
Abstract
A water soluble extract from the medicinal mushroom Agrocybe aegerita has been shown to stimulate splenocyte proliferation, cytotoxic activity, and tumor rejection effect in tumor-bearing mouse models. In the present study, the crude extract was separated into a protein component fraction (Yp), mainly containing lectins and serine proteinase, and a small molecule component fraction (Ys), mainly containing triethylene glycol, α-bisabolol, n-hexadecanoic acid, and so on. The antitumor activity of the fractions was investigated in a tumor-bearing BALB/c mouse model. Repeat administration of Yp and Ys significantly inhibited tumor growth (P<.001), but little toxicity was observed. Moreover, the protein fraction Yp performed better than Ys in both antitumor and lifespan-prolonging activity. The cytokine expression levels in serum and splenocytes from extract-treated mice were selectively screened by enzyme-linked immunosorbent assay and quantitative real-time polymerase chain reaction, and the results showed that Yp upregulated the mRNA level of Th2 cytokine interleukin-10 (P<.01), and Ys increased the mRNA level of granulocyte-macrophage colony-stimulating factor (P<.01) and anti-inflammatory cytokine transforming growth factor-β (P<.01). All these data suggest that Yp and Ys can inhibit tumor growth via different mechanisms, which promotes the understanding of antitumor properties of medicinal fungi.
Introduction
T
The bioactive compounds from medicinal mushrooms have been reported including mainly polysaccharides, glycol-protein, protein/peptide, selenium, lipids, and phenols; 3,4 β-glucan is a well-known biologic response modifier (BRM), which is the major molecular structure of lentinan from Lentinus edodes, schizophyllan from Schizophyllum commune, and grifolan from Grifola frondosa. The polysaccharides contribute to the activation of immune cells, such as hematopoietic stem cells, lymphocytes, macrophages, dendritic cells, and natural killer cells, resulting in the production of cytokines and antitumor activity. 5 However, the research into the antitumor mechanism of other active constituents, especially in vivo data, remains limited. 6
Cytokines are important regulators of host defense, inflammation, and immune response. The cytokine networks are remarkable signals when metabolic homeostasis is altered. 7 –9 There are Th1 and Th2 cells as balancers in humoral and adaptive immunity, which secrete Th1 cytokines interleukin (IL)-2, IL-12, interferon (IFN)-γ, and tumor necrosis factor (TNF)-α/β, and Th2 cytokine IL-4 and IL-10 respectively. 10 The dynamic crosstalk of cytokines collectively induces either pro- or anti-carcinogenic signals. In the present study, the cytokines IL-2, IL-4, IL-10, TNF-α, IFN-γ, anti-inflammatory cytokine transforming growth factor (TGF)-β, and granulocyte-macrophage colony-stimulating factor (GM-CSF) were selected to describe the bioactivity of the extracts in tumor-bearing mouse models.
Previously, a water-soluble component with trace polysaccharides from the medicinal mushroom Agrocybe aegerita (Yt) showed high antitumor activity against several tumor cell lines, and tumor rejection activity in H22 and S180-bearing mouse models, and the immunostimulating effect revealed proliferation of splenocytes, enhancement of cytotoxic activity, and alteration of cytokine production. 11 In the present study, Yt was separated into a protein component fraction (Yp) and a small molecule component fraction (Ys) for a bioactivity study. The antitumor activity analysis of the two components in mouse models will help with the assessment of the medicinal value of mushrooms and the discovery of new potential drug candidates.
Materials and Methods
Sample preparation
The medicinal fungi A. aegerita were collected from the Sanming Institute of Fungi. The fungi were grown on compost under typical cultivation parameters at the Mushroom Research Center of Huazhong Agriculture University. The fresh fruit body was dried at 60°C and crushed into a powder. Total component (Yt) was extracted as previously described, 11 separated by chromatography over a column (10 cm×1.5 cm) of ion-exchanger Diaion HP-20, and eluted by a stepwise gradient of aqueous ethanol (75–85%) at flow rate 1 mL/min. The flow-through fraction was named as Yp (the protein component fraction), and the elution fraction was Ys (small molecule component fraction).
