
Abstract
This study was performed to investigate the hypolipidemic, antiobese, and antiatherogenic effects of resveratrol in apoE-deficient mice fed an atherogenic diet (20% fat and 1% cholesterol). These animals were fed an atherogenic diet containing 0.02% lovastatin (w/w) or 0.02% resveratrol (w/w) for 12 weeks. Resveratrol and lovastatin supplementation significantly reduced either the body weight or epididymal fat weight without altering the food intake and food efficiency ratio. Resveratrol significantly decreased the plasma total cholesterol (total-C), low-density lipoprotein cholesterol (LDL-C), non-high-density lipoprotein cholesterol (non-HDL-C) concentrations, apoB/apoA-I ratio, hepatic cholesterol, and triglyceride (TG) contents, whereas significantly it increased the plasma HDL-C concentration compared with the control and lovastatin groups. Plasma and hepatic TG and plasma apoB levels were significantly lower in both the lovastatin and resveratrol groups than in the control group without altering the plasma apoA-I concentration. Both resveratrol and lovastatin significantly decreased hepatic fatty acid and TG synthesis, whereas they increased fatty acid oxidation (β-oxidation) except for the carnitine palmitoyltransferase activity compared with the control group. However, there was no difference in hepatic 3-hydroxyl-3-methylglutaryl-CoA reductase activity among the groups, although hepatic acyl-CoA: cholesterol acyltransferase activity was significantly lower in the lovastatin groups than in the control group. In epididymal adipose tissue, resveratrol supplementation led to an increase in β-oxidation and decrease in TG synthesis, compared with the control group. Tissue morphology revealed that there were dramatic decreases in hepatic lipid droplets and aortic fatty streaks by resveratrol and lovastatin supplementation. This study demonstrates that resveratrol exerts not only antiobesity and hypolipidemic effects, but also protective effects for the liver and aorta through the modulation of lipid metabolism in both the liver and white adipose tissues.
Introduction
C
Many studies have searched for phytochemicals possessing hypolipidemic and antiobesity capacities. 4,5 Most of all, resveratrol is a naturally occurring phytochemical in grapes, peanuts, and other plants, and it has been demonstrated to improve CVD, 6 obesity, 7 diabetes, 8 cancer, 9 and to regulate angiogenesis 10 as well as to ameliorate the aging process 11 since its bioactivity was first elucidated in the 1990's. Lovastatin, a 3-hydroxyl-3-methylglutaryl-CoA (HMG-CoA) reductase inhibitor, is a member of the statin drug class used for lowering cholesterol in hypercholesterolemic patients. 12 Among the experimental animal models, apoE-deficient mice exhibit severe hypercholesterolemia and atherosclerotic lesions because apoE is an important apolipoprotein with antiatherosclerotic functions. Thus, this animal model has been widely used in studies on experimental atherosclerosis as an ideal model of human atherosclerosis. 13,14 This study was performed to assess the hypolipidemic, antiobese, and antiatherosclerotic effects of resveratrol compared with lovastatin in apoE-deficient mice.
Materials And Methods
Animals and diets
Four-week-old male homozygous apoE-deficient mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA). After a 1-week adaptation period, thirty apoE-deficient mice were randomly divided into three groups. The control group was provided with an atherogenic diet (20% fat and 1% cholesterol) based on an AIN-76 semisynthetic diet, while the two experimental groups received an atherogenic control diet supplemented with 0.02% (w/w) lovastatin (Chong Kun Dang Pharm., Seoul, Korea) or 0.02% (w/w) resveratrol (Sigma Chemical Co., St Louis, MO, USA) for 12 weeks (Table 1). Body weight and food intake were measured every week and every day, respectively. After 12 weeks, blood was collected from the inferior vena cava to determine the plasma biomarkers. Plasma was prepared by centrifugation at 1500 g, 4°C for 10 min. The liver, fat pads (epididymal and perirenal fat), and aorta were removed, rinsed, and weighed. All samples were stored at −70°C until analysis. The current study protocol was approved by the Ethics Committee at Kyungpook National University for animal studies.
AIN-mineral, AIN-76 mineral mixture (Harlan Teklad Co. Madison, WI, USA); AIN-vitamin, AIN-76 vitamin mixture (Harlan Teklad Co.).
