
Abstract
The purpose of this study was to investigate how proliferation and apoptosis in human leukemia U937 cells are affected by four hydroxycinnamic acid derivatives (HCADs) in corn (Zea mays L.) bran: p-coumaric (CA), ferulic acids (FA), dicoumaroylputrescine (DCP), and diferuloylputrescine (DFP). Of the four HCADs, DFP dose dependently exerted the strongest cytotoxic effect and induction of apoptosis in the U937 cells. In addition, DFP induced distinct morphological changes characteristic of cellular apoptosis, such as chromatin condensation, apoptotic bodies, and DNA fragmentations. The DFP-induced apoptosis was also associated with released cytochrome c in the cytosol with activation of caspase 3, together with the downregulation of anti-apoptotic proteins, including XIAP and cIAP2, Bcl-2, and Mcl-1. Finally, the DFP-induced apoptosis was a cell-specific response in leukemia cells, as compared with those of other cancer cells, such as Caki, HT29, SK-Hep1, and MDA-MB231. Thus, these results suggest that DFP may be useful as a potential source of natural antileukemic agents.
Introduction
A
Corn (Zea mays L.) bran, which is obtained as a byproduct during the corn dry-milling process, is rich in several functional constituents, including unsaturated fatty acids, tocopherols, phytosterols, dietary fibers, carotenoid pigments, and so on. 10 Recently, there has been renewed interest in corn bran as a functional phytochemical source due to its high levels of HCADs, including p-coumaric acid (CA), ferulic acid (FA), and their phenolic amides, such as diferuloylputrescine (DFP), p-coumaroylferuloylputrescine (CFP), and dicoumaroylputrescine (DCP). 11 Phenolic amides in corn bran have been recently reported to have many biological effects, including antidiabetic, antimicrobial, antioxidant and antimelanogenic, and anti-inflammatory activities. 12 –15 However, little is known about the anticancer activity of HCADs in corn bran.
Naturally occurring polyphenolic compounds, such as phenolic acids, cinnamic acids, flavonoids, coumarins, stilbenes, lignans, and tannins, have received much attention as potential chemopreventive and chemotherapeutic agents. 16,17 They have different mechanisms of action, including cell growth suppression, modulation of cell differentiation, and induction of apoptosis. 18 In particular, cinnamic acid derivatives, such as ferulic acid and caffeic acid, are attracting considerable interest as potential chemopreventive agents without higher cellular toxicity. 19,20 However, information on the anticancer activity of phenolic amides with a cinnamic acid moiety is very limited.
In the present study, we evaluated the effects of four HCADs in corn bran on proliferation and apoptosis in human monocytic leukemia U937 cells using the XTT assay and flow cytometry analysis. Furthermore, the apoptosis events induced by DFP showing the strongest antiproliferative activity, including characteristic morphological changes, DNA fragmentation, apoptotic gene expression, and cytochrome c release, were examined by fluorescence microscopy, DNA fragmentation assay, and Western blot analysis.
Materials and Methods
Materials
Corn bran obtained after the dry milling of corn was purchased from Hankook Goksan Co. (Goyangsi, Gyeonggido, Korea). The two phenolic amides, DFP and DCP, were isolated from the corn bran, as previously described. 14 The defatted 80% aqueous ethanol extract of corn bran powder was fractionated with CH2Cl2, and then successively chromatographed over silica gel, ODS-A, and Sephadex LH-20 to obtain the two phenolic amides.
Chemicals
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS),
Cell culture and cytotoxicity assay
U937, Caki, HT29, SK-Hep1, MDA-MB231 cells were obtained from the American Type Culture Collection (Rockville, MD, USA). The mesangial cell (MC) and the normal human skin fibroblasts (HSF) were a gift from Dr. T.J. Lee (Yeungnam University, Korea). U937 and HT29 cells were grown at 37°C in a humidified atmosphere with 5% CO2 in RPMI 1640 media supplemented with 10% heat inactivated FBS and 1% antibiotics–antimycotics. Caki, SK-Hep1, MDA-MB231, MC, and HSF cells were grown at 37°C in 5% CO2 atmosphere in DMEM, supplemented with the same concentrations of serum and antibiotics–antimycotics as stated above. The cytotoxic effects of the four HCADs were measured using the XTT assay described by Hansen et al. 21 Briefly, the cancer cell lines and normal cells were seeded on a 96-well plate (1.5×104 cells/well) and treated with test samples at various concentrations for 24 h. The culture solutions were then treated with 20 μL/well XTT reagent containing PMS (phenazine methosulfate) for 2 h and measured spectrophotometrically at 450 nm using a microplate reader. The cytotoxic effects were expressed as the percentage of cell growth compared to control nontreated cells (for which growth is considered 100%) and was calculated by A 450nm (sample)/A 450nm (control)×100. The experiment was repeated five times using three wells for each HCAD concentration.
