
Abstract
Inflammatory diseases remain the leading cause of mortality worldwide in both men and women. Schizonepeta tenuifolia (ST) exerts a wide range of physiological activities and has been found to possess beneficial efficacies against inflammation-related diseases; however, the molecular mechanisms underlying this anti-inflammatory activity remain to be elucidated. We investigated the molecular basis for the downregulation of toll-like receptor 4 (TLR4) signal transduction by ST ethanol extract in lipopolysaccharide (LPS)-stimulated macrophages. In this study, ST ethanol extract (100 μg/mL) did not induce cell cytotoxicity and was used in all the following experiments. Treatment of LPS-stimulated macrophages with ST ethanol extract resulted in a significant decrease in cyclooxygenase-2 and prostaglandin E2 levels, and inducible nitric oxide synthase-mediated NO production. LPS-induced expression of cell surface molecules (CD80 and CD86) and production of pro-inflammatory cytokines (tumor necrosis factor-α, interleukin [IL]-1β, and IL-6) were inhibited by ST ethanol extract. Further, we also found that the anti-inflammatory activities of ST ethanol extract was caused by inhibition of LPS-induced activation of mitogen-activated protein kinases, such as extracellular signal-regulated kinase 1/2 and p38, and the translocation of nuclear factor κB through TLR4 in macrophages. Thus, ST ethanol extract may possess novel and potent therapeutic efficacy for the treatment of inflammatory disease.
Introduction
I
Materials and Methods
Materials
LPS isolated from Escherichia coli O111:B4 was purchased from Sigma (San Diego, CA, USA). Inducible nitric oxide synthase (iNOS) polyclonal (p) antibody (Ab), COX-1/2 pAb, anti-phosphorylated ERK1/2 monoclonal (m) Ab, anti-ERK1/2 pAb, anti-phosphorylated p38 mAb, anti-p38 pAb, and inhibitory (I)κB-α pAb were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Horseradish peroxidase (HRP)-conjugated anti-mouse immunoglobulin G (IgG) Ab and HRP-conjugated anti-rabbit IgG Ab were obtained from Calbiochem (San Diego, CA, USA). FITC-conjugated mAb against CD11b, and phycoerythrin (PE)-conjugated mAb against CD80, CD86, and cytokine detection kits for IL-1β, IL-6, and TNF-α were all purchased from eBioscience (San Diego, CA, USA).
Preparation of ST ethanol extract
A sample of ST was obtained from an oriental drug store, Tojongherb Ltd. (Seoul, Republic of Korea). ST was extracted by 70% ethanol (120 mg/mL) and carried out at room temperature with continuous agitation for 24 h. The extract was collected by centrifugation (at 3200 g for 5 min) and filtered through a 0.45 μm syringe filter. It was dried using a rotary evaporator and stored below −20°C. The extract was diluted to the required concentrations on the day of use.
Generation of BMDMs
BMDMs were isolated from C57BL/6 mice by using standard methods. 19 Briefly, bone marrow cells from the femur and tibia were cultured in DMEM containing 2 mM L-glutamine, 100 U/mL penicillin, 100 U/mL streptomycin, 10% fetal bovine serum (FBS), and 25 ng/mL recombinant mouse macrophage colony-stimulating factor (R&D Systems, Minneapolis, MN, USA) and incubated at 37°C in 5% CO2. After 4 days, nonadherent cells were removed and allowed to differentiate.
Cell proliferation
Cell proliferation was examined using the EZ-Cytox cell viability kit (Daeil Laboratories, Seoul, Republic of Korea) according to the manufacturer's instructions. BMDMs were seeded in 96-well plates at a density of 2×104 cells/well in DMEM. After incubation at 37°C for 4 h, the medium was replaced with DMEM containing ST for 24 h. EZ-Cytox kit reagent (10 μL) was added to each well, and the cells were incubated for 1 h. The optical density was measured at 450 nm in an automated micro-plate reader (Zenyth 3100; Anthos Labtec Instruments GmbH, Salzburg, Austria).