Matrix-assisted laser desorption/ionization–tandem mass spectrometry (MALDI-TOF/TOF)
Yp was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) followed by staining with Coomassie blue. Protein bands were excised from the gel, and in-gel tryptic digestion, N-terminal derivatization of the tryptic peptides with 4-sulfophenyl isothiocyanate (SPITC), and the collection and analysis of mass spectrometry (MS) data was performed as previously described. 11
Gas chromatography/mass spectrometry (GC-MS) analysis
The GC-MS analysis was performed by using a gas chromatograph equipped with a 50-m HP-5MS capillary column coupled to an Agilent MSD 5973N quadrupole mass spectrometer with an electron impact (EI) ion source as the detector. The temperature program was 2 min at 150°C, then 5°C/min to 180°C and 15°C/min to 260°C, and holding time was 5 min. UHP helium was used as the carrier gas at flow rate of 1.0 mL/min. The detector was operated at 120 ev in EI mode with full scan (35–450 amu). The ion source temperature was 230°C. The temperature of the quadrupole was 150°C. The software used was D.01.02, with the NIST02 mass database.
Animal treatment
Male and female BALB/c mice (male and female; 6 weeks old) were purchased from the animal center of the epidemic prevention sector in Hubei province and kept under standard conditions (in laminar airflow cabinets under pathogen-free conditions with a 12-h light/12-h dark schedule) and fed with an autoclaved Harlan Teklad sterilizable rodent diet ad libitum.
Tumors were established by subcutaneous injection of 5×105 H22 cells in a volume of 0.1 mL in the right oxter of BALB/c mice. The mice were randomly distributed into control and treatment groups (n=20 mice per group) and intratumorally injected with phosphate-buffered saline (PBS; control group) or with Yp (2.5 mg/kg body weight), Ys (2.5 mg/kg body weight), or Yt (5 mg/kg body weight) diluted in PBS from day 4 and boosted every other day. The treatment schedule was continued until the tumors reached a volume of 2500 mm3. At this point, the animals were withdrawn from the study and euthanized. Blood samples from mice were collected for the blood counts and chemistry value testing. Two perpendicular diameters of the tumor were measured by a vernier caliper and the weight of the mice was determined every other day. The tumor volume was calculated using the formula V=L 1 L 2 2/2, where L 1 was the long diameter and L 2 was the short diameter. 11
Serum cytokine levels
Serum samples were collected, and TNF-α (sensitivity <20 pg/mL) and IL-2 (sensitivity <20 pg/mL) levels were measured by enzyme-linked immunosorbent assay (ELISA) using commercially available kits (Boster). Studies were performed according to the manufacturer's instructions. Optical density, with wavelength correction, was read on an automated microplate photometer. 12
Quantitative real-time polymerase chain reaction
Spleen samples were isolated from mice and discharged into TRIzol reagent (Invitrogen/Life Technologies). Total RNA was isolated and reverse-transcribed with ImProm-II™ Reverse Transcriptase (Promega). IFN-γ, IL2, IL4, IL10, TNF-α, TGF-β, GM-CSF, and GAPDH mRNA levels were determined by quantitative real-time polymerase chain reaction (qRT-PCR). Briefly, the primer sequences were designed and cDNA was amplified as previously described. 11 Relative quantitation of mRNA expression was calculated and normalized to an endogenous control (GAPDH) by the comparative threshold cycle (Ct) method.
Statistics
Data were analyzed using SigmaStat version 3.5, and first were checked for normality and transformed when necessary to meet the assumption of normal distribution. The contents of tumor size and cytokine expression among treatments were compared by one-way analysis of variance (ANOVA), followed by Tukey post hoc tests. All data are expressed as mean±SD.
Results
Preparation of Yp and Ys
Yt was extracted as previously described, 11 and contained proteins/peptides and low molecular weight compounds. These two fractions Yp and Ys were separated from Yt by chromatography over a column of ion-exchanger HP-20 and used in the subsequent animal study. About 5.4±0.3 mg Yp and 4.1±0.2 mg Ys could be obtained from 10 mg Yt. Gel stained with Coomassie blue showed that several major protein bands distributed from 12 to 66 kDa in the Yt lane, major protein bands from 12 to 35 kDa in the Yp lane, and few protein bands in the Ys lane (Fig. 1A).