Flow cytometry analysis for measurement of sub-G1 phase
The effect of HCAD-induced apoptosis was measured by flow cytometry as described by Oh et al. 22 U937 cells were either untreated or treated with 50 or 100 μM HCADs for 24 h. After these treatments, ∼106 cell pellets were prepared and washed twice with phosphate-buffered saline (PBS) without Ca2+ plus 2% FBS, suspended in 100 μL of PBS, and 200 μL of 95% cold ethanol was added while vortexing. The cells were incubated at 4°C for 1 h, washed twice with PBS containing 2% FBS and resuspended with 12.5 μg of RNase in 250 μL of 1.12% sodium citrate buffer (pH 8.45). Incubation was continued at 37°C for 30 min before staining of the cellular DNA with 250 μL of propidium iodide (50 μg/mL) for 30 min at 4°C. The stained cells were analyzed on a fluorescent activated cell sorter (FACS) flow cytometer (Becton & Dickinson, San Jose, CA, USA) for the relative DNA content, based on an increased red fluorescence.
Nuclear staining and morphological analysis
U937 cells (2.5×105 cells/well) were cultured in 24-well plates in RPMI 1640 medium containing 10% FBS in the absence or presence of 50–200 μM DFP. After 24 h, cells were washed with PBS and stained with the DNA specific 300 nM 4′,6-diamidine-2′-phenylindole dihydrochloride (DAPI; Sigma) solution for 30 min at 37°C. The cells were then washed twice with PBS and observed under fluorescence microscopy (Olympus, Tokyo, Japan).
DNA fragmentation assay
DNA fragmentation assay was measured according to the method of Herrmann et al. 23 Cells were treated with various concentrations of DFP at a range of 50–200 μM for 24 h. After harvesting, the cell samples were washed with PBS and pelleted by centrifugation. The cell pellets were then treated for 30 min with lysis buffer (1% NP-40 in 20 mM EDTA, 50 mM Tris-HCl, pH 7.5; 50 μL per 106 cells). After centrifugation for 10 min at 1600 g, the supernatant was collected, and the extraction was repeated with the same amount of lysis buffer. The supernatants (and the resuspended nuclei as control for the complete recovery of the apoptotic DNA fragments) were brought to 1% SDS and treated for 2 h with RNase A (final concentration 5 μg/μL) at 56°C and subsequently with proteinase K for 2 h at 37°C. After the addition of 1/2 vol. 10 M ammonium acetate, the DNA was precipitated with 2.5 vol. ethanol, dissolved in gel loading buffer, and separated by electrophoresis in 1.5% agarose gels.
Analysis of cytochrome c release
A total of 2×104 cells were harvested, washed once with ice-cold PBS, and gently lysed for 2 min in 80 μL lysis buffer (250 mM sucrose, 1 mM EDTA, 20 mM Tris-HCl [pH 7.2], 1 mM DTT, 10 mM KCl, 1.5 mM MgCl2, 5 μg/mL pepstatin A, 10 μg/mL leupeptin, and 2 μg/mL aprotinin). Lysates were centrifuged at 1600g at 4°C for 10 min to obtain the supernatants (cytosolic extracts free of mitochondria) and pellets containing mitochondria. The cytosolic fractions were used for Western blot analysis with an anticytochrome c antibody.
Western blotting
Western blotting of the cell lysates was performed as previously described. 24 Cells were treated with various concentration of DFP ranging from 50 to 200 μM for 24 h. Cells were collected and lysed in ice-cold lysis buffer (0.5 M Tris-HCl [pH 7.5], 10 mM EDTA, 1% Triton X-100, and 100 μM phenylmethylsulfonyl fluoride). The cells were then disrupted by sonication and extracted at 4°C for 30 min. The protein concentrations of the extracts were estimated using the Bio-Rad protein assay (Bio-Rad Laboratories, Hercules, CA, USA) with bovine serum albumin as a standard. Total proteins (30–50 μg) were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) using 8% polyacrylamide gel. The protein in the gel was transferred to nitrocellulose membranes (Milipore Corporation, Bedford, MA, USA). The membranes were blocked with 5% skim milk in PBST (0.05% v/v Tween-20 in PBS, pH 7.2) for 1 h. The membranes were incubated with a primary antibody (1:1000) overnight at 4°C, and then with a secondary antibody (1:1000) conjugated with horseradish peroxidase for 1 h at room temperature. The membranes were washed in PBST three times for 10 min. The bands were detected using the Amersham ECL system (Amersham-Phamacia Biotech, Arlington Heights, IL, USA).
Statistical analysis
All data are presented as mean±standard deviation (SD) from at least three separate experiments. The data were analyzed with one-way analysis of variance (ANOVA) followed by post hoc comparisons (Student–Newman–Keuls) using the SPSS v8.0 (SPSS, Inc., Chicago, IL, USA).