Nitric oxide production
The concentration of nitric oxide (NO) in culture supernatant was determined by measuring its oxidation product, nitrite. In this experiment, the Griess method was employed to detect NO, which is based on the chemical diazotization reaction. Briefly, supernatants from experimental BMDM cultures were mixed with the Griess reagent (1:1) and incubated at room temperature for 15 min, and then, the absorbance of the solution at 517 nm was measured using a microplate reader (Zenyth 3100). NaNO2 freshly prepared in deionized water was used to generate a standard curve (0–100 μM) to calculate the nitrite concentration in cell culture supernatants.
Elisa
Supernatants from experimental BMDM cultures were collected and stored at −70°C until use. The levels of IL-1β, IL-6, and TNF-α in the supernatants were determined using cytokine detection ELISA kits (BD Biosciences, San Jose, CA, USA) according to the manufacturer's instructions, with detection at 450 nm using a microplate reader. The concentration of prostaglandin E2 (PGE2) in cell culture supernatants was determined by using a PGE2 detection ELISA kit (Cayman Chemicals, Ann Arbor, MI, USA) following the manufacturer's instructions.
Measurement of cell surface molecules by flow cytometry
Four days after differentiation, BMDMs were harvested, washed with PBS, and resuspended in washing buffer (2% FBS and 0.1% sodium azide in PBS) for fluorescence-activated cell sorting (FACS) on the FACSCalibur flow cytometer (BD, Franklin Lakes, NJ, USA). The cells were preincubated with 0.5% BSA in PBS for 30 min and washed with PBS. The cells were then stained with PE-conjugated anti-CD80 and anti-CD86, along with FITC-conjugated anti-CD11b (BD PharMingen, San Diego, CA, USA) for 45 min at 4°C. All antibodies were diluted 100-fold before use. Cells were washed thrice and resuspended in a final volume of 500 μL of PBS. The markers of BMDM maturation were then analyzed by flow cytometry, and the data were analyzed using the Cell Quest software (BD).
Immunoblotting analysis
BMDMs were lysed in a lysis buffer (Cell Signaling, Danvers, MA, USA) containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Triton-X100, 1 mM EDTA, 50 mM NaF, and 30 mM Na4PO7, phenylmethylsulfonyl fluoride (Sigma), and lysates were separated by 10% SDS-PAGE and transferred onto PVDF membranes. The PVDF membranes were blocked with 5% skim milk and incubated with each of the required primary Abs (1:1000) for 2 h, followed by incubation with HRP-conjugated secondary Abs (1:2000) for 1 h at room temperature. Proteins were visualized using an ECL Advance kit (GE Healthcare, Little Chalfont, United Kingdom).
Nuclear extract preparation
Nuclear extracts from cells were prepared as follows. Cells were treated with 100 μL lysis buffer (10 mM HEPES [pH 7.9], 10 mM KCl, 0.1 mM EDTA, 0.5% Nonidet P-40, 1 mM dithiothreitol [DTT], and 0.5 mM PMSF) on ice for 10 min. Following centrifugation at 13,000 g for 5 min, the pellet was resuspended in 100 μL extraction buffer (20 mM HEPES [pH 7.9], 400 mM NaCl, 1 mM EDTA, 1 mM DTT, and 1 mM PMSF) and incubated on ice for 30 min. After centrifugation at 39,000 g for 10 min, the supernatant containing nuclear extracts was collected and stored at −80°C until required.
Statistical analysis
All experiments were repeated at least 3 times with consistent results. Mean and standard deviations for results were calculated using the SPSS software version 10.0 (2000). Differences among the mean values were examined by the Student's two-tailed t-test. Results are expressed as the mean±SD. Values of P<.05, P<.01, and P<.001 were considered statistically significant.
Results
Proliferative effect of ST ethanol extract
To determine a safe concentration of ST ethanol extract for use in all the following experiments, the effect of ST ethanol extract on cell viability of BMDMs was investigated, primarily by Ez-Cytox cell viability kit. ST ethanol extract was administered in a sequential and cumulative manner (12.5–100 μg/mL) to the cultured BMDMs. The administration of ST ethanol extract to BMDMs did not induce cell cytotoxicity compared to the control group (Fig. 1). For this reason, ST ethanol extract at a concentration of 100 μg/mL or less was used in all of the subsequent experiments.