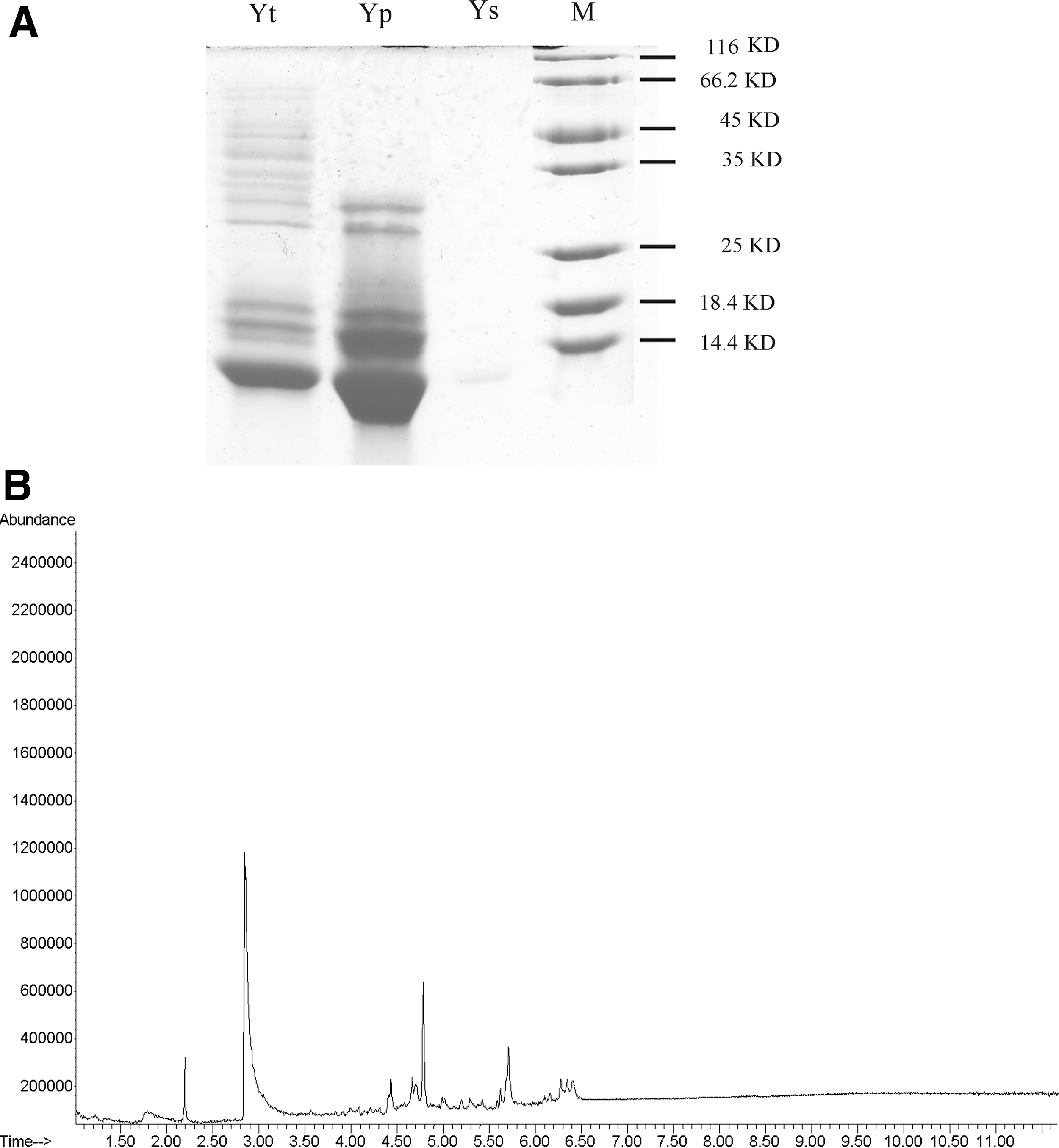
Preparation of Yp and Ys.