Plasma and hepatic lipid profiles
The plasma triglyceride (TG), total-C and HDL-C concentrations were quantitatively determined using the respective commercial kits (Asan Pharm a Company, Hwasung, Republic of Korea) based on the enzymatic reactions. Plasma apoA-I and apoB levels were also measured using the respective enzymatic kits (Eiken, Tokyo, Japan). The lipid fraction in the liver was extracted with a procedure developed by Folch et al., 15 and the hepatic cholesterol and TG contents were analyzed with the same enzymatic kits used in the plasma analyses.
Lipid metabolic enzyme activities in the liver and epididymal fat pad
To measure the lipid-regulating enzyme activities in the liver and epididymal white adipose tissue (WAT), cytosolic, mitochondrial, and microsomal fractions were prepared according to the method by Hulcher and Oleson. 16 The protein concentrations in each fraction were measured according to Bradford's method 17 using bovine serum albumin as the standard. The cytosolic fatty acid synthase (FAS) activity was measured according to Nepokroeff's method 18 by monitoring the malonyl-CoA-dependent oxidation of the NADPH at 340 nm, and malic enzyme (ME) activity was measured according to Ochoa's method 19 by monitoring the production of NADPH at 340 nm. The glucose-6-phosphate dehydrogenase (G6PD) activity was assayed by spectrophotometric methods according to the procedures described by Pitkanen et al., 20 and the phosphatidate phosphohydrolase (PAP) activity was determined using Walton's method. 21 The mitochondrial carnitine palmitoyltransferase (CPT) activity was analyzed using Markwell's method, 22 and fatty acid β-oxidation was determined using Lazarow's method 23 by monitoring the reduction of NAD to NADH at 340 nm.
Hepatic HMG-CoA reductase and acyl-CoA:cholesterol acyltransferase (ACAT) activities were determined as described by Shapiro et al. 24 and by Gillies et al., 25 respectively.
Histopathological analysis in the liver and aorta
The liver and aorta were fixed in 10% paraformaldehyde/PBS, embedded in paraffin, and stained with hematoxylin and eosin (H&E). To determine the presence of lipids, an adjacent section of the aortic arch was cryosectioned and stained with the Oil Red O (ORO) solution. The stained area was then viewed using a microscope at a magnification of 200×.
Statistical analysis
The data are expressed as the mean±standard error (SE). Significant differences among the groups were determined by one-way ANOVA using the SPSS program (SPSS, Inc., Chicago, IL). The differences between the means were assessed using the Duncan's multiple range test and statistical significances were considered at P<.05.
Results
Body weight gain, food intake, and organ weight
Body weight gain was significantly lower in both the resveratrol and lovastatin groups compared with the control group, whereas the food intake and food efficiency ratio were not different between the groups (Table 2). When comparing the organ weights, resveratrol and lovastatin supplementation resulted in significant reductions in epididymal WAT weight without the differences in the liver and perirenal WAT weights (Table 2).
Data are the mean±SE, n=10.
Means in the same row not sharing a common superscript are significantly different between the groups at P<.05.
FER, food efficiency ratio=body weight gain/food intake; WAT, white adipose tissue.
Plasma and hepatic lipid profiles
Both total-C and LDL-C concentrations in plasma were significantly lower in the resveratrol group than in the control group after 12 weeks, whereas plasma TG concentration was significantly lower in both the lovastatin and resveratrol groups than in the control group (Table 3). Resveratrol supplementation resulted in significantly higher plasma HDL-C concentrations compared with the control and lovastatin groups without any differences in the plasma apoA-I concentrations (Table 3). The plasma apoB concentration and apoB/apoA-I ratio were significantly lower in both lovastatin and resveratrol groups than in the control group (Table 3). Consequently, the resveratrol group exhibited lower levels of all atherogenic factors, including plasma total-C, LDL-C, TG, and apoB concentrations and the apoB/apoA-I ratio (Table 3).
Data are the mean±SE. LDL-cholesterol=(Total-cholesterol)−(HDL-cholesterol)−(Triglycerides/5).
Means not sharing a common letter are significantly different among groups at P<.05.
ApoA-I, apolipoprotein A-I; apoB, apolipoprotein B.
Hepatic lipid analyses revealed that the resveratrol group had significantly lower levels of both cholesterol and TG, whereas the lovastatin group only had lower TG content, compared with the control group (Table 3).