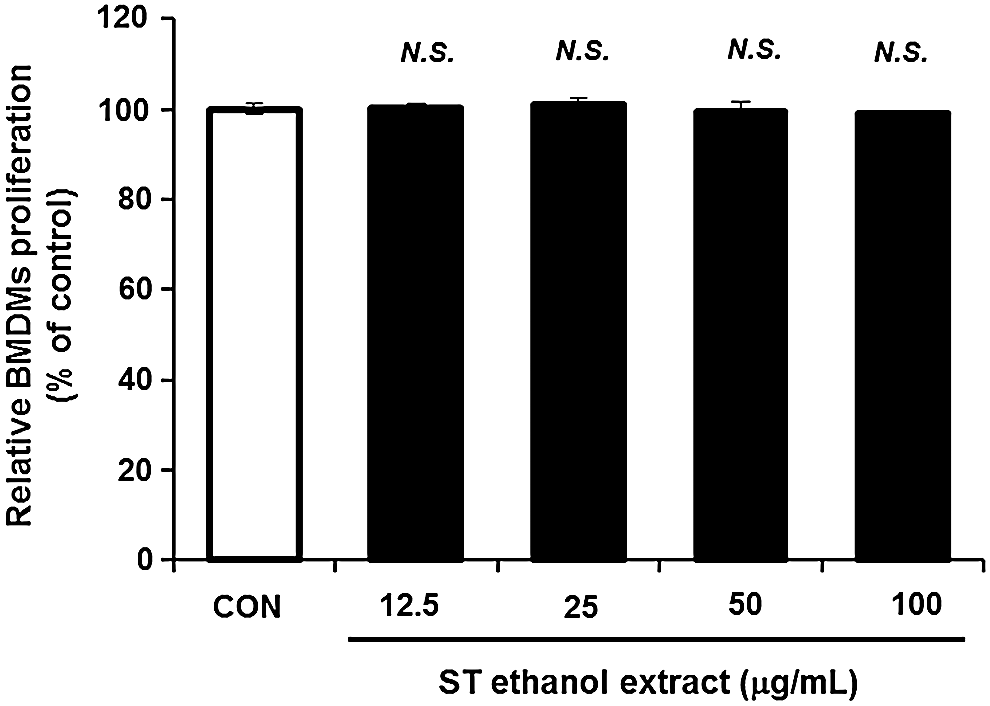
Effect of Schizonepeta tenuifolia (ST) ethanol extract on proliferation of bone marrow-derived macrophages (BMDMs). Cells were treated with ST ethanol extract (from 12.5–100 μg/mL) for 24 h, and the cell viability was assessed by Ez-Cytox cell viability kit. The results are expressed as mean±SD (n=6). Significant differences between control and ST ethanol extract were evaluated by unpaired Student's t-test. N.S., no significance.
Effect of ST ethanol extract on the expression of LPS signaling mediators
NO and PGE2 and iNOS or COX-2 are major mediators involved in systemic inflammation, and these mediators are readily induced in response to LPS. 20 As shown in Figure 2, Treatment of BMDMs with LPS induced the production of high levels of iNOS-mediated NO, COX-2, and PGE2, which were significantly inhibited by treatment with ST ethanol extract. Taken together, these data show that ST ethanol extract is capable of regulating LPS-induced macrophage activation.

Effect of ST ethanol extract on the expression of LPS signaling mediators. Cells were incubated with ST ethanol extract (25–100 μg/mL) for 1 h and then treated with LPS (100 ng/mL) for 24 h.
Effect of ST ethanol extract on LPS-induced pro-inflammatory cytokines
Based on the results obtained above, we next measured whether ST ethanol extract could influence the modulation of LPS-stimulated pro-inflammatory cytokines in BMDMs. As shown in Figure 3, treatment with ST ethanol extract (100 μg/mL) significantly decreased LPS-induced production of IL-1β, IL-6, and TNF-α compared to that in BMDMs treated with LPS alone. These results strongly suggest that ST ethanol extract is able to modulate inflammation by suppressing the production of pro-inflammatory cytokines.