To further characterize the composition of Yp and Ys, mass spectrometry and GC-MS were performed respectively. As shown in Supplementary Fig. S1 (Supplementary Data are available online at
Therapeutic effects of Yp and Ys on tumor growth in vivo
The in vivo cancer prevention activity of Yp and Ys was examined on H22-bearing BALB/c mice. The tumor inhibition effects are shown in Figure 2. As expected, tumors in the negative control group (vehicle) rapidly grew from the seventh day after subcutaneous H22 tumors were established. The treatment of 2.5 mg/kg Yp, Ys, and 5 mg/kg Yt inhibited tumor growth significantly (Fig. 2A). Statistical analysis showed no significant differences between Yp and Yt, and significant differences between Yp and Ys (Fig. 2B). All animals were euthanized when tumors in the vehicle control group reached a volume of 2500 mm3. Figure 2C showed that the tumor tissues in Yp- and Yt-treated groups were very small, and were rejected in five mice of the Yt group and three mice of the Yp group. The lifespan of mice in the treated groups was significantly prolonged compared with that in control group, and all mice in the Yp group were alive, six mice in Ys died, and two in Yt died by the end of the study (Fig. 2D).

Yp and Ys inhibition of tumor growth with improvement of survival rate in H22-bearing BALB/c mouse model. H22 (5×105) cells were subcutaneously implanted into the right oxter of BALB/c mice. After 3 days, Yp, Ys, and Yt were intratumorally injected every 2 days.
Compared with that in control group, no significantly different trends were observed in hematology and blood chemistry values of 2.5 mg/kg Yp, 2.5 mg/kg Ys, and 5 mg/kg Yt-treated mice (Tables 3 and 4), except that the Yp-treated group showed a small increase in platelet distribution width, platelet larger cell ratio, and decrease in albumin. The Ys-treated group showed a small increase in white blood cells (WBCs) and glucose, and the Yt-treated group showed a small increase in WBCs, red blood cell counts, and glucose and decrease in albumin. The body weights of all four groups of mice gradually increased over the treatment time with no significant differences (Supplementary Fig. S1).
Values are reported as mean±SD.
P<.05.
PDW, platelet distribution width; P-LCR, platelet larger cell ratio; WBC, white blood cell.
Values are reported as mean±SD.
P<.05; ** P<.01.
All data indicated that both Yp and Ys could inhibit tumor growth significantly, and Yp performed better than Ys in antitumor and lifespan-prolonging activity.
Cytokine expression differed among the treated group and control group
In vivo, the function of IL-2 and TNF-α was reported to be closely associated to antitumor activity. 13,14 To study the effect of Yp and Ys on tumor-bearing mice, the serum was collected, and the expression of cytokine IL-2 and TNF-α were measured by ELISA. As shown in Figure 3A, the expression of IL-2 was significantly increased by Yt treatment (Control 38.9±27.6 pg/mL vs. Yt 77.4±39.7 pg/mL, and Yp 33.8±19.2 pg/mL vs. Ys 14.9±5.4 pg/mL; P<.05). TNF-α expression level was not significantly changed by treatments (Control 188.4±91.3 pg/mL, Yp 324.7±154.0 pg/mL, Ys 105.6±154.0 pg/mL, Yt 200.4±52.2 pg/mL; P>.05; Fig. 3B).