Lipid metabolism-related enzyme activities in the liver and epididymal WAT
The results of the lipid metabolism-related enzyme activities in the liver and epididymal fat pad are presented in Table 4. In the liver, the activities of adipogenic enzymes, including G6PD, FAS, ME, and PAP were all significantly lower in both the lovastatin and resveratrol groups than in the control group. When compared with the control group, the hepatic mitochondrial fatty acid β-oxidation was significantly elevated by lovastatin and resveratrol supplementation without differences in the activity of CPT, the enzyme acting at the first step of fatty acid oxidation. Hepatic HMG-CoA reductase activity was not significantly different among the groups, while the activity of ACAT, an esterification enzyme of free cholesterol, was significantly lower in the lovastatin group than in the control group (Table 4).
Data are the mean±SE.
Means not sharing a common letter are significantly different among groups at P<.05.
G6PD, glucose-6-phosphate dehydrogenase; ME, malic enzyme; FAS, fatty acid synthase; CPT, carnitine palmitoyl transferase; PAP, phosphatidate phosphohydrolase; HMG-CoA reductase, 3-hydroxyl-3-methylglutaryl coenzyme A reductase; ACAT, acyl-CoA: cholesterol acyltransferase.
Unlike the results of the hepatic lipid-regulating enzyme activities, there were no significant differences in the epididymal enzyme activities of G6PD, ME, FAS, and CPT among the groups. However, resveratrol supplementation resulted in significantly higher epididymal β-oxidation and lower PAP activities, respectively, compared with the control group.
Histopathological changes in the liver and aorta
The effects of resveratrol on the pathogenesis of steatosis and atherosclerosis were evaluated in apoE-deficient mice. As shown in Figure 1, both lipid droplet numbers and their sizes were higher due to the accumulation of hepatic lipids in the control group, whereas the lovastatin and resveratrol groups showed much less and smaller lipid droplets (Fig. 1A). Figure 1B and C show the representative H&E- and ORO-stained histological differences of the aortas from the experimental groups. Dietary resveratrol and lovastatin exposure remarkably inhibited the endothelial damage and fatty streak formation shown by the H&E staining of the aortic arch. ORO staining revealed that resveratrol and lovastatin significantly inhibited fatty plaques and lesion formation by lipid deposition in the aortic arch when compared with the control group, which was fed an atherogenic diet.
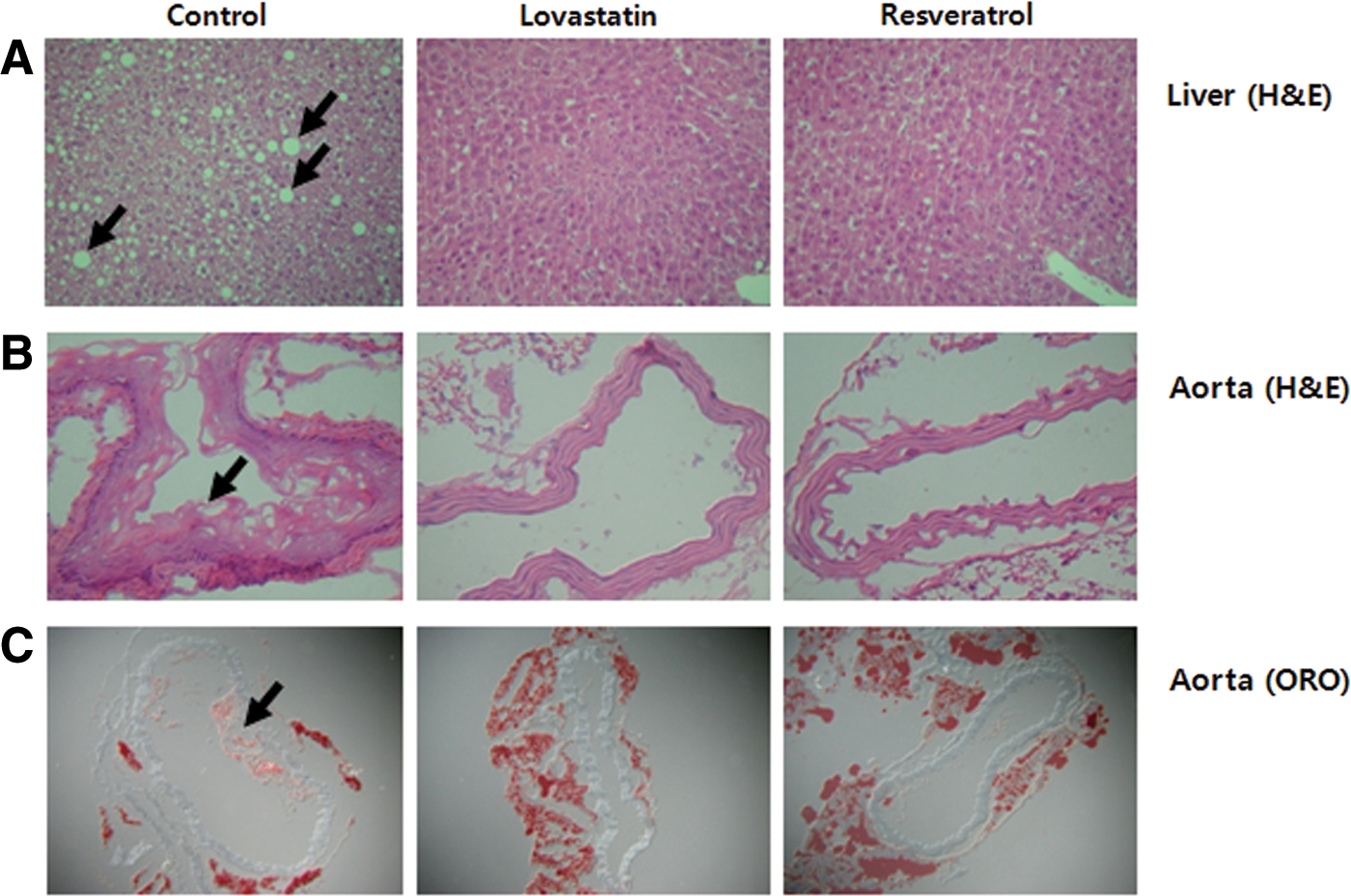
Effects of resveratrol supplementation for 12 weeks on the liver
Discussion
The ApoE-deficient mice used in this experiment are one of the hypercholesterolemic animal models that have also been shown to mimic the initiation and progression of human atherosclerosis. 26 This study demonstrated that dietary resveratrol in apoE-deficient mice resulted in hypolipidemic, antiobese, and antiatherosclerotic effects by improving lipid metabolism in the plasma, liver, and WAT. Several studies have provided evidence regarding the potential antiobesity effects of resveratrol, which reduced the body weight, liver weight, and/or adiposity in animals fed a high-fat diet or in obese Zucker rats fed a normal diet. 27 –29 The present results showed a similar body weight and adiposity-lowering effects of resveratrol without any differences in the liver weight.