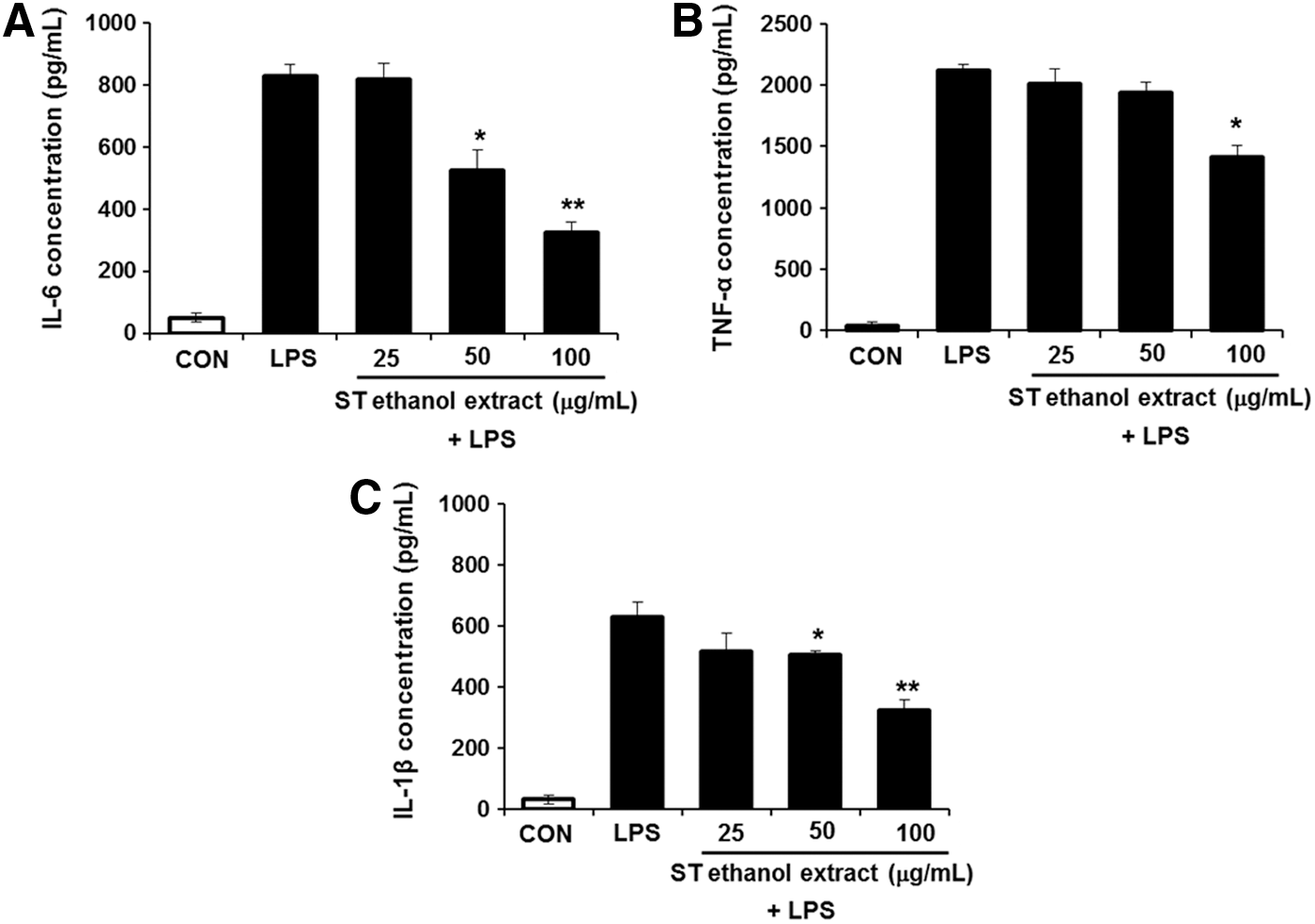
Effect of ST ethanol extract on pro-inflammatory cytokine release in LPS-induced bone marrow-derived macrophages (BMDMs). Cells were pretreated with ST ethanol extract (25–100 μg/mL) for 1 h, and then stimulated with LPS (100 ng/mL) for 24 h. The amount of
Effect of ST ethanol extract on LPS-induced CD80 and CD86 expression
We subsequently examined whether ST ethanol extract had a modulatory role in the LPS-induced expression of macrophage surface molecules, such as CD80 and CD86. BMDMs upregulated CD80 and CD86 protein expression in response to LPS, and the expression of these molecules was significantly lower in BMDMs treated with ST ethanol extract (50 or 100 μg/mL) compared with treatment with LPS alone (Fig. 4).
