
Abstract
Aging is a progressive process, and it may lead to the initiation of neurological diseases. In this study, we investigated the effects of wild Indian Curcuma longa using a Morris water maze paradigm on learning and spatial memory in adult and D-galactose-induced aged mice. In addition, the effects on cell proliferation and neuroblast differentiation were assessed by immunohistochemistry for Ki67 and doublecortin (DCX) respectively. The aging model in mice was induced through the subcutaneous administration of D-galactose (100 mg/kg) for 10 weeks. C. longa (300 mg/kg) or its vehicle (physiological saline) was administered orally to adult and D-galactose-treated mice for the last three weeks before sacrifice. The administration of C. longa significantly shortened the escape latency in both adult and D-galactose-induced aged mice and significantly ameliorated D-galactose-induced reduction of cell proliferation and neuroblast differentiation in the subgranular zone of hippocampal dentate gyrus. In addition, the administration of C. longa significantly increased the levels of phosphorylated CREB and brain-derived neurotrophic factor in the subgranular zone of dentate gyrus. These results indicate that C. longa mitigates D-galactose-induced cognitive impairment, associated with decreased cell proliferation and neuroblast differentiation, by activating CREB signaling in the hippocampal dentate gyrus.
Introduction
A
There have been numerous attempts to overcome age-related memory impairment and to reduce the ROS in the brain. Some medical plants have the potential to attenuate memory deficits in artificial and natural aging models. 6 –9 One of these medicinal plants, Curcuma longa, has well-established neuroprotective effects in the brain against ischemic damage, 10 –12 Alzheimer's disease, 13 –15 Parkinson's disease, 16,17 and parathion-induced damage. 18 C. longa extract contains a polyphenolic nonflavonon compound—curcumin—which is a pharmacologically active substance. 19 Although curcumin has poor bioavailability, 20,21 it has been reported that it is a very effective agent in brain disease models such as amygdaloid kindled seizures in rats, 22 an iron-induced experimental model of epileptogenesis, 23 and electroshock seizures in mice 24 because it can cross the blood–brain barrier to enter brain tissue in spite of its poor bioavailability. Curcumin has been concentrated chiefly in the hippocampus in the kinetics of tissue distribution. 25 It has been reported that oral administration of curcumin improves learning and memory in aged rats 26,27 and mice, 28 and that it is protective against chronic unpredictable stress-induced cognitive impairment and associated oxidative damage in mice. 29
The hippocampus region of the brains plays an important role in memory formation and spatial navigation. There is increasing evidence supporting the view that adult hippocampal neurogenesis is closely related to pattern separation and spatial memory. 30,31 Progenitor cells located in the subgranular zone of the hippocampal dentate gyrus divide, proliferate, differentiate, and give rise to new neurons. 32 Although several studies have found an overall age-related decline in cell proliferation, 33 –39 there are few reports identifying factors that promote neurogenesis in the dentate gyrus of aged mice. Improvement in the microenvironment of neural stem cells in the subgranular zone of dentate gyrus, including ROS elimination and neurotrophic factor restoration, may be a promising strategy for enhancing successful aging. In this study, we examined the effects of C. longa using a Morris water maze paradigm on learning and spatial memory, as well as cell proliferation and neuroblast differentiation, in adult and D-gal-induced aged mice.
Materials and Methods
Preparation of wild Indian C. longa
Rhizomes of C. longa were provided by Natural Pond Co., Ltd. (Seoul, South Korea). They were cleaned, dried, chopped, and blended using a Waring blender (1-L laboratory blender, 8010S; Waring Laboratory Science, Torrington, CT). The blended powder was left to soak in 95% ethanol at a ratio of 1:10 of plant to ethanol for three days at room temperature with shaking. The mixture was then filtered, and the resulting liquid was concentrated under a vacuum at 45°C in an EYELA rotary evaporator (N-1001S-W; EYELA USA, Bohemia, NY, USA) to yield a dark gummy-yellow extract (7.4%, w/w). The concentrated extract was then kept in an incubator at 45°C for 5 days to evaporate the ethanol residue, yielding the crude rhizome extract. Extracts were kept at room temperature with silica gel until used. Extracts were dissolved in 10% Tween-20 before being orally administrated to animals, and the insoluble materials were removed through centrifugation (1000 g) at room temperature.
