explored.
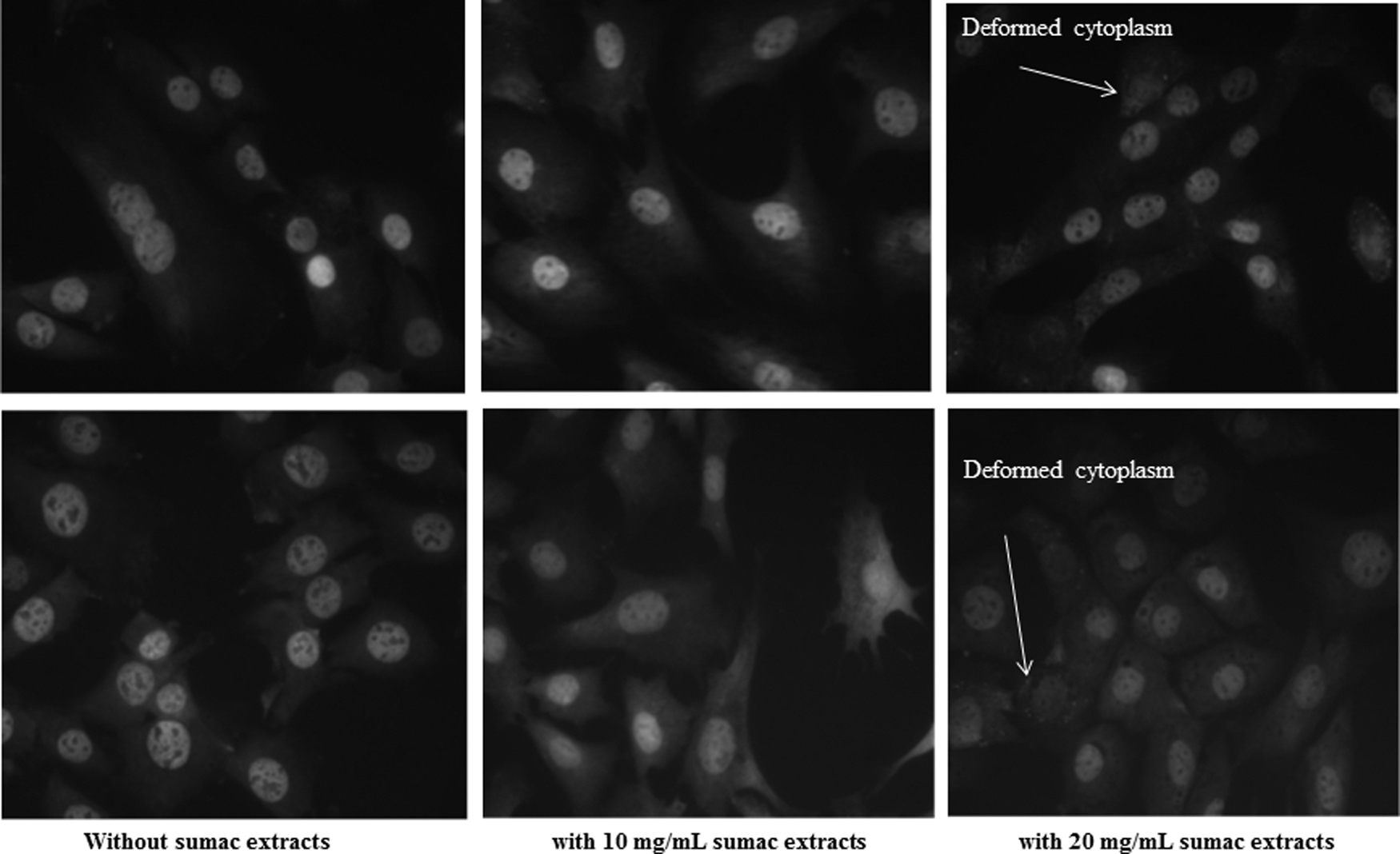
The effect of sumac extracts on gelatinase activity and morphology of myoblast (the top row) and fibroblast (the lower row) cells. Fibroblasts (RAT-2) and myoblasts (C2C12) were cultured with or without sumac extracts for 24 h and applied with a biaxial deformation (15% magnitude, 1 Hz) using the Flexcell FX-4000 system.
The cytotoxicity of sumac was observed in the cultured RAT-2 and C2C12 cell lines; the use of staghorn sumac as a natural source of dietary antioxidants should thus be with caution. On the other hand, it is widely recognized that identification of cytotoxic compounds might direct the discovery process of conventional anticancer agents. Additional biological functions (e.g., antitumor activity) of edible-plant-based materials (e.g., sumac) might go beyond their ACs. 1 The intention in our next approach was therefore to identify the chemotherapeutic potential of staghorn sumac.
