
Abstract
Liver fibrosis, a common condition occurring during the evolution of almost all chronic liver diseases, is the consequence of hepatocyte injury that leads to the activation of Kupffer cells and hepatic stellate cells (HSC). Silymarin (Si) is a herbal product widely used for its hepatoprotective potential. Our study aims to investigate the effects of two different doses of Silymarin on a CCl4-induced model of liver fibrosis with a focus on the early stages of liver injury. Fifty Wistar rats were randomly divided into five groups (n=10): control group (sunflower oil twice a week); CMC group (carboxymethyl cellulose five times a week, sunflower oil twice a week); CCl4 group (CCl4 in sunflower oil, by gavage, twice a week); CCl4+Si 50 group (CCl4 twice a week, Silymarin 50 mg/b.w. in CMC five times a week); and CCl4+Si 200 group (similar to the previous group, with Si 200 mg/b.w.). One month after the experiment began we explored hepato-cytolysis (aminotransferases and lactate dehydrogenase), oxidative stress, fibrosis (histological score, hyaluronic acid), markers of HSC activation (transforming growth factor β1 [TGF-β1], and α-smooth muscle actin [α-SMA] expression by western blot) and activation of Kupffer cells by immunohistochemistry. Our data showed that Si 50 mg/b.w. had the capacity of reducing oxidative stress, hepato-cytolysis, fibrosis, activation of Kupffer cells, and the expression of α-SMA and TGF-β1 with better results than Si 200 mg/b.w. Thus, the usual therapeutic dose of Silymarin, administered in the early stages of fibrotic changes is capable of inhibiting the fibrogenetic mechanism and the progression of initial liver fibrosis.
Introduction
L
Silymarin (Si) is a herbal product widely used in liver diseases all over the world for its hepatoprotective potential. In fact, it was its capacity of stabilizing cell membranes which first recommended Silymarin for medical use. In the last decade, other beneficial roles of Silymarin administration were emphasized from experimental research and clinical trials: antioxidant, anti-inflammatory, immunomodulatory, and lately even antifibrotic and liver regenerating capacity. 4
In terms of clinical efficacy, the existing results are not definitive. 5 However, a few conclusions are constantly confirmed by the performed experimental studies: the antifibrotic effect of Silymarin on well-established fibrosis is evident, 6 –8 while its capacity of reversing well-established cirrhosis is irrelevant. 9 There is no efficiency on the mortality rate of patients with cirrhosis. 10 Concurrently, reports of adverse effects while receiving Silymarin are rare. 10
But what about the early stages of fibrosis?
Existing studies focus on the effects of Silymarin in advanced stages of liver fibrosis. However, in medical practice, Silymarin is recommended whenever the transaminase level is high. Some of these situations could occur in chronic liver disease, when the persistence of hepatocytes aggression and the association of chronic inflammation can lead, in time, to the onset of liver fibrosis. It would be important to know whether Silymarin administration in such pathologies, with the main objective to reduce hepatocytolisis, did not have, in time, a beneficial effect on the initiation and progression of liver fibrosis as well, even if, as first intention, this is not the desired effect. Theoretically, the stabilization of hepatocyte membranes with the decrease of hepatocytolisis and a decline in the release of activators for HSC stimulation should have a beneficial effect on the subsequent evolution of liver fibrosis. To our knowledge, although there are numerous studies regarding the effects of Silymarin on liver fibrosis, the echo of commonly used Silymarin therapeutic doses in chronic hepatopathies accompanied by the increase of transaminases on the initiation and subsequent progression of liver fibrosis has not yet been sufficiently studied.
It was proven, 11 especially by in vitro studies, that the effects of Silymarin are dose-dependent. If Silymarin could interfere with the initial stages of fibrosis, could this effect be achieved by the usual doses of prescribed Silymarin, or only by higher doses?
The aim of our study is to evaluate, on a CCl4-induced model of experimental liver fibrosis, the impact of Silymarin, in usual doses, on the initiation and progression of fibrosis during its early stages and to establish whether the effects are more important for a higher dose of herbal product than for the usual, therapeutic, dose.
Materials and Methods
Experimental design
Fifty male Wistar rats (weight: 200±10 g) have been used throughout the experiment. Thirty of them formed the test groups and received CCl4 solution 1.2 mL/kg b.w by gavage, twice a week (25% in sunflower oil). 12 After 2 weeks, they were randomly divided in three groups (n=10): one group (CCl4) continued to receive the CCl4 solution twice a week, the other two groups received not only CCl4, according to the previous protocol of administration, but also Silymarin extract suspended in 0.7 mL carboxymethyl cellulose (CMC) solution 0.5% as vehicle. Silymarin extract from Silybum marianum Gaertner was provided by Woerwag Pharma and the HPLC analysis (data not shown) described a 44.7% Silymarin content in the provided powder expressed as silibinin.
The Silymarin was administered by gavage, five times a week, in a dose of 50 mg Silymarin/kg b.w. (group CCl4+Si 50) or in a dose of 200 mg Silymarin/kg b.w. (group CCl4+Si 200). Two control groups were used in the experiment: the control group (ctrl) received 1 mL/kg b.w. sunflower oil twice a week, by oro-gastric administration, and the CMC group received only sunflower oil as the ctrl group for 2 weeks, but, from the third week on, 0.7 mL CMC 0.5% solution was added five times a week to the oil administration. After 4 weeks, all the animals were sacrificed. Blood and liver were collected.