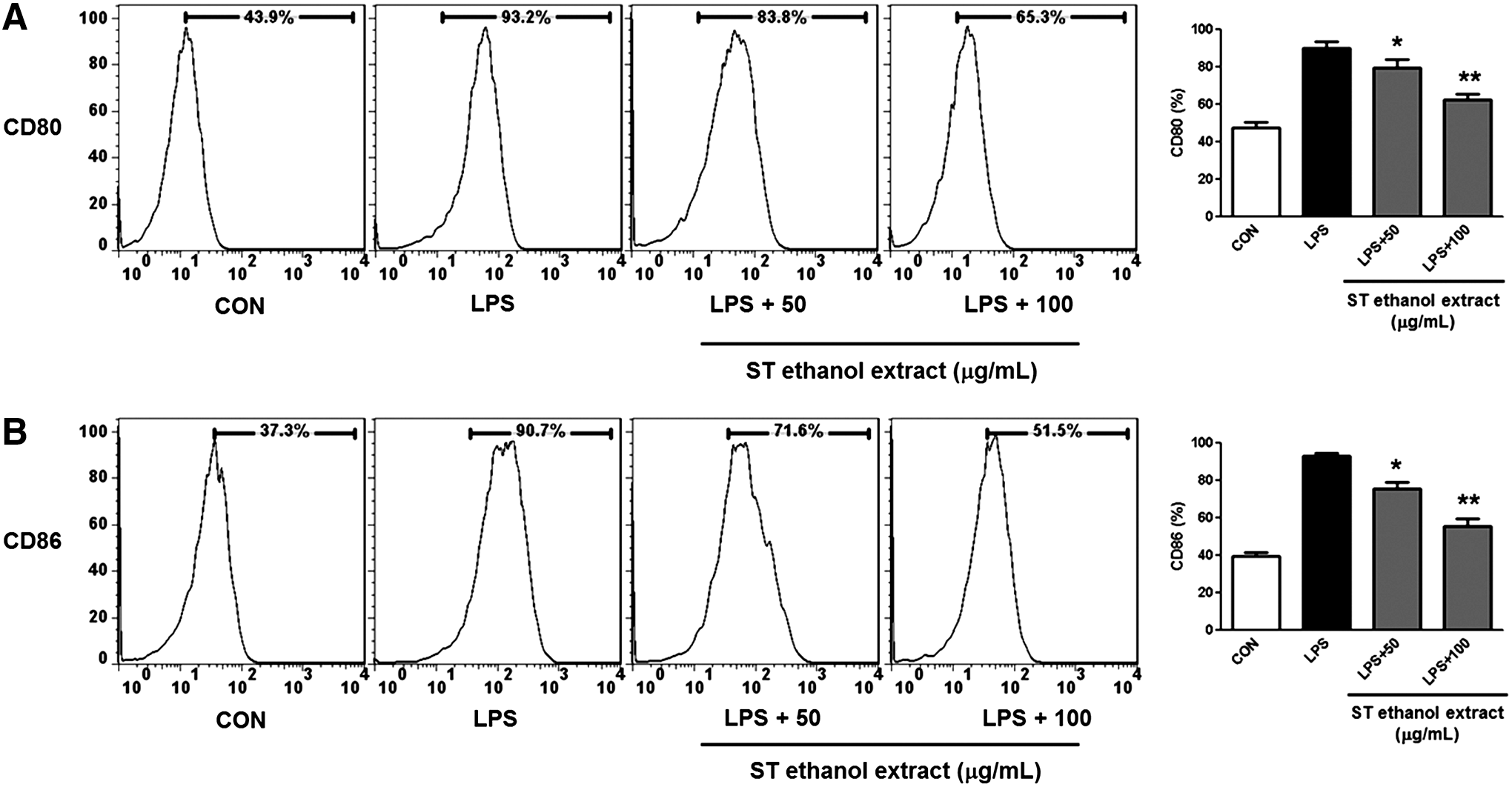
Effect of ST ethanol extract on
Effect of ST ethanol extract on LPS-induced NF-κB and MAPK signaling through TLR4
TLR4 is activated by LPS and is required for LPS-induced NF-κB and AP-1 activation and chemokine expression. 8,9 To investigate whether ST ethanol extract suppresses the expression of TLR4, cells were treated with 50 and 100 μg/mL ST ethanol extract. Protein levels of TLR4 were not affected under a 1-h ST ethanol extract treatment (Fig. 5A). On the other hand, when the cells were incubated with ST ethanol extract for 6–24 h, protein levels of TLR4 was markedly lower in a time- or dose-dependent manner (Fig. 5A). Next, we examined whether ST ethanol extract suppresses LPS-induced MAPK and NF-κB activation through TLR4, because the activation of MAPK and NF-κB signaling through TLR4 is a critical step in