Results and Discussion
Cytotoxic effect of the four HCADs on the growth of U937 cells
Four HCADs in corn bran (CA and FA, and their phenolic amides, DCP and DFP) have been reported to possess many biological effects, such as antidiabetic, antimicrobial, antioxidant, antimelanogenic, and anti-inflammatory activities. 12 –15 Currently, there is great interest in the potential biological activity of phenolic amides derived from natural sources, but little is known about the anticancer activity of phenolic amides from corn bran. Therefore, the antiproliferative effects of the four HCADs in corn bran on U937 cells determined by XTT assay are shown in Figure 1. DFP significantly reduced the viability of U937 cells in a dose-dependent manner at concentrations of 50 and 100 μM. In particular, DFP (51%) showed a potent cytotoxic effect at 100 μM, while DCP, CA, and FA were less cytotoxic than DFP. DFP was a predominant phenolic amide in corn bran. 11 The cytotoxic effects of DFP in the range of concentration was similar to those of resveratrol derivatives reported by Kang et al. 25 and epigallocatechin gallate (EGCG) reported by Achiwa et al. 26 in leukemia cells.
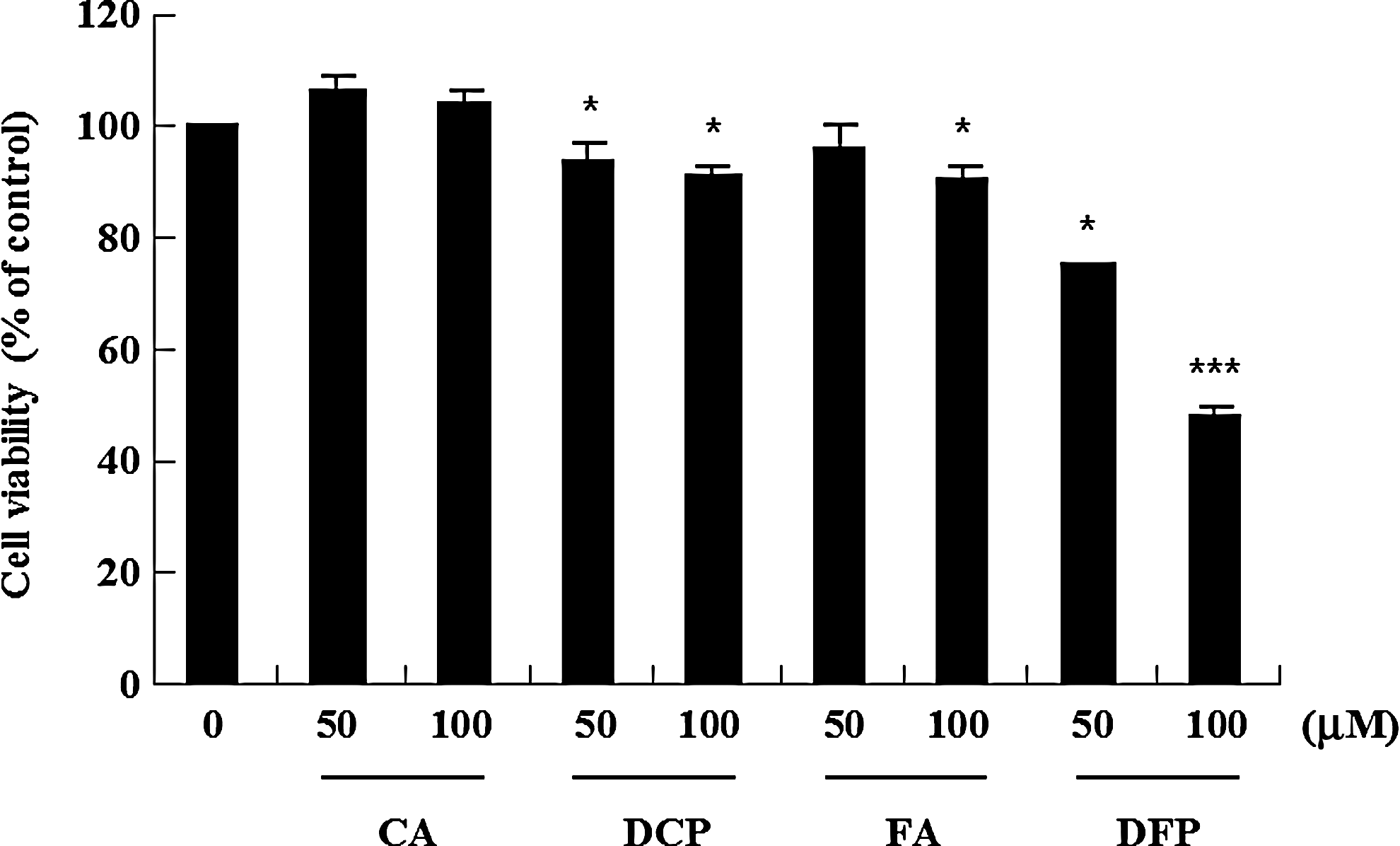
Cytotoxic effect of the four hydroxycinnamic acid derivatives (HCADs) on the growth of U937 cells. The cells were seeded in 96-well plates and cultured in triplicate in the presence or absence of the four HCADs (50 and 100 μM) for 24 h, and the viabilities of the cells were determined by XTT assay. Cell survivals are shown as a percentage untreated of control cells and are the mean±standard deviation (SD) from three independent experiments. *P<.05, ***P<.001 compared to control. CA, p-coumaric acid; DCP, dicoumaroylputrescine; FA, ferulic acid; DFP, diferuloylputrescine.