The animals were housed in the animal facility of the Physiology Department from the “Iuliu Haţieganu” University of Medicine Cluj-Napoca, at 24°C temperature, under a 12:12 light/dark cycle and with free access to water and food. The experimental protocol has been approved by the Ethics Committee of the University of Medicine and Pharmacy Cluj-Napoca.
Preparation of biological samples
Blood was centrifuged at 1000 g for 5 min to separate the plasma. The left lateral lobe and right medial lobe of the liver were immersed in 10% formalin solution and prepared for histological analysis, and the rest was washed with cold saline and homogenized with a Polytron homogenizer (Brinkman Kinematica, Lucerne Switzerland) in 50 mM TRIS–10 mM EDTA buffer (pH 7.4). The homogenates were centrifuged at 2500 g to obtain the supernates. From each animal, plasma and supernates were stored in aliquots at −80°C until assayed.
Cell membrane injury assessment
Measurements were performed as described in our earlier publications. 13 The activity of aspartate aminotransferase (ASAT) and alanine aminotransferase (ALAT) were assessed by semiautomatic analysis, using colorimetric kits, according to the manufacturer's instructions. 14
Lactate dehydrogenase (LDH) activity was evaluated with a spectrophotometric method 15 with some adjustments. The amount required to catalyze the reduction of 1 μmol nicotinamide adenine dinucleotide (NAD)/min mg protein was considered one unit of activity.
Measurement of oxidative stress parameters
The oxidative stress parameters were assessed following the same protocols as in our earlier publications. 16 The Bradford method was used to measure the protein concentration in liver homogenates. 17
Briefly, to determine the malondialdehyde (MDA) level in liver homogenates, we used a fluorimetric method with 2-thiobarbituric acid. 18 MDA was determined in the organic phase using a synchronous technique with excitation at 534 nm and emission at 548 nm. For protein carbonyls (PC), a fluorimetric method with 2,4-dinitrophenyl-hydrazine was used. 19 The readings were performed with a spectrophotometer at 355–390 nm.
Reduced glutathione (GSH) and oxidized glutathione (GSSG) were fluorimetrically measured using o-phtalaldehyde. The fluorescence was recorded for 350 nm excitation and 420 nm emission. The concentrations for GSH and GSSG were determined using standard curves. 20
Fibrosis assessment
Histologic evaluation and hyaluronic acid assay were performed. For the histological exam, hepatic samples were fixed in 10% buffered neutral formalin and embedded in paraffin. Sections were made at 4 μm and the slides were stained using Hematoxilin–Eosine and Masson's trichrome methods. Sections were examined by an independent observer blinded to the experimental protocol. The score of fibrosis was semiquantitatively assessed, using a histological grading system described by Matsuzawa et al. 21 with minor modifications. According to this description, 0 was defined as no fibrosis, 1 was defined as perivenular fibrosis, 2 was defined as focal bridging fibrosis, 3 was defined as bridging fibrosis with lobular distortion without cirrhosis, and 4 was defined as cirrhosis.
Hyaluronic acid is an essential component of extracellular matrix (ECM), synthesized in the liver by HSC. Hyaluronic acid assay was performed using an Hyaluronan Duoset DY 3614 ELISA kit from R&D Systems (Minneapolis, MN, USA), according to the manufacturer's instructions.
Activation of HSC evaluation
To evaluate the activation of HSC, α-smooth muscle actin (α-SMA) and TGF-β1 expression were assessed by western blot. The liver samples were homogenized as we previously described. 12 Total proteins (20 μg/lane) were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (12% polyacrylamide) at 200 mV, blotted on polyvinylidenedifluoride membranes for 60 min at 100 mV using Biorad Miniprotean system (BioRad-Hercules, USA), and incubated with the primary antibodies for α-SMA, TGF-β1 (both provided by Abcam plc, Cambridge, United Kingdom), or GAPDH (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The membranes were then incubated with corresponding secondary peroxidase-coupled antibody (Santa Cruz Biotechnology), with GAPDH serving as endogenous control. The detection of proteins was performed with Supersignal West Femto Chemiluminiscent substrate (Thermo Fisher Scientific, Rockford, IL, USA). The membranes were exposed to an X-ray film (Kodak, Rochester, New York, USA) for ∼2 min and the films were developed and analyzed using Phoretix Array.
Evaluation of Kupffer cells activation
The immunohistochemical study, using antibodies against CD68 (dilution 1/150), provided by Linaris Biologische Produkte (Dossenheim, Germany), was performed with a Leica Bond-max™ Immunohistochemistry system to identify activated Kupffer cells. CD68-positive cells were quantified on 10 fields of×40 sections using an Olympus BX51 light microscope.
Statistical analysis
The results were expressed as mean±standard deviation. Data were analyzed with GraphPad Prism 5.0 software (GraphPad, San Diego, CA, USA), using one-way analysis of variance (ANOVA) followed by the Tukey's multiple comparisons post-test. A P<.05 was considered statistically significant.