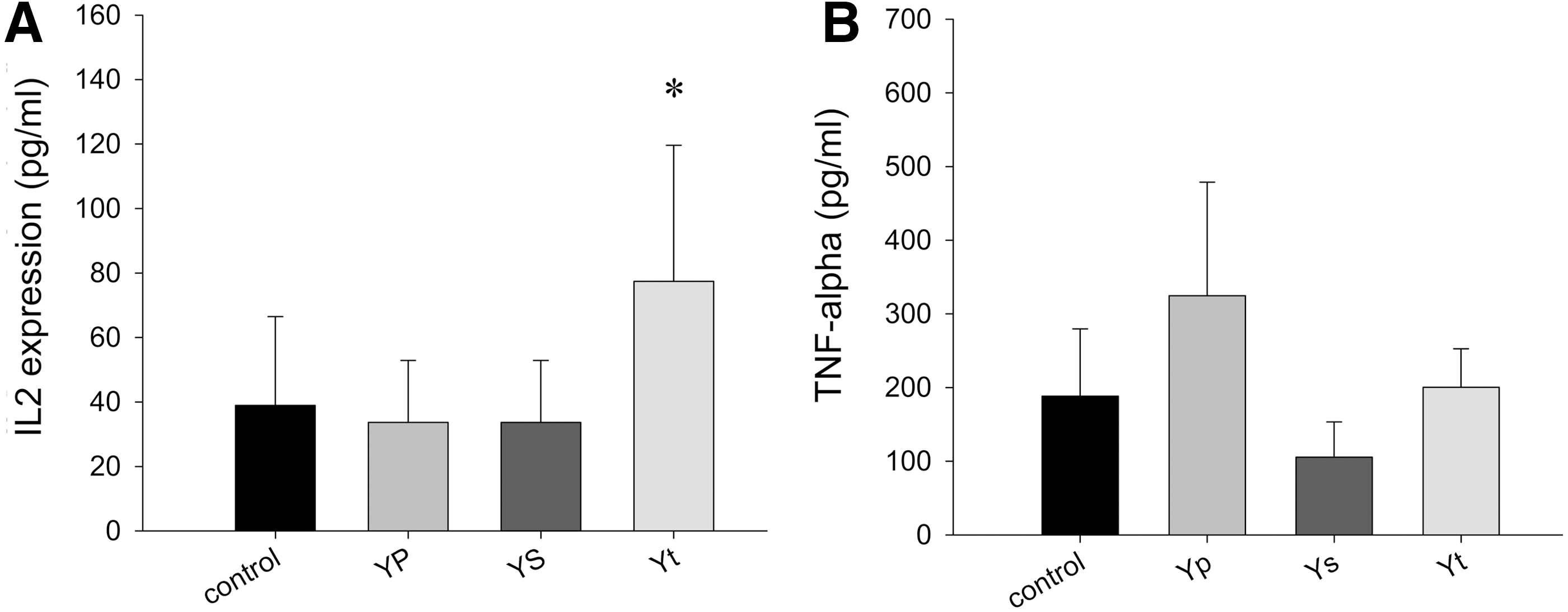
The cytokine IL-2
The mRNA levels of cytokines were regulated by Yp and Ys treatment
Although the protein expression of the cytokines did not change significantly, the mRNA expression level of cytokines was monitored by qRT-PCR (Fig. 4). The treatment of Yt upregulated IL-2 expression (10.9±2.5-fold, P<.01), TNF-α (2.33±1.3-fold, P<.05), GM-CSF (2.23±1.16-fold, P<.05), and IFN-γ (21.7±2.66-fold, P<.01) and downregulated the expression of TGF-β (4.5±0.1-fold, P<.01). Yp increased expression of IL-10 (9.77±2.12-fold, P<.01). Ys treatment upregulated the expression of GM-CSF (6.09±0.23-fold, P<.01) and TGF-β (1.77±0.05-fold, P<.01). Statistical analysis also revealed a significant difference of the effect on IL-2 (P<.05) and IL-10 (P<.01) between Yp and Yt, and the effect on IFN-γ between Ys and Yt (P<.001). Collectively, these results demonstrated that fraction-treatment regulated cytokine expression greatly, and that Yp and Ys inhibited tumor growth through different mechanisms.