Regarding the hypolipidemic effects of resveratrol, Cho et al. 30 observed decreases in the amount of both plasma and hepatic lipids with decreased hepatic HMG-CoA reductase activity and mRNA expression in hamsters fed a high-fat diet. Both Soleas et al. 31 and Goldberg et al. 32 indicated that trans-resveratrol treatments significantly decreased the intracellular concentration of apoB, resulting in less VLDL and LDL production in a HepG2 cell line. Daily consumption of resveratrol (10 mg/kg of body weight) significantly decreased the TG and cholesterol concentrations in both the plasma and liver of hyperlipidemic obese Zucker rats when compared with the control group. 28 In the present study, the amount of daily resveratrol consumed by the apo E-deficient mice was approximately 28.5 mg/kg body weight based on food intake, that is equivalent to 85.5 mg resveratrol/60 kg man based on the conversion of the mice dose to a human equivalent dose using the body surface area normalization method. 33 As previously discussed, our study also showed a significant decrease in the plasma total cholesterol, LDL-C, TG, and apoB concentrations as well as hepatic cholesterol and TG accumulation by resveratrol supplementation. The plasma HDL-C concentration was also increased by resveratrol in the present study, which corresponds with the report by Penumathsa et al. 34 The National Cholesterol Education Program (NCEP) update recommended the use of non-HDL-C as a target for individuals with high TG (≥200 mg/dL). However, some patients who achieve a significantly lowered LDL-C or are within the recommended LDL-C range still develop CV events. 35 Some epidemiological studies have indicated that higher the apoB/apoA-I ratio, the higher the CV risk is because high TG and low HDL-C are also part of the atherogenic lipid profile. 35,36 A report by Do et al. 37 showed the spontaneous HDL-C-raising and TG-lowering effects of resveratrol in normal diet fed apoE-deficient mice. In general, there is an inverse relationship between plasma TG and HDL-C, 38 and the results of this study show a substantial decrease in plasma TG coupled with an increase in HDL-C as well as an increase in the apoB/apoA-I ratio. On the other hand, several studies suggested that resveratrol administration did not alter the lipid profile. This includes the plasma total-C, LDL-C, and TG as well as HDL-C in rats receiving either a standard or high-fat diet. 39,40 Interestingly, lovastatin, used as a positive control, was not effective in lowering the plasma and hepatic cholesterol levels in this particular atherogenic animal model. However, it significantly decreased plasma and hepatic TG levels, although it is one of the most frequently used cholesterol-lowering agents. A study by Xu et al. 41 suggested that simvastatin supplementation with a high cholesterol diet significantly lowered plasma TG, but not the LDL-C. Steinmetz et al. 42 also suggested that treatment with simvastatin had no effect on the serum cholesterol levels in either normal or hypercholesterolemic mice.