Results
All animals survived the experiment. The livers of the animals in the ctrl and CMC groups showed no macroscopic changes. Grossly, the liver of experimental animals had a finely nodular surface, and pale color.
Protective effect of Silymarin on hepatocyte membrane damage
The levels of ALAT (Fig. 1A) and ASAT (Fig. 1B) were significantly increased after 4 weeks of CCl4 administration (63.75±2.63 U/L, respectively 119.30±17.40 U/L) compared with controls (14.92±2.25 U/L and 37.75±1.71 U/L, respectively). Both doses of Silymarin significantly decreased the levels of ALAT and ASAT compared with the CCl4 group. We noticed that for ALAT there was no significant difference between the effects of Silymarin at both doses, while for ASAT the higher dose (Si 200 mg/b.w.) had a more potent effect on the decrease of this parameter. The value for ASAT in the group that received Si 200 mg/b.w was not significantly different compared to both control groups (ctrl and CMC), but was significantly decreased when compared with the group that received Silymarin 50 mg/b.w.
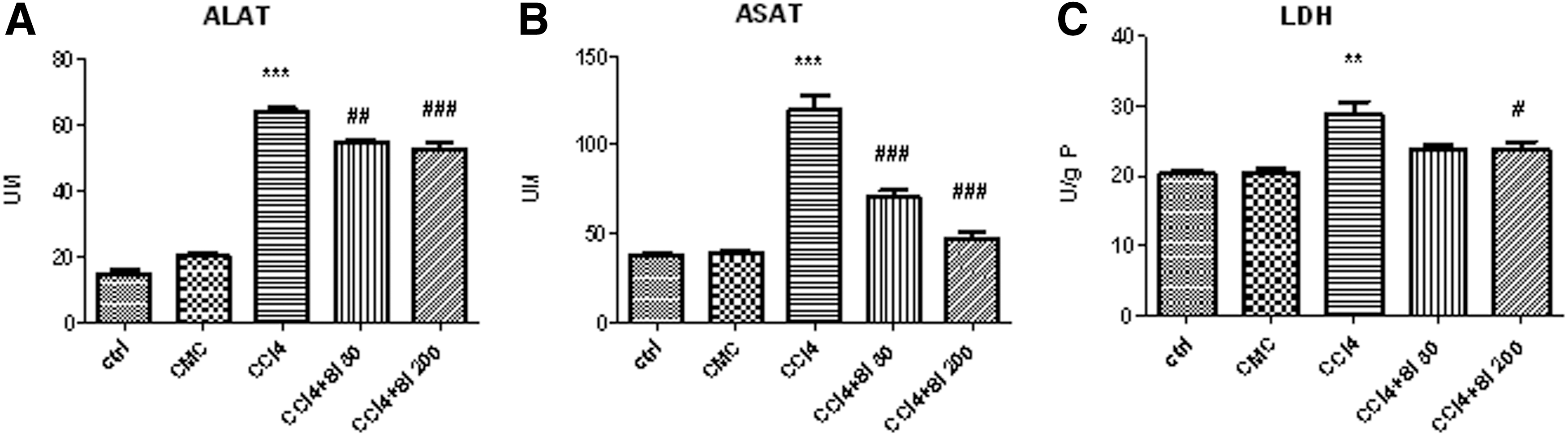
Effects of Silymarin administration, 50 mg/b.w or 200 mg/b.w., on alanine aminotransferase (ALAT)
For LDH level (Fig. 1C) only the higher dose of Silymarin, 200 mg/b.w, significantly decreased the level of LDH compared with the CCl4 group (23.68±3.03 and 28.78±5.32 U/g protein [P], respectively), but there was no statistical difference between both doses of Silymarin. At the same time, there were no significant differences for LDH levels between the values obtained in control group (ctrl), CMC group, CCl4+Si 50 mg/b.w. group, and CCl4+Si 200 mg/b.w. group.
Silymarin reduces oxidative stress in liver
Lipid peroxidation, quantified through MDA level in liver homogenates (Fig. 2A), was significantly increased after CCl4 administration (0.22±0.012 nmol/mg protein [P]) compared with control group (ctrl), and also with CMC group (CMC) (0.096±0.012 and 0.097±0.013 nmol/mgP, respectively). Si 50 mg/b.w. significantly decreased the MDA level (0.08±0.01 nmol/mgP) compared with CCl4 group, to values similar to those of the control (ctrl) and vehicle (CMC) groups. Administration of Si 200 mg/b.w also significantly decreased the level of MDA (0.14±0.02 nmol/mgP) compared with the CCl4 group, but the mean value remained significantly increased compared with the ctrl group, CMC group, and CCl4+Si 50 group.