the induction of many genes involved in inflammation. As shown in Figure 5B, LPS-induced phosphorylation of ERK1/2 and p38 was inhibited by treatment with ST ethanol extract in a concentration-dependent manner. ST ethanol extract significantly suppressed LPS-induced phosphorylation of IκB-α in a concentration-dependent manner (Fig. 5C). Further, we investigated the effect of ST ethanol extract on nuclear translocation of p65 induced by LPS. We also found that ST ethanol extract significantly suppressed LPS-induced nuclear translocation of p65 in control cells (Fig. 5C). Ultimately, the results of the present study showed that ST ethanol extract suppressed MAPK and NF-κB-mediated signaling through TLR4 in LPS-stimulated macrophages.
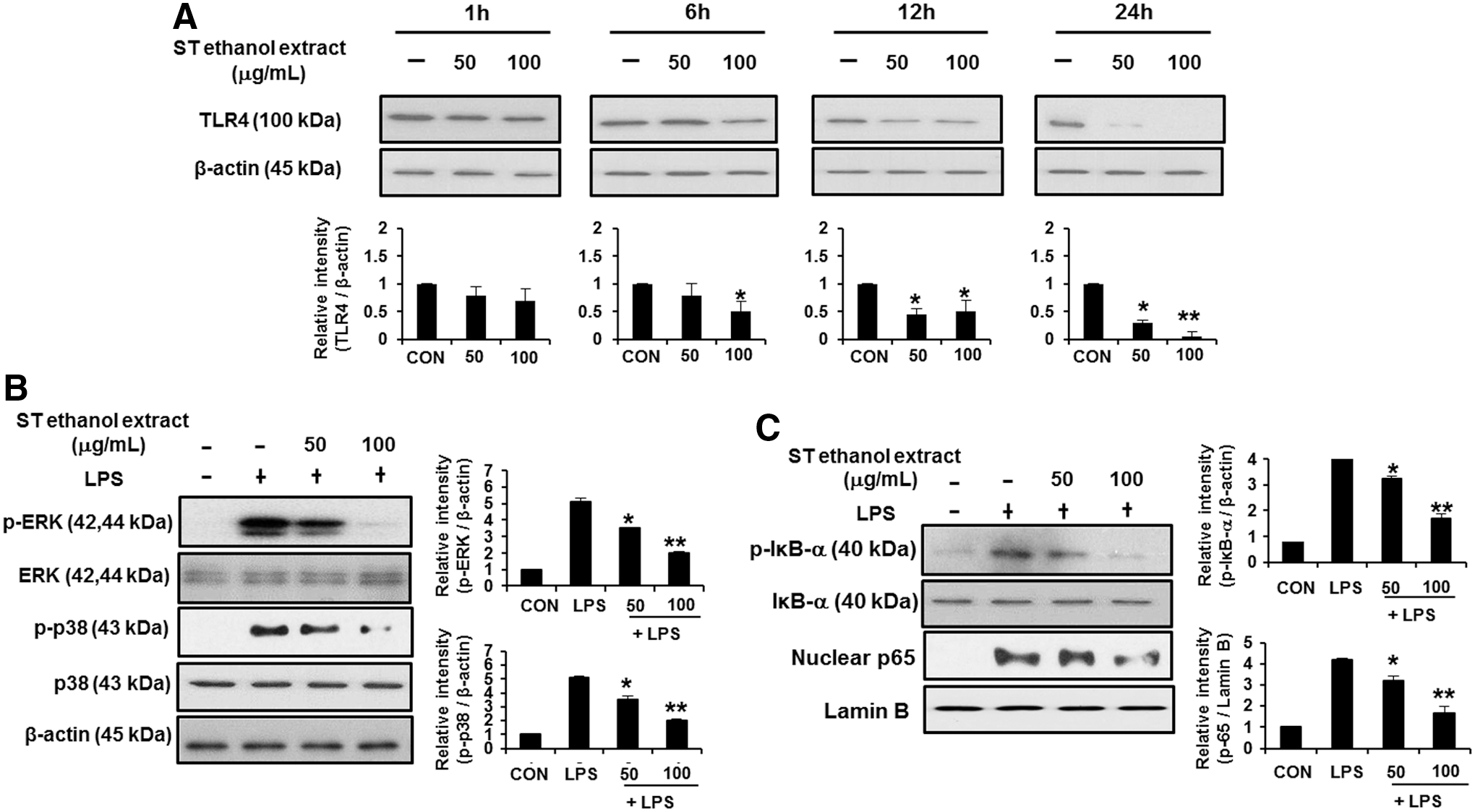
Effect of ST ethanol extract on LPS-induced NF-κB and MAPK signaling through TLR4.