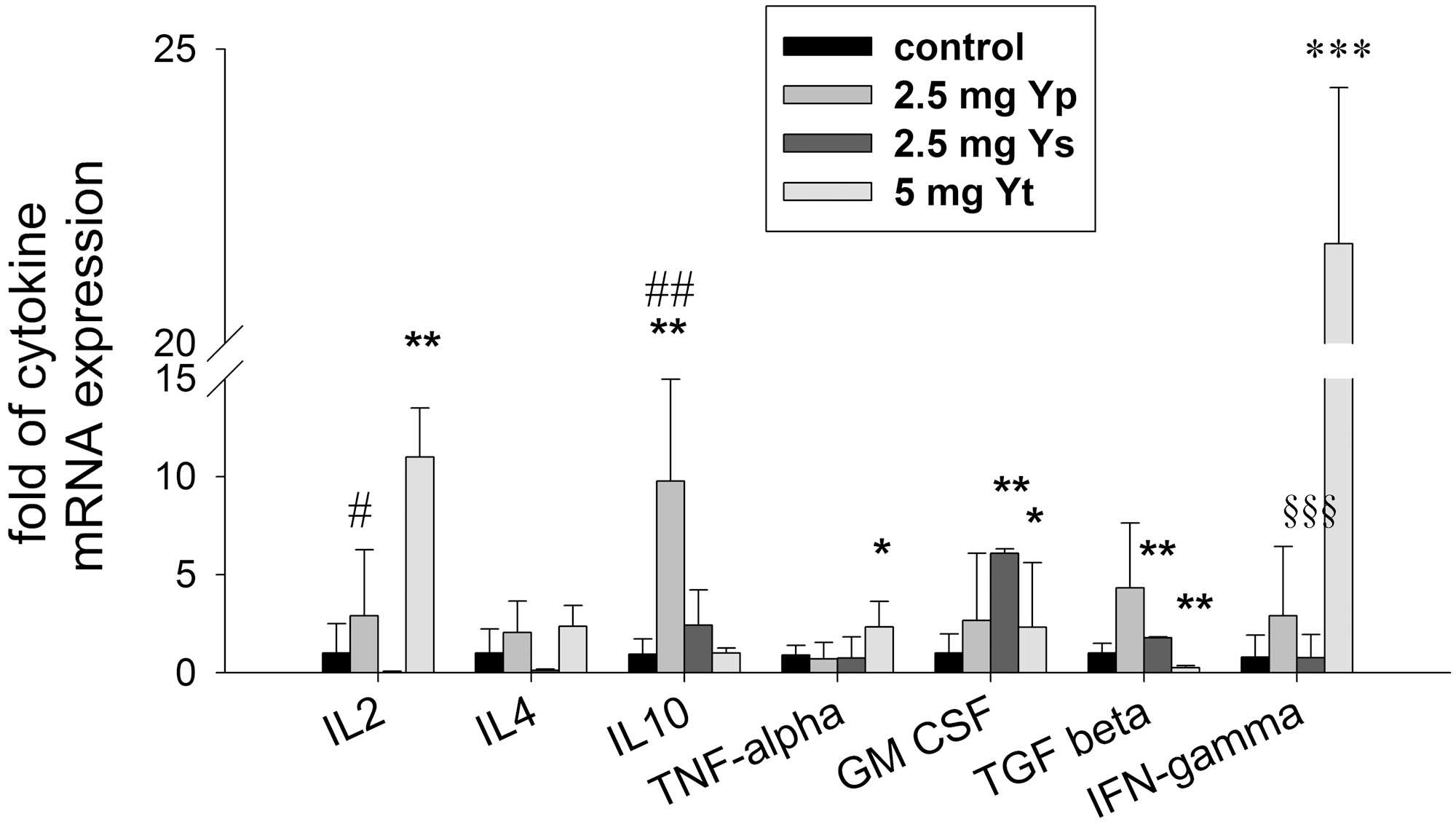
The mRNA levels of cytokines differed among treated group and control group. The mRNA levels were analyzed by quantitative real-time polymerase chain reaction. The experiment results were expressed as mean±SD of at least five mice. Statistical significance was determined by planned comparisons by one-way ANOVA with Tukey post hoc test, *P<.05; **P<.01, ***P<.001, treatments vs. control, # P<.05, ## P<.01, Yp vs. Yt, §§§ P<.001, Ys vs. Yt.