Effect of the four HCADs on apoptosis in U937 cells
To determine the effect of the four HCADs on apoptotic cell death, U937 cells were treated with four different HCADs at 50 and 100 μM for 24 h, and hypodiploid cell populations were quantified by flow cytometric analysis. As shown in Figure 2, the four HCADs increased sub-G1 population in a dose-dependent manner, and 100 μM DFP markedly increased the accumulation of sub-G1 phase cells by 29.3%, whereas FA, DCP, and CA treatments resulted in fewer sub-G1 cells than DFP. These results indicate that of the four HCADs, DFP most potently induced apoptotic death of human leukemia cells. Therefore, based on the two results above, a more detailed investigation of the apoptotic events induced by DFP was undertaken through morphological and biochemical analyses.
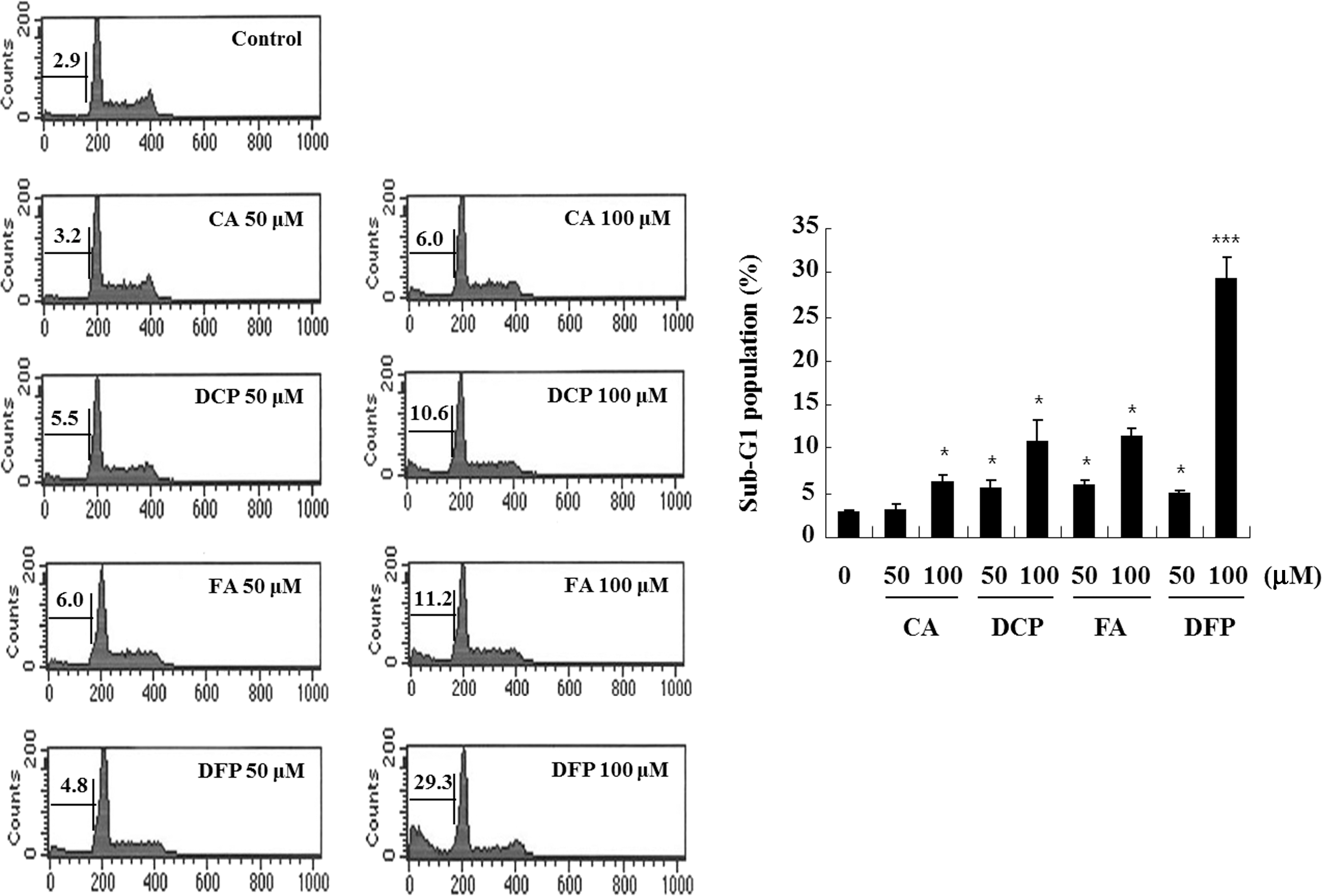
Effect of the four HCADs on apoptosis in U937 cells. Cells were treated with 50 and 100 μM the four HCADs for 24 h, and then evaluated for DNA content after propidium iodide staining. The percentages of the sub-G1 phases are indicated. Data represent the mean±SD of three independent experiments. *P<.05, ***P<.001 compared to control.