Discussion
Acute or chronic inflammation is an important public health problem, particularly in developed countries, and epidemiologic reports have indicated that natural compounds used in traditional medicine may be associated with decreased incidence of various diseases, especially inflammation. 21 –24 Natural compounds from various herbs are currently attracting intense interest, because they have many beneficial effects without the side effects of pharmaceutical treatments. 12,21,25 As demonstrated by other reports, ST is known to be useful for treating cold, allergy, and inflammatory disease. 17,26,27 However, it has not yet been established how ST regulates the inflammatory responses. The present study was undertaken to elucidate the potential of ST ethanol extract as a novel anti-inflammatory herb, and found that ST ethanol extract exerts anti-inflammatory effects through the inhibition of TLR4 signaling, leading to downregulation of various inflammatory mediators in LPS-stimulated macrophage cells.
Primarily, to determine the moderate concentration of ST ethanol extract used in all the following experiments, the effect of ST ethanol extract on proliferation of BMDMs was investigated in a concentration-dependent manner. The administration of ST ethanol extract (≤100 μg/mL) to BMDMs did not induce cell cytotoxicity and was used in all the following experiments. Here, we next showed that ST ethanol extract resulted in a dose-dependent decrease of NO, iNOS, PGE2, and COX-2 expression in LPS-stimulated BMDMs. The present study also showed that the extract decreased the production of LPS-stimulated pro-inflammatory cytokines, such as IL-1β, IL-6, and TNF-α. These results confirm that the production of NO, PGE2, IL-1β, and TNF-α is an important part of immune response; however, excessive production of these mediators ultimately induce chronic inflammation. 28 –30 In the NOS family, iNOS is particularly involved in pathological overproduction of NO, which is known to play a central role in inflammation. 28 Additionally, previous studied reported that the key signaling pathway of inflammation induced by LPS in macrophages involves NO production through NF-κB and MAPK-mediated signaling, and suggested that inhibition of the NF-κB and MAPK pathways are two major mechanisms underlying the attenuation of LPS-induced inflammatory cytokine production. 28,29,31 Consistent with the above observaitons, it has been reported that several natural compounds directly inhibit the expression of NF-κB and MAPK-dependent cytokines, iNOS, PGE2, and COX-2 genes, 32 –35 which is consistent with our results that ST ethanol extract suppressed LPS-induced MAPK and NF-κB activation. In additional studies, we next examined whether ST ethanol extract suppresses LPS-induced MAPK and NF-κB activation through TLR4, because the activation of MAPK and NF-κB signaling through TLR4 is a critical step in the induction of many genes involved in inflammation. The present data showed that ST ethanol extract suppressed LPS-stimulated TLR4 signaling, leading to NF-κB and MAPK-mediated signaling. As demonstrated by previous studies, the activation of LPS-stimulated TLR4 ultimately leads to MAPKs and NF-κB-mediated production of pro-inflammatory cytokines, such as IL-6 and TNF-α, and surface molecule expression, such as CD80 and CD86, in macrophages. 8,36,37
Taken together, present findings suggest that ST ethanol extract has potent anti-inflammatory effects through the inhibition of TLR4, leading to MAPK and NF-κB-mediated signaling in LPS-stimulated macrophages (Fig. 6). Our results provide a new insight into the pharmacological role of ethanolic extract of ST against the development and progression of chronic inflammation, and we recommend further in vitro and in vivo studies to completely elucidate the mechanism(s) that underlie the anti-inflammatory activity of this natural compound.