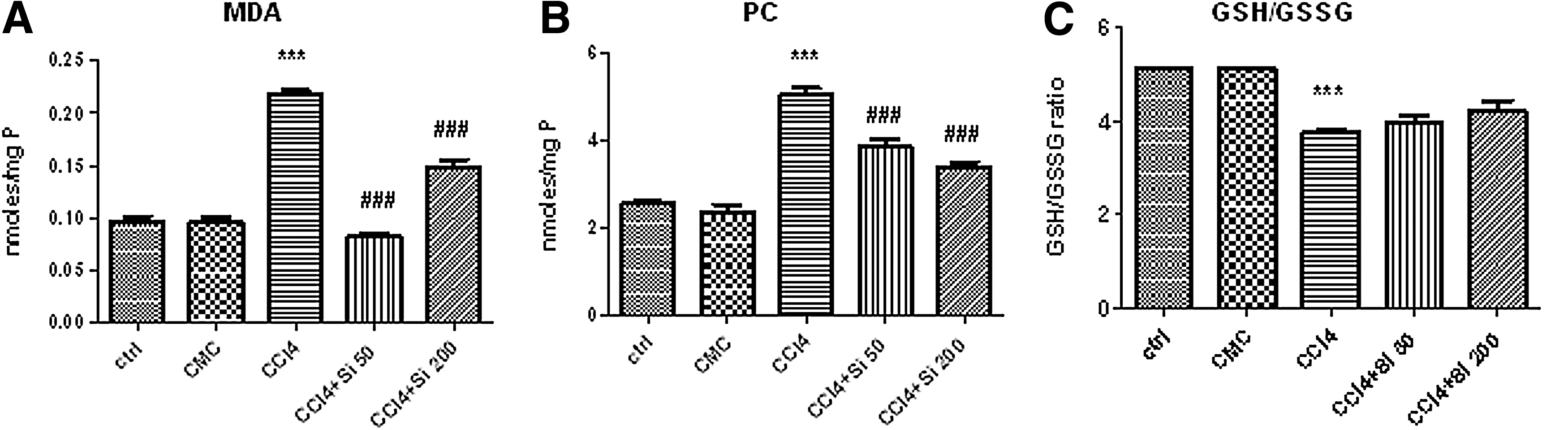
Effects of Silymarin administration upon the oxidative stress parameters in liver homogenates, using an experimental model of CCl4-induced hepatic fibrosis. CCl4 group received CCl4 by gavage twice a week for 1 month and the groups with Silymarin administration received only CCl4 for the first 2 weeks, but from the third week on Silymarin 50 mg/b.w. was added five times a week (CCl4+Si 50 group), or, respectively, Silymarin 200 mg/b.w (CCl4+Si 200 group) until the end of 1 month of experiment. For all the assessed parameters, ***P<.001 was used to indicate the statistical difference between the CCl4 group and the ctrl and CMC groups.
Both doses of Silymarin, 50 mg/b/w. and 200 mg/b.w. have similar effects on PC generation (Fig. 2B), with a significant decrease of their level compared with the CCl4 group (3.87±0.45 nmol/mgP, 3.42±0.26 nmol/mgP, and 5.06±0.41 nmol/mgP respectively). There was no significant difference between the groups treated with Silymarin in either dose, but the PC level in the Si-treated groups remained statistically increased compared with the ctrl group, or to the CMC group.
The antioxidant capacity was quantified using the ratio between reduced GSH and GSSG in the liver (Fig. 2C). The CCl4 administration significantly decreased the GSH/GSSG ratio (3.76±0.12) compared with the ctrl group (5.14±0.21) and to the CMC group (5.15±0.02). The administration of Si 50 mg/b.w. or 200 mg/b.w. increased this ratio (3.99±0.24 and 4.23±0.47, respectively) but with no statistical significance when compared to CCl4 group. The ratio remained significantly lower when compared with the ctrl and CMC, and between the two doses there was no significant statistical difference.
Silymarin reduces liver fibrosis
The fibrosis was assessed through the histological fibrosis score and the hyaluronic acid evaluation using ELISA assay from liver homogenates. Liver sections from animals in the control group showed normal liver architecture. The CCl4 administration determined vacuolar dystrophy, oval cell hyperplasia, and necrosis of the hepatocytes from the central areas (Fig. 3A). Central fibrosis and centro-central bridging fibrosis were also common features. The centro-central fibrous bridges lead to the forming of liver nodules.