Effect of DFP on morphological changes and DNA fragmentation in U937 cells
After U937 cells were treated with various concentration of the DFP at 50, 100, and 200 μM for 24 h and then stained with a DNA-binding dye, DAPI, morphological changes were microscopically examined. Cells without DFP treatment showed circular shapes with healthy appearances and clear outlines, which are normal cell growth phenomena. However, after 24 h of DFP (100 and 200 μM) treatment, cells appeared with plasma membrane blebbings and an apoptotic body (data not shown). The nuclei of cells exhibited typical morphological features of apoptosis such as chromatin condensation and nuclei fragmentation (Fig. 3A). Furthermore, increasing concentrations of DFP induced a progressive accumulation of fragmented DNA, which appeared as a typical ladder pattern of DNA fragmentation due to internucleosomal cleavage associated with apoptosis (Fig. 3B). These data strongly suggest that U937 cells undergo apoptosis after exposure to DFP. The inhibited cell growth and the induction of apoptosis in U937 cells by DFP may be related to its strong antioxidant activity and anti-inflammatory effects. 14,15 Previous studies reported that HCADs including ferulic and caffeic acids had considerable apoptotic effects on breast cancer cells 19 and on U937 cells, 20 and curcumin, a diketone form of ferulic acid, induced apoptosis of leukemia cancer cells. 27 However, few reports on the apoptotic effects of plant-derived phenolic amides are available. It is very interesting to note that DFP, a main phenolic amide in corn bran, induced considerable apoptosis in U937 cells.

Effects of DFP on morphological changes and DNA fragmentation in U937 cells.
Effect of DFP on expression of anti-apoptotic proteins and release of cytochrome c in U937 cells
Apoptosis is an endogenous programmed cell death that can be triggered by various stimuli, including death receptor-mediated signaling (extrinsic or cytoplasmic pathway) and intracellular stresses (intrinsic or mitochondrial pathway). It is widely accepted that alterations to mitochondrial structure and function, as well as caspase activation, play important roles in apoptosis. 2 In response to apoptotic stimuli, the outer membrane becomes permeable, leading to the release of cytochrome c and activation of second mitochondria-derived caspase. Cytochrome c, released in the cytosol, interacts with Apaf-1, leading to the activation of the caspase cascade and to subsequent apoptosis. Activated caspases lead to the cleavage of apoptotic molecules, phospholipase C gamma 1 (PLC-γ1), cleaved caspase-3 (active form of caspase-3), and poly-(ADP-ribose) polymerase (PARP). 2,28 To investigate the underlying mechanisms involved in DFP-induced apoptosis, the changes in the expression levels of various anti-apoptotic proteins were analyzed by Western blotting. As shown in Figure 4A, DFP-treated U937 cells caused cleavage of PLC-γ1 and PARP, which serves as a marker of apoptosis, and remarkably decreased protein levels of IAP family proteins such as cIAP2 and XIAP, and Bcl-2 family proteins such as Bcl-2 and Mcl-1 and procaspase-3 in a concentration-dependent manner at 50–200 μM. In addition, Western blotting analysis using cytosolic fractions was performed to examine the release of mitochondrial cytochrome c in DFP-treated U937 cells. DFP markedly induced dose-dependent release of cytochrome c into the cytoplasm (Fig. 4B). NF-κB is known to be an important survival mechanism in cancer cells. DFP slightly increased IκB expression, and inhibited NF-κB promoter activity in a dose-dependent manner (Fig. 4C). These results suggest that DFP-induced apoptosis is associated with inhibition of Bcl-2 and IAP family protein functions, and inhibition of NF-κB transcriptional activity.