Inhibitory effects of sumac, raspberry, and their mixture on normal and cancerous cells
Here, colon and breast cell lines were of our particular interest. Colorectal cancer and breast cancer are the two most prevalent types of cancers in North America. Early detection and chemotherapy have only modestly improved the survival rate of patients. Combinations of different foods may produce additive, synergistic, or antagonistic interactions that may modify the physiological effects, such as the anticancer properties, of foods or diets. Identifying potential interactions among food mixtures or food extract mixtures, which more realistically reflect the pattern of consumption in the human diet, is an important step in determining the utility of plant-based diets in cancer treatment or prevention. Further, conventional cancer treatments, such as chemotherapy and radiation, lack the specificity needed to kill cancer cells without simultaneously damaging normal cells, which is evidenced by serious side effects (e.g., nephrotoxicity, hepatotoxicity, ototoxicity, and neurotoxicity) that accompany these treatments. 24,31 Using paired cell lines allowed us to determine whether selected food combinations limit proliferation of cancerous cells with less or no effect on normal cells.
Two paired cell lines are selected in this study to evaluate whether the mixture(s) of raspberry and sumac would have the reasonable cytotoxic effects on cancer cells, as well as chemoprotective effects on normal cells, and compared with that when applied with sumac or raspberry alone. The first paired cell lines include normal rat colonic epithelial cells (4D/WT) and transformed colonic epithelial cells (D/V-Src), which have been used to determine dietary docosahexaenoic acid (DHA) selectively the nucleoside analogue arabinosylcytosine toxicity toward rat colon tumor cells D/V-Src, while protecting the normal cells 4D/WT. 32 While the second pair includes human spontaneously immortalized breast epithelial cells (MCF-10A) and breast cancer cells (MCF-7), which have been used to evaluate selective cytotoxicity of selected specific polyphenol fractions from red grape wine toward MCF-7 breast cancer cells, as compared with the nontumorigenic MCF-10A cells. 18
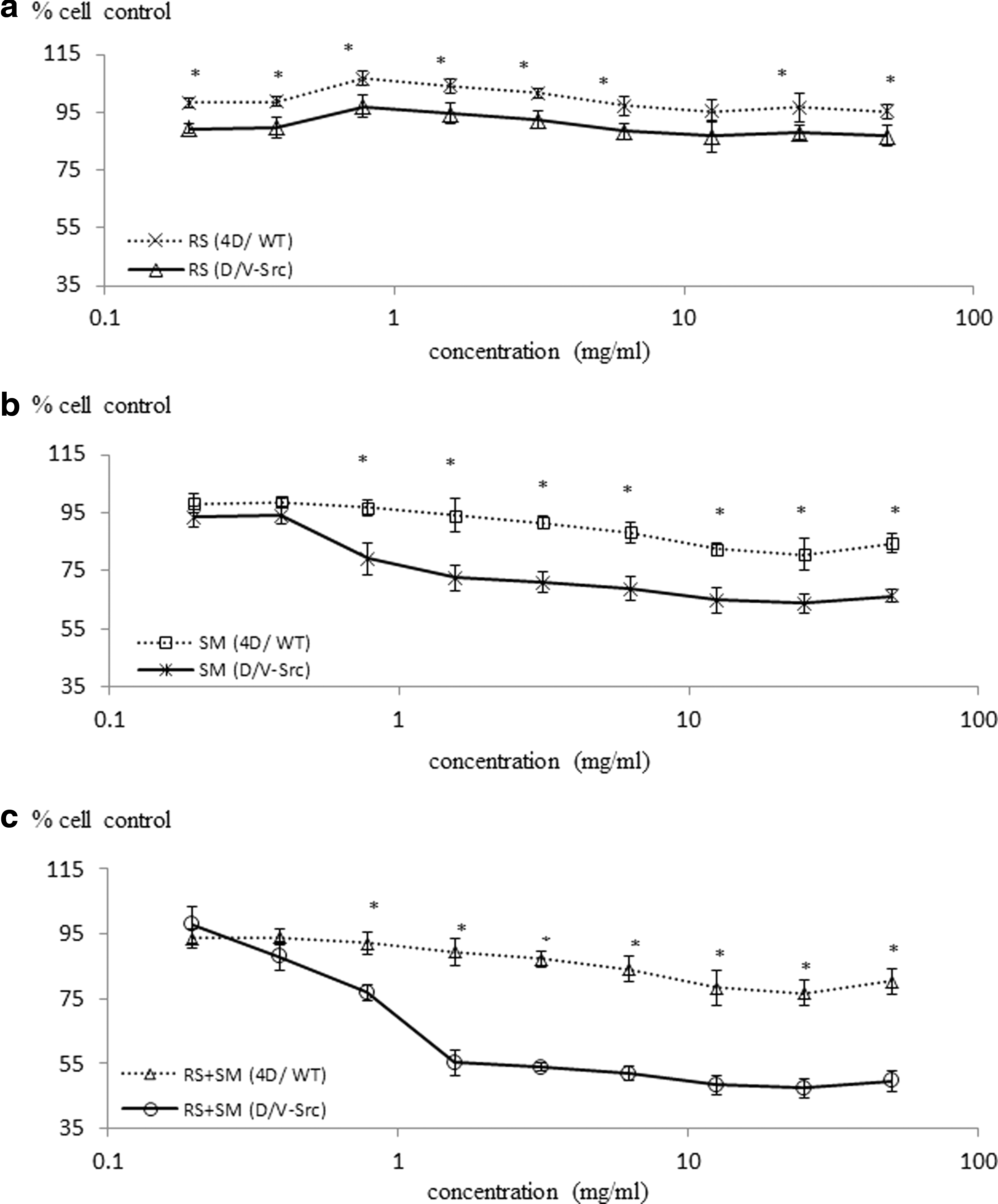
Figure 5 showed the viability of MCF-7, MCF-10A, 4D/WT, and D/V-Src cells that was studied when they were exposed to sumac, raspberry, or their mixtures. The number of cells in the control wells, grown in the absence of extracts, was taken as 100% (SRB staining intensity). Both sumac and the mixture inhibited cell proliferation in a dose-dependent manner (Fig. 5). Among sumac, raspberry, and their mixture, the mixture was the most inhibitory to cell proliferation, followed by sumac, while inhibition in the presence of raspberry was relatively minor for all four cell lines.