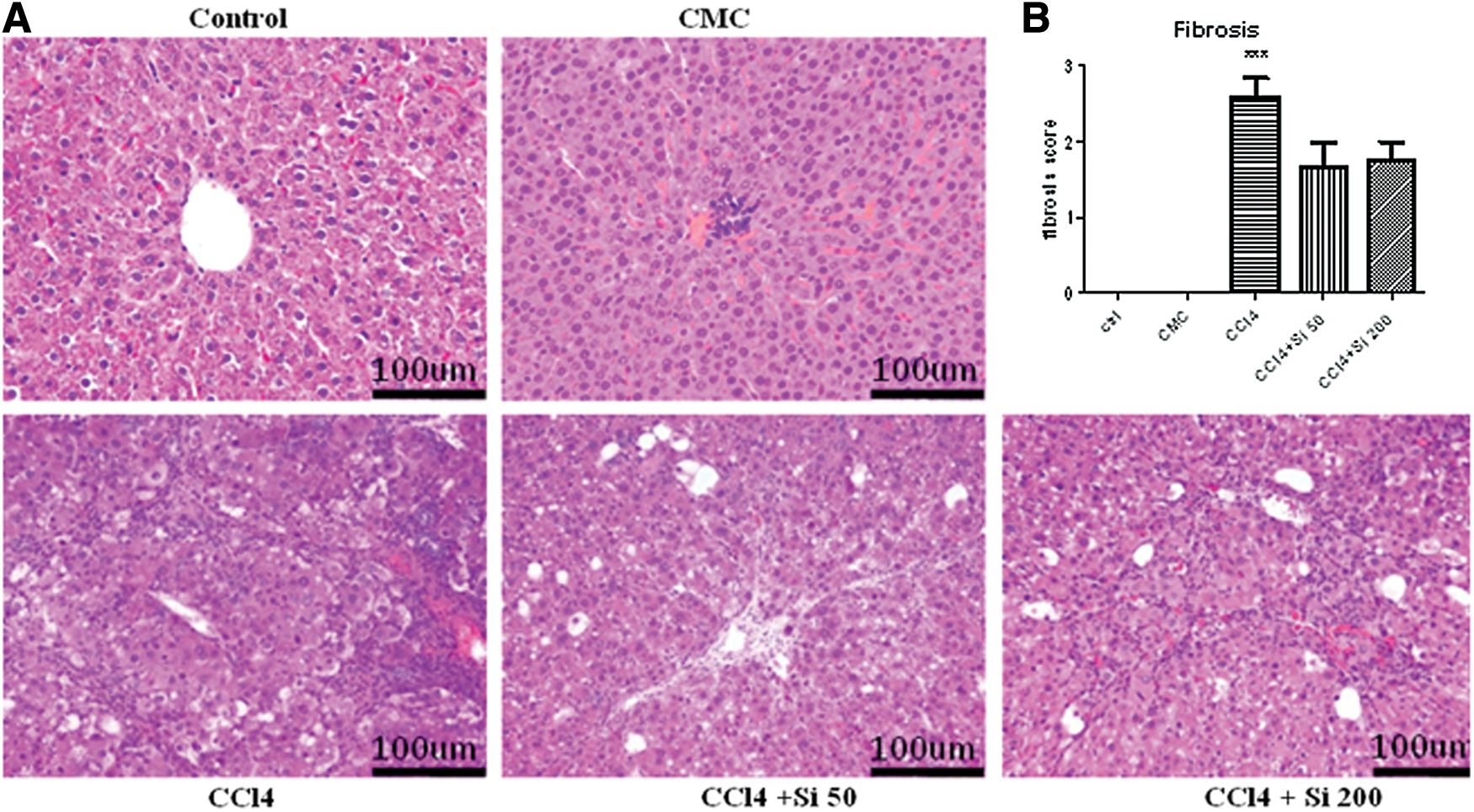
Effects of Silymarin administration upon the histological changes produced by the CCl4 administration.
The severity of the lesions varied between different experimental groups, Silymarin significantly reducing the fibrosis score.
The fibrosis score for ctrl and CMC groups was 0, while for the CCl4 group it was 2.6±0.5. The Silymarin administration significantly decreased (P<.001) the fibrosis score for both doses compared with the CCl4 group (1.66±0.57 and 1.75±0.52, respectively), with no significance between them (Fig. 3B).
The CCl4 administration alone significantly increased the level of hyaluronic acid in the liver (Fig. 4) compared with the ctrl and CMC groups (6.95±1.15 pmol/mgP vs. 3.21±0.32 pmol/mgP and 3.08±0.52 pmol/mgP, respectively). Both doses of Silymarin significantly decreased the hyaluronic acid level compared with the CCl4 group (4.02±0.73 pmol/mgP; 4.32±0.65 pmol/mgP), without a significant difference between the two results. The administration of Silymarin, in either dose, reduced the levels of hyaluronic content to values similar to the ctrl and CMC groups.

Effects of Silymarin administration (50 mg/b.w. or 200 mg/b.w.) upon the hyaluronic acid content in liver homogenates, using an experimental model of CCl4-induced hepatic fibrosis. The CCl4 administration alone significantly increased the level of hyaluronic acid in the liver (***P<.001) compared with ctrl and CMC groups. Both doses of Silymarin significantly decreased the hyaluronic acid level compared with CCl4 group (### P<.001), without a significant difference between the two results.
Silymarin reduces HSC activation
The quantification of α-SMA and TGF-β1 was performed by using western blot.
By using western blot assay, increased levels of α-SMA (Fig. 5A, B) were detected in the liver lysates of rats in the CCl4 group compared with both ctrl and CMC groups (P<.001). The administration of Silymarin 50 mg/b.w. significantly decreased α-SMA (P<.001), while Silymarin 200 mg/b.w. did not show any statistical decrease compared with the CCl4 group.