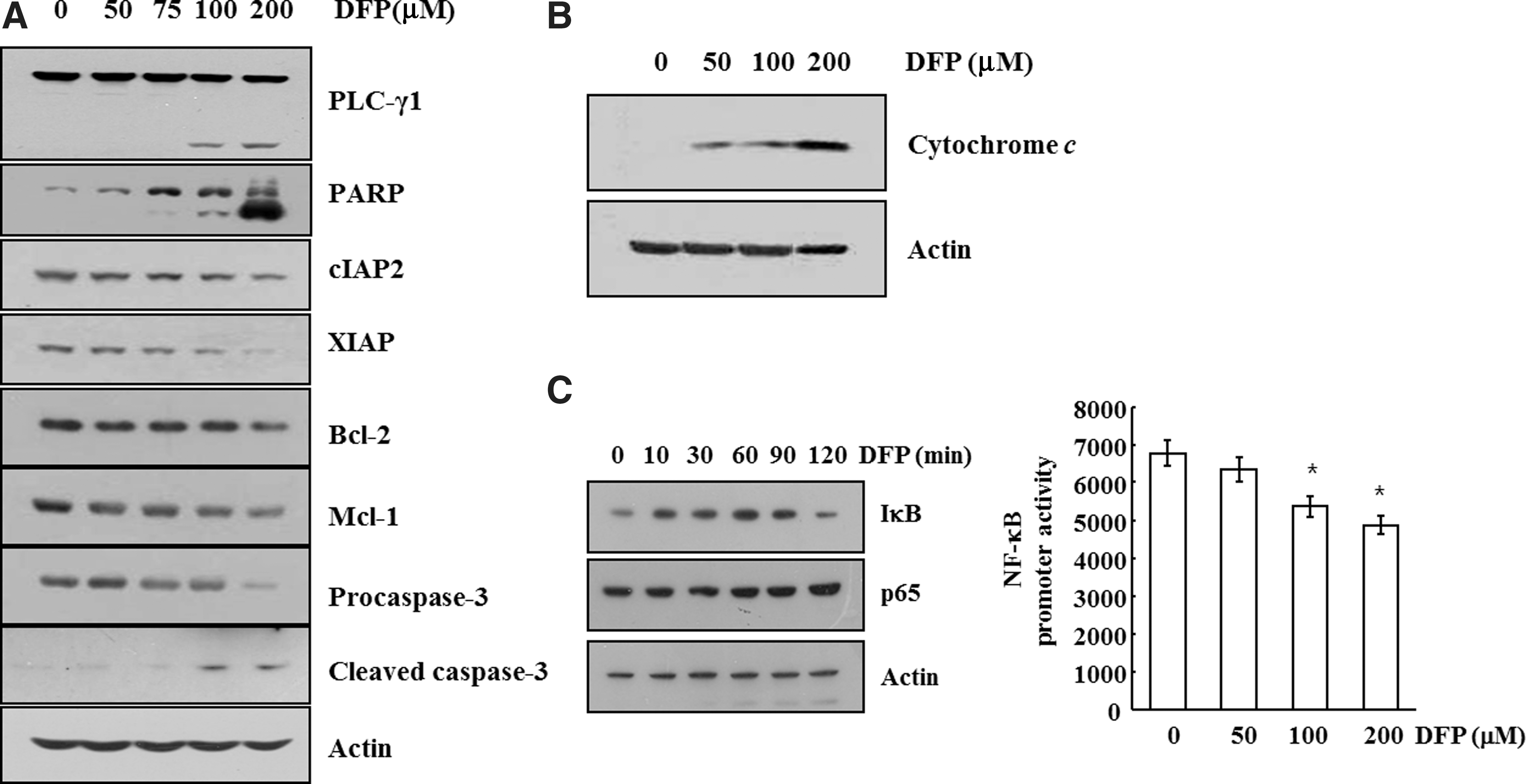
Effects of DFP on expressions of anti-apoptotic proteins, release of cytochrome c, and NF-κB promoter activities in U937 cells.
Effect of DFP on apoptosis in various cancer cells
To investigate the cell-specific actions of the DFP-induced apoptosis in leukemia cells, four tumor cell types, including renal carcinoma cells (Caki), colon cancer cells (HT29), hepatocarcinoma cells (SK-Hep1), and breast cancer cells (MDA-MB231), were treated with different concentration of the DFP at a range of 50–200 μM for 24 h and apoptosis induction determined using flow cytometry. As shown in Figure 5, DFP dose dependently induced the apoptosis against Caki and SK-Hep1 cells at a range of 50–200 μM through accumulation of sub-G1 cells, although the apoptotic effects were lower in Caki and SK-Hep1 cells than U937 cells. In addition, DFP induced apoptosis in HT29 cell only at a high concentration of 200 μM, and little apoptosis against MDA-MB231 cells. DFP did not exhibit any noticeable cytotoxicity against normal human skin fibroblast and mesangial cells (Fig. 6). These observations indicated that DFP selectively induced apoptotic cell death in leukemia cell with low cytotoxicity against the normal cells.