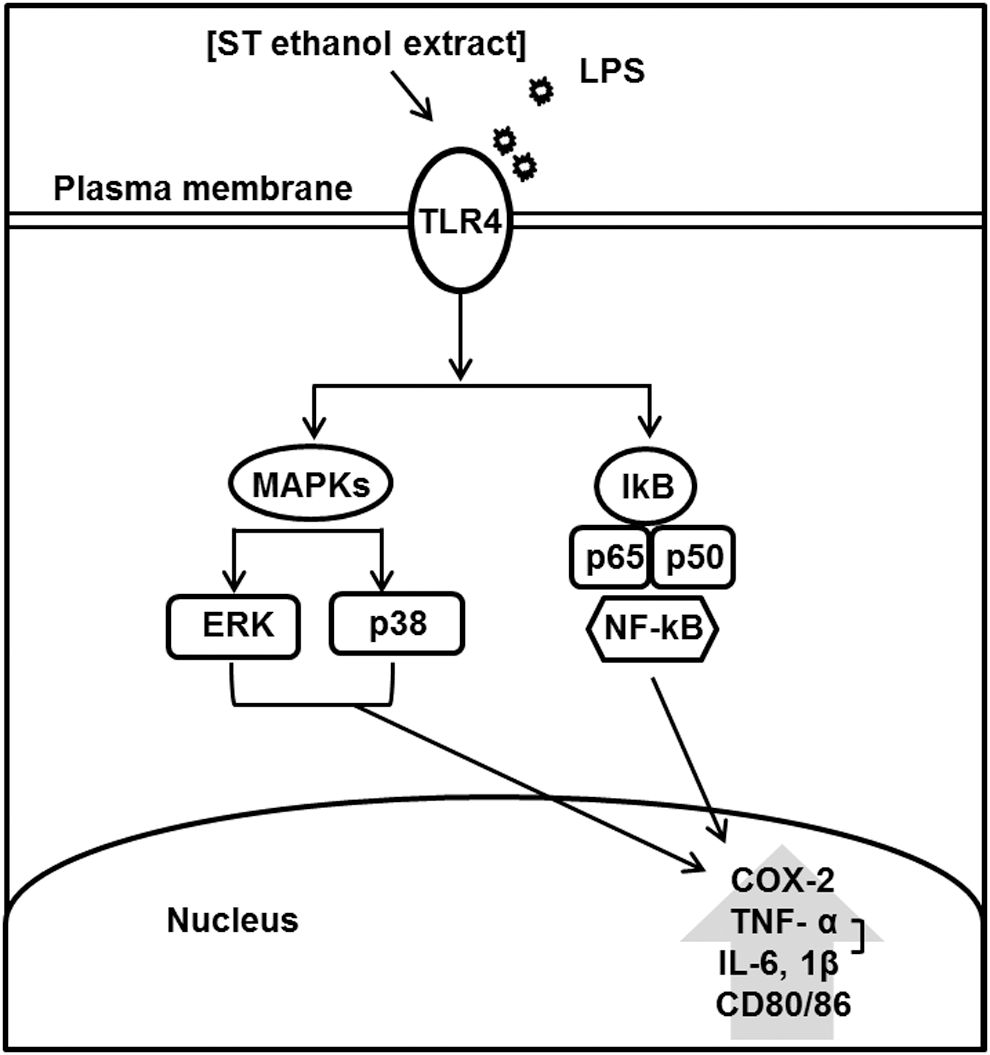
Proposed mechanism of ST ethanol extract-induced anti-inflammation in bone marrow-derived macrophages.
Footnotes
Acknowledgment
This study was supported by Woosong University in 2013. The author thanks Ms. Yang Mi-So (Chungnam National University) for her helpful support in the cell experiments.
Author Disclosure Statement
The author declares that there are no conflicts of interest.