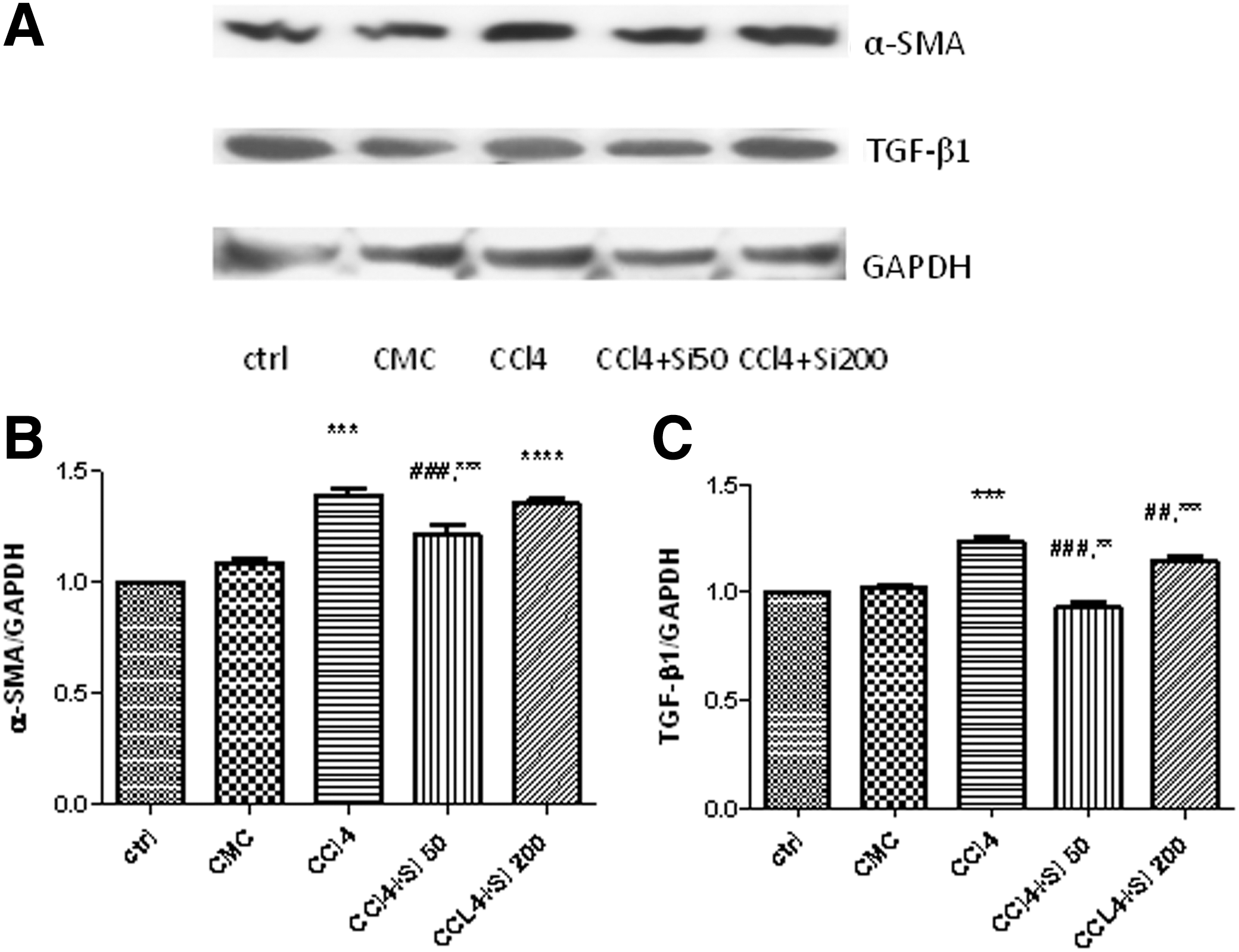
TGF-β1 expression (Fig. 5A, C) was significantly increased (P<.001) in the CCl4 group compared with the ctrl and CMC groups. Silymarin in either dose significantly decreased the TGF-β1 expression, but the effect was more important (P<.001) for the Silymarin 50 mg/b.w. administration, with significant decrease (P<.01) compared with the CCl4 group.
Silymarin reduces Kupffer cells activation
Kupffer cell identification was performed by CD68 immunohistochemistry (Fig. 6). Regarding the CMC group, the administration of carboxymetylcellulose induced the activation and mobilization of CD68-positive Kupffer cells (Fig. 6B). Carbon tetrachloride administration further increased the number of activated Kupffer cells, while the administration of Silymarin in either dose significantly decreased (P<.001) the number of activated Kupffer cell compared with the CCl4 group, and without statistical significance between the two doses (Fig. 6E).