The inhibition of cancer cells by other raspberry varieties has been previously reported. Liu et al. observed that four varieties of raspberries (Heritage, Kiwigold, Goldie, and Anne) possessed inhibitory effects on HepG2 cell proliferation, 15 while others reported that raspberries were capable of inhibiting the growth of MCF-7, with IC50=190.80±0.05 μg/mL. 17 Here, we found that the inhibition of raspberries was relatively minor to MCF-7 and D/V-Src cancer cell proliferation (Figs. 4 and 5). The difference may be attributed to varying phytochemicals in raspberries of different genotypes. 15,17

The inhibitory effect of
The inhibitory effect of
Sumac extract was more antiproliferative toward both rat colon and human breast cancer cells as compared with both normal rat colon and human breast epithelial cells at some doses (P<.05) (Figs. 4 and 5). However, the relative effectiveness of the sumac on the four cell lines was also different. Of the four cell lines studied, MCF-7 was the most susceptible to inhibition, followed by D/V-Src cells, MCF-10A, and 4D/WT. These findings, in turn, have generated opportunities for the development of therapy potential of sumac toward specific type of cancer.
With increasing interest in nontoxic natural and therapeutic approaches in the cancer treatment, major emphasis has been placed on employing specific dietary component or foods to target cancer cells with minimal toxic effects to normal cells. Polyphenolics had such inhibition effects on human breast cancer cells, while having less effect on normal cells. 18,33 Up to date, little attempt has been made to evaluate the impact of complex food combinations on cell survival and cell death of normal and cancerous cells. We found that the combination of sumac and raspberry resulted in a synergistic antiproliferative effect against MCF-7 and D/V-Src cells compared with that of sumac or raspberry treatment alone, with the concentration of 1.5 and 3.1 mg FW/mL, respectively (Figs. 4 and 5). An interesting aspect of this mixture effect was that the 4D/WT cell lines did not have any cytotoxic effects even at concentrations where the growth of D/V-Src cells was inhibited by over 80%. It has been reported that only a small proportion (<4 mg) was excreted in the urine 5 h after consumption of 3.57 g of total anthocyanins in 150 mL of elderberry juice. The limitation of this study is that only in vitro models, including cells in culture, were used in identifying the therapeutic potentials of testing sample. In vivo evaluations are probably the most important one in the future for understanding the regulatory roles of digestion, absorption, and metabolism of sumac and raspberry mixture in determining tissue concentrations of potentially active compounds. Only with this information will we be able to translate the experimental findings into tools or dietary recommendations for human populations and for industrial development of desirable sumac-based functional or nutraceutical products. The mechanism of synergistically enhanced ACs and selectivity observed in the inhibition of cancerous cell proliferation by the sumac and mixtures of raspberries and sumac need to be explored. Polyphenolic compositional differences (e.g., anthocyanins) between sumac and raspberry 13,14 are most likely to result in multisite targeting and synergistic effects. It should be noted that the synergistic response in the mixtures may also possibly arise from phytochemicals other than polyphenolics interacting with each other.
Summary
We found that the mixture of raspberry and sumac extracts expressed synergistic responses in TPC, FRAP, ORAC, and DPPH assays. It should be noted that nonphenolic phytochemicals could contribute to the antioxidant and biological activities of single foods and their mixtures. Additional study will be required to identify other potential bioactive compounds. The mixture containing phytochemicals could possibly act through more than one antioxidant mechanism. This combination was considered one of the strongest candidates for potential antioxidant ingredients in the functional food development. Meanwhile, sumac extracts appeared toxic to RAT-2 and C2C12 cells and inhibited both cells in a dose-dependent manner. This could result in challenges in developing sumac-derived antioxidant supplementation for food application. Encouragingly, the mixture of sumac and raspberry was found to be a strong inhibitor of proliferation of cancer cells (MCF-7 and D/V-Src), while having less effect on normal cells (MCF-7 and 4D-WT). It is most likely that diverse phytochemicals in the sumac/raspberry mixture could exhibit multisite targeting and synergistic detrimental influence on cancer cells, whereas they are marginally cytostatic toward normal cells. Promisingly, synergism derived from the mixtures of sumac and raspberries could be functional foods with enhanced AC and/or anticancer potentials.
Footnotes
Author Disclosure Statement
No competing financial interests exist.