The antiadipogenic activity of resveratrol has been elucidated in several experiments. Studies on isolated cells revealed the inhibition of adipogenesis in the presence of resveratrol. 43,44 Resveratrol (10–100 μM) was found to inhibit fatty acid synthesis from acetate in isolated rat hepatocytes. This effect was accompanied by the lessened activity of acetyl-CoA carboxylase (ACC) and decreased incorporation into TG, but the FAS and HMG-CoA reductase activities were unchanged. 45 The biological benefits of resveratrol have also been suggested to involve AMPK, which prevents the development of hyperlipidemia, atherosclerosis, and diabetes by increasing glucose uptake 46 and by inhibiting fatty acid and/or TG synthesis 45,47 as well as cholesterol synthesis. 48 In addition to the results discussed above, the present study also showed evidence for not only decreased hepatic FA and TG synthesis, but also for increased β-oxidation from resveratrol. Some reports confirmed that resveratrol increases the mitochondrial content of the liver. 40,49 It seems that the extra mitochondria content induced by resveratrol may affect the improvement in hepatic lipid metabolism, especially mitochondrial β-oxidation, although the mitochondrial contents were not measured directly in this study. Results suggest that resveratrol consumption is effective in enhancing FA oxidation in both the liver and WAT, and a significant lowering of hepatic TG content was partially due to the improvement in overall lipid-regulating enzyme activities. A high-fat and cholesterol-containing diet induces the accumulation of hepatic cholesterol and TG as well as an increase in liver weight. 50 In this study, the ingestion of resveratrol with an atherogenic diet significantly lowered the hepatic cholesterol and TG contents and hepatic lipid droplets without altering the liver weight in apoE-deficient mice.
However, hepatic cholesterol biosynthesis and cholesterol esterification were not influenced by resveratrol, although it did cause a significant decrease in plasma total-C, LDL-C, and apoB secretions, which corresponded with the report by Gnoni. 45 In addition, this result seems to be due to hepatic cholesterol homeostasis. However, in high-fat fed hamsters, resveratrol was effective in attenuating hepatic HMG-CoA reductase activity and mRNA expression. 30 The endothelium plays an important role in the maintenance of vascular homeostasis. It is well known that endothelial damage disturbs vascular homeostasis, following the initiation of the atherosclerotic process. 51 Several studies have suggested that resveratrol contributes to the prevention of atherosclerosis by inhibiting aortic vascular smooth muscle cell proliferation and endothelial foam cell formation. 52,53 Resveratrol and lovastatin significantly attenuated the atherosclerotic plaque and endothelial lesions to which cholesterol-fed apoE-deficient mice were subjected. Resveratrol preserved the endothelial lining and exerted vasoprotective effects. Clinical trials have demonstrated that statins exert antiatherosclerotic effects that are independent of their cholesterol-lowering properties. 54,55 Consistent with the results mentioned above, lovastatin treatment was also found to be effective in preventing atherosclerotic lesions without changing the plasma or hepatic cholesterol levels.
The results suggest that resveratrol supplementation improves dyslipidemia and lowers the fat pad mass and body weight, possibly by regulating the hepatic lipoprotein/apolipoprotein secretion, hepatic and adipocytic lipid metabolism, including FA synthesis and/or oxidation and TG synthesis, and attenuates steatosis and atherosclerotic lesion formation. Accordingly, it is considered that resveratrol may possess antiobese, hypolipidemic, and antiatherosclerotic capacities along with steatosis-preventing effects against diet-induced metabolic disorders in apoE-deficient mice under atherogenic conditions.
Footnotes
Acknowledgment
This work was funded by the Basic Science Research Program (Center for Food & Nutritional Genomics: grant number 2012-0000644, 2012M3A9C4048818) of the National Research Foundation (NRF) of Korea funded by the Ministry of Education, Science and Technology.
Author Disclosure Statement
No competing financial interests exist.