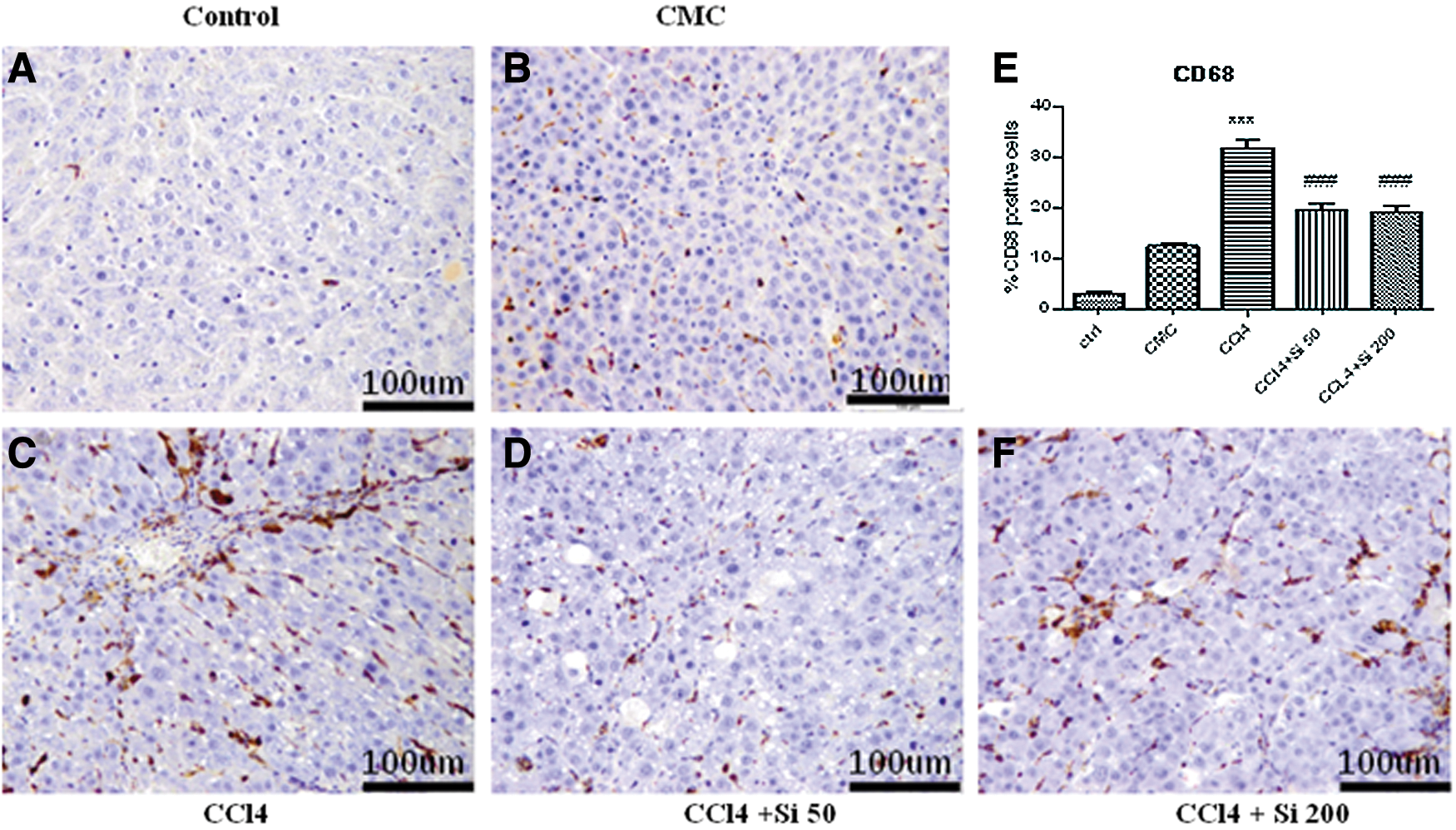
Identification of activated Kuppfer cells by CD68 immunohistochemistry; original magnification of 200×; scale bar=100 μm.
Discussion
The pathogenesis and evolution of chronic hepatic disease vary according to multiple factors (etiologic agent, other associated or concomitant hepatotoxic effects, comorbidities, etc.), but from the morpho-pathological point of view, the unfavorable common pathway is the evolution from the necro-inflammatory dominance, a characteristic of chronic hepatitis, toward progressive hepatic fibrosis and, in the end, the onset of hepatic cirrhosis. 22
The fibrogenic response in the liver, with the excessive production of the ECM proteins, is especially realized by the activation of HSC, which are transformed into myofibroblasts and express α-SMA. 23 ROS are key mediators for HSC activation, both directly and indirectly 24 and also for collagen I gene regulation. 25 Oxidative stress not only anticipates HSC activation, 26 but is both sufficient and indispensable for this activation, as emphasized from in vitro and in vivo studies. 27 Also, activated Kupffer cells, through cytokines release, participate in HSC activation. 3 TGF-β 1 is also one of the most powerful stimulators of HSC. 3
The experimental model of CCl4-induced fibrosis is an optimal in vivo model of oxidative stress-related fibrogenesis, and also a reference model for the study of chronic liver diseases. 26,28 In our previous studies using this model, 12 early central fibrosis was obtained from the third week of CCl4 administration, while central fibrosis with centro-central fibrosis bridges was observed after 4 weeks. So, in this experiment, we realized the same CCl4-induced model of fibrosis, expecting to obtain the first fibrotic changes between the second and the third week of hepatic toxic administration.
Silymarin was added in some groups 2 weeks after the beginning of the experiment in parallel with the CCl4 administration, in two doses: 50 mg/b.w. and 200 mg/b.w.
In choosing the doses used, we have taken into account the results obtained with different doses described in the existing studies and we also considered the fact that the in vitro studies show that the effects of Silymarin are dose-dependent. 11 The 50 mg/b.w. dose was chosen because it is known to be the correspondent of the clinically relevant dose of the standard hepatoprotective treatment. 29 In medical practice, this dose corresponds to the usual Silymarin administration in case of increased transaminase levels. The 200 mg/b.w. dose is the highest dose used so far in experimental studies to prove its indubitable ability to increase the resolution of hepatic changes in well-established fibrosis (8 weeks of CCl4 administration). 6 For this reason, we considered that, by comparing these two doses, we could get a conclusive answer to the question whether the usual therapeutic dose used in clinical practice would be enough to influence the progression of fibrosis in early stages, or higher doses are needed, doses which were proven to have a beneficial effect in other stages of evolution of liver fibrosis.