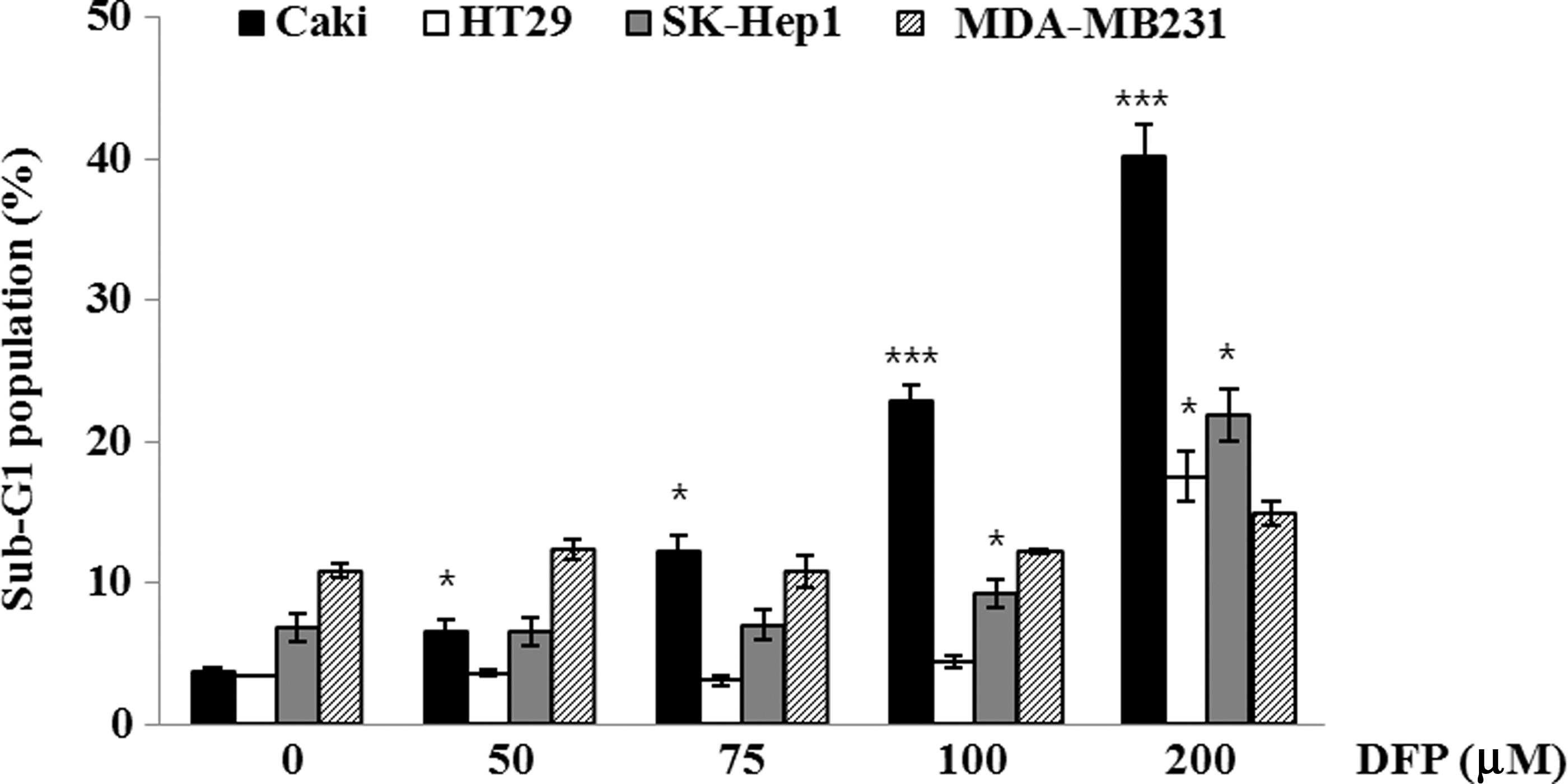
Effect of DFP on apoptosis in various cancer cells. Caki, HT29, SK-Hep1, and MDA-MB231 cells were treated with the indicated concentrations of DFP for 24 h. Apoptosis was analyzed as the sub-G1 fraction by FACS. Data represent the mean±SD of three independent experiments. *P<.05, ***P<.001 compared to control.