Discussion
The bioactive substances from Agrocybe mushroom mostly consist of protein-lectins, 15 –17 DNAase, 18 laccase, 19 ubiquitin-like peptide, 20 polysaccharides, 21 –25 indole, 26 sesquiterpenoid, 27,28 agrocybolacton, 29 and compound(s) with antibiotic properties. 30 The major antitumor components have been reported to be lectins and polysaccharides. Yt with trace polysaccharides derived from A. aegerita exhibited tumor-rejection activity in a previous study, which raised the question of the bioactivity compounds. 11 Here, the Diaion HP-20 column chromatography was used to separate protein and small molecule component fractions.
The MS results indicated that Yp contained AAL, AAL-2, serine proteinase, and other unknown proteins. AAL 17 and AAL-2 15 have been reported to have antitumor activity, and there were few reports about A. aegerita serine proteinase. An in vivo study showed that Yp induced tumor rejection and life span extension (Fig. 2), which was in agreement with the results of previous studies that the antitumor activity of Yt sharply declined when the proteins were degraded by proteinase K. 11 The existing knowledge of the bioactive protein/peptide from mushroom support that lectin, fungal immunomodulatory protein, DNases, RNases, and ribosome-inactivating proteins contribute antitumor and/or immunoregulation activity. 31 Taira described that a novel antimutagenic factor derived from Agrocybe cylindracea mushroom was extracted in water and was sensitive to heat treatment, which suggest that the component might be a protein/peptide. 32 These data also supported our results that proteins from medicinal mushroom possessed bioactivity and deserve better study. Besides AAL and AAL-2, the activity of other proteins in Yp needed further characterization.
Thin layer chromatography and customary fluorescent indicators were employed to define the constituents of Ys, and the results suggested that it may not contain flavonoid, anthraquinone, saponin, and terpenoid. GC-MS data showed that Ys mainly contained triethylene glycol, α-bisabolol, and n-hexadecanoic acid. Triethylene glycol was reported to have anti-infection 33 and reproductive toxicity 34,35 ; α-bisabolol exhibited antitumor activity in cell lines and animal models 36,37 and anti-inflammatory effects 38 ; n-hexadecanoic acid was found to inhibit phospholipase A(2) and control inflammation. 39 In a tumor-bearing mouse model, Ys showed high antitumor activity, which was possibly related to α-bisabolol and n-hexadecanoic acid. Yt was found to have reproductive toxicity in a previous study, 40,41 which may be associated to triethylene glycol. Neither indole nor sesquiterpenoid was identified in Ys, and it may be due to the extraction procedure or analysis approach.
Natural compounds found in mushrooms are well characterized for use as BRMS, and enhanced innate and cell-mediated immune response through regulation of cytokine production.
6
Fip-wo, a fungal protein isolated from Volvariella volvacea, exerted its immunomodulatory effect via upregulation of mRNA expression of IL-2, IL-4, IFN-γ, and TNF-α in mice.
42
Agaricus bisporus lectin (ABL) was reported to upregulate the mRNA expression of IL-1β and TNF-α in mouse splenocytes.
43
A low molecular weight protein fraction (MLP-fraction) from G. frondosa enhanced the production of IL-12 and IFN-γ by splenocytes in colon-26 carcinoma-bearing mice.
44
IFN-γ, TNF-α, and GM-CSF expressions in splenocytes of DBA/2 mice was altered by SCG, a 1,3-β-
Medicinal mushrooms are valuable potential sources for natural bioactive products. We separated the protein component fraction Yp and low molecular weight component fraction Ys from crude extract Yt, which exhibited tumor rejection activity, and found that both Yp and Ys had significant antitumor activity through regulating cytokine expression, especially Yp. Compared with the low molecular weight component fraction Ys, the protein component fraction Yp had higher antitumor efficacy and lower toxicity. Further purification and definition of the constituents coupled with investigation of the mechanism at molecular and cellular levels will be helpful to elucidate the unique and advantageous effect of medicinal mushrooms.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of China (Program no. 81102850), Scientific Research Program for Educational Commission of Guangdong Province (LYM11070), Medical Scientific Research Foundation of Guangdong Province (A2011434), Higher Education Institutions of Dongguan (Program no. 2011108102049), and Science and Technology Foundation of Zhanjiang (2011C3109015).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.