We concomitantly evaluated the effects of Silymarin on different mechanisms involving the fibrogenic response. Silymarin proved to have a very potent antioxidant effect, with a significant decrease of MDA and PC in the liver as compared to the CCl4 administration alone. For the GSH/GSSG ratio, Silymarin had the tendency to restore the normal values, but with no statistical significance. When comparing the two doses of Silymarin, the highest dose did not prove a more potent antioxidative effect. On the contrary, the MDA decrease was better for the usual dose of this herbal compound administration.
Regarding the effect on hepatic cytolysis, both doses of Silymarin decreased ALAT and LDH similarly, the only difference being the most important effect of Silymarin 200 mg/b.w. on the ASAT level, which was restored to almost normal values.
The histological evaluation and immunohistochemical assessment of activated Kupffer cells revealed the same capacity of both doses of Silymarin of decreasing the fibrosis score and the number of activated Kupffer cells compared to CCl4 administration, while more sensitive tests emphasized the better antifibrotic capacity of Silymarin 50 mg/b.w. in these early stages, with the decrease of hyaluronic acid content of the liver, and the decreased α-SMA and TGF-β1 expression.
Considering the existing data in the scholarly literature and the important protective effect that the high Silymarin dose has on the hepatocytolysis in our experiment, compared with the usual dose effect, we would expect Si 200 mg/b.w. to have a better antifibrotic effect than 50 mg/b.w. However, paradoxically, the significant decrease of α-SMA and of TGF-β1 expression compared with the simple CCl4 administration is triggered by Si 50 mg/b.w. and not by the high Silymarin dose, although the latter has effects similar to the usual Silymarin dose both on the hyaluronic acid values and on the histological assessment of fibrosis. Equally surprising are the results on the oxidative stress parameters because the high Silymarin dose produces the expected changes in the assessment of the PC and in the GSH/GSSG ratio, but MDA stays significantly high under the Si 200 mg/b.w. action compared with the control, CMC, and CCl4+50 groups. At this stage of our experiments, we cannot fully explain the results. Given the experimental model, fibrosis stage, and dosage, it is likely that pathogenic intracellular pathways are activated, pathways that our current research cannot yet reveal but aim to elucidate in future studies.
In experimental models with CCl4-induced liver injury, the evaluation of Silymarin effects after 6–8 weeks of concomitant administration of the two substances revealed the capacity of Silymarin of reducing the liver MDA content, transaminases, and LDH, 30 of increasing GSH, 10,30 of reducing hidroxyproline content of the liver, 31 of reducing α-SMA and TGF-β1 expression, 8 and of decreasing the fibrosis score. 7
Kupffer cells, which are the resident macrophages of the liver, have, in the last years, lost their position as principal cells involved in the progression of liver fibrosis, while HSC have become the leading player in the ECM accumulation. 32 However, the role of Kupffer cells in the activation of HSC should not be overlooked. Hepatocyte injury is the first step that promotes fibrosis. Oxidative stress triggers the mitochondrial pathway of apoptosis and the phagocytosis of apoptotic bodies by liver macrophages activates HSC, leading to fibrosis. 1 At the same time, the influx of Kupffer cells in the injured areas is concomitant with the appearance of HSC activation markers. 32 It has been demonstrated that not only in patients, but also in cultured hepatocytes, there is an in situ correlation between the presence of MDA and the collagen gene expression by HSC 33 and the collagen synthesis by cultured HSC under the MDA stimulation. 34 Also, the injured hepatocytes are the source of these fibrogenic lipid peroxides. However, the fibrogenetic mechanisms are much more complex, as even in contact with isolates HSC, Silymarin is capable of suppressing their activation. 35
Taking into account all these considerations, we can conclude that the administration of Silymarin in the early phases of liver injury is capable of interfering with the mechanisms leading to further progression of liver fibrosis by stabilizing the cell membrane. Our study clearly indicates that Silymarin administration reduces not only the levels of transaminases, but also the oxidative stress in liver and the activation of HSC, with the reduction of fibrosis score and hyaluronic liver content, in the early stages of fibrosis onset. This effect can be obtained for the usual therapeutic dose of Silymarin. Also, the fact that in patients with HCV and normal levels of transaminases, the progression of fibrosis is stopped is a confirmation of the validity of our hypothesis. 36
In conclusion, the usual dose of Silymarin administered in the early stages of fibrotic changes is capable of inhibiting the fibrogenetic mechanism and the progression of liver fibrosis, even if the antifibrotic effect is not the targeted one.
Footnotes
Acknowledgments
We are grateful to Dr. Simion Baltes for the Silymarin extract. We thank Mr. Remus Moldovan for animal handling and Ms. Nicoleta Decea for the assessment of biochemical parameters. We also express our gratitude to Mr. Alupei Marius and Dr. Manuela Banciu for their contribution to western blot assessment.
Author Disclosure Statement
No competing financial interests exist.