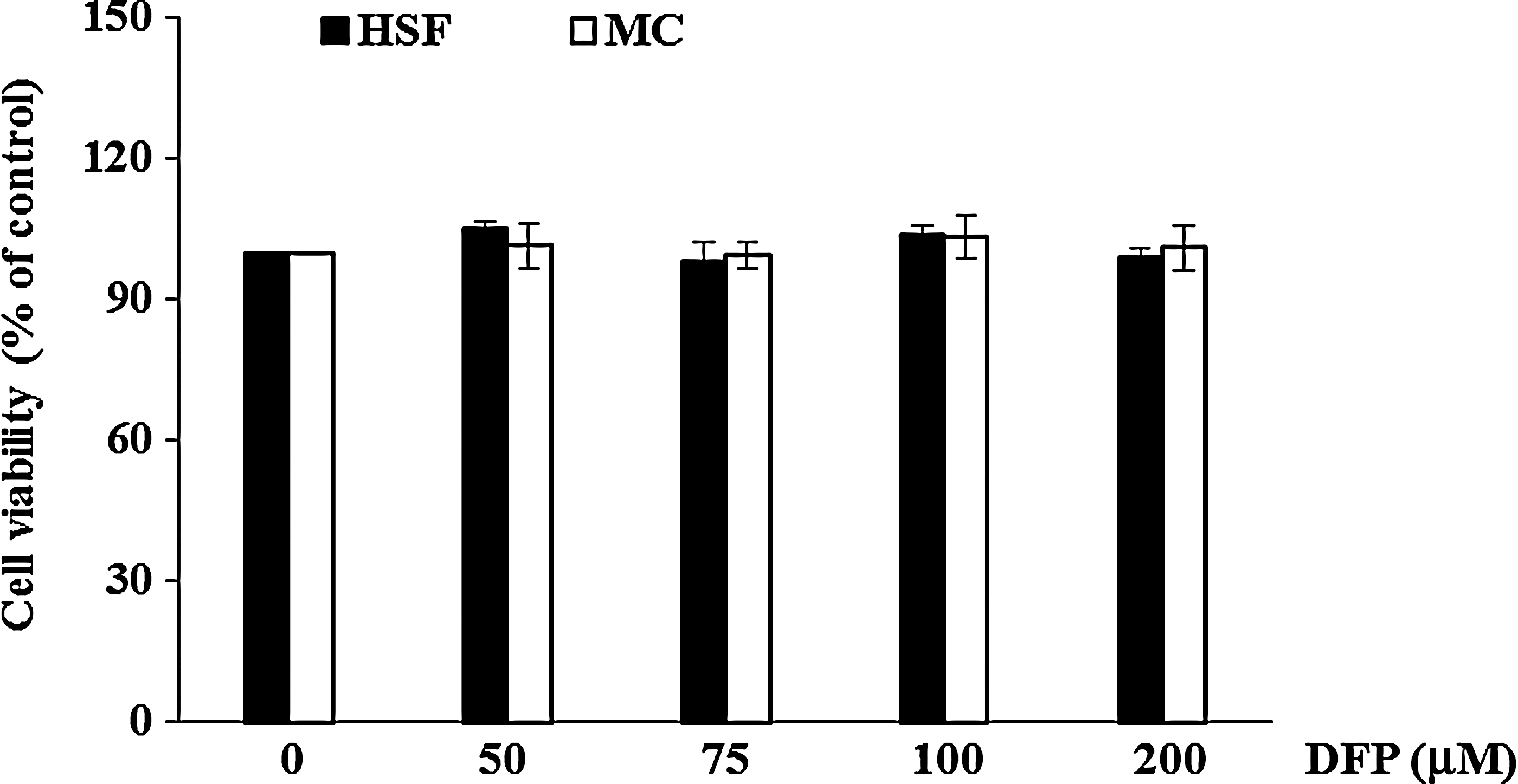
Cytotoxic effect of DFP on the growth of normal cells. Effect of DFP on viability of human skin fibroblast (HSF) and mesangial cells (MC). The cells were seeded in 96-well plates and cultured in triplicate in the presence or absence of various concentrations of DFP (50–200 μM) for 24 h, and the viability of the cells was determined by XTT assay. Cell survival is shown as a percentage of untreated control cells are the mean±SD from three independent experiments.
To date, FA, the parent moiety of DFP, have been reported to possess anticancer effect by apoptosis in T47D human breast cancer cells and HepG2 human hepatoma cells. 19,29 Park 30 recently reported that the becatamide of synthesized phenolic amides significantly inhibited H2O2-induced apoptosis of PC-12 cells via protecting mitochondrial membrane integrity. However, anticancer effects of naturally occurring phenolic amides have not been subjected to much investigation, probably due to the scarcity of the amides. This is the first report of anticancer activity of DFP, a major phenolic amide isolated from corn bran, and its possible apoptotic mechanism at simple molecule levels.
In conclusion, DFP, a predominant phenolic amide in corn bran, exerted potent cytotoxic and proapoptotic activities through suppressed gene expression of Bcl-2 and IAP family and inhibited transcriptional activity of NF-κB.
DFP selectively induced apoptosis in U937 leukemia cells without cytotoxicity against the normal cells. Further studies are required to investigate detailed mechanisms of the apoptosis-inducing activity of DFP in cancer cells and to ascertain if DFP exerts anticarcinogenic activity in vivo.
Footnotes
Acknowledgment
This research was supported by a research grant by Catholic University of Daegu, Hayang, Gyeongbuk, Korea.
Author Disclosure Statement
No competing financial interests exist.