Experimental animals
Male C57BL/6 mice were purchased from Japan SLC, Inc. (Shizuoka, Japan), and housed under in a controlled environment maintaining temperature (23°C) and humidity (60%) and a 12/12 h light/dark cycle, and free access to food and tap water. The handling and caring of the animals conformed to published guidelines (NIH Guide for the Care and Use of Laboratory Animals, NIH Publication No. 85-23, 1985, revised 1996), and were approved by the Institutional Animal Care and Use Committee (IACUC) of Seoul National University (SNU-110511-6). Every effort was made to minimize the number of animals used and the suffering caused by the procedures.
Administration of drugs and C. longa
The animals were divided into four groups (n=10 per group): vehicle (10% Tween-20), 300 mg/kg C. longa extract (C300), D-gal (100 mg/kg; Sigma, St. Louis, MO, USA), and D-gal with C300 (D-gal+C300; Fig. 1). D-gal was subcutaneously administered to six-week-old mice once a day for 10 weeks. At 13 weeks of age, vehicle or C300 was orally administered to mice once a day for three weeks. The different experimental groups ran individually in time. The dosage of C. longa was chosen because oral administration of C. longa significantly decreases lipid peroxidation and increases antioxidants activity. 40 These schedules were adopted because doublecortin (DCX; a marker for neuroblasts) is exclusively expressed in immature neurons from 1 to 28 days of cell age. 41,42
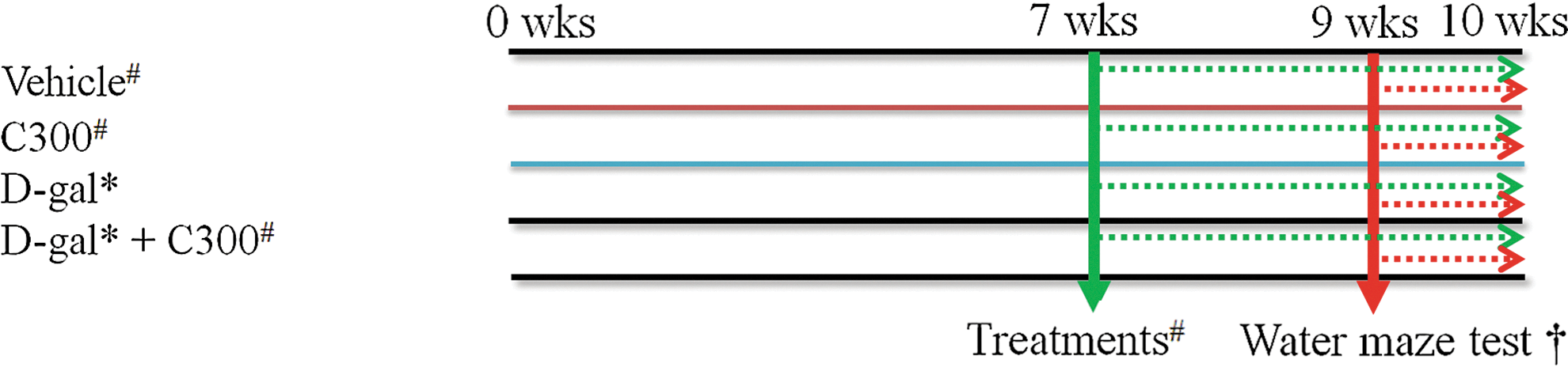
Experimental protocol in this study. *D-gal treatment (100mg/kg, S.C.) (black line) for 10 weeks; #Vehicle and C300 (300 mg/kg) treatment (green dot) for 3 weeks; †Water maze test (red dot) for 1 week. Color images available online at
Water maze performance
At the 10th week after D-gal administration, spatial memory was assayed using Morris water maze tests as previously reported. 43,44 The behavioral test and sacrifice of animals were conducted at 9:00–11:00 a.m. At three days after the training, the time required for an individual mouse to find the submerged platform within two minutes (escape latency) and the swimming distance were monitored by a digital camera and a computer system for four consecutive days and four trials per day. The administration of D-gal and/or C300 was continued during the water maze performance. For each trial, a mouse was placed in the water facing the wall at one of four starting positions and released. The swimming speed and the time required for the mouse to find the hidden platform were recorded via a visual tracking system. The probe test was done on day 5; the platform was removed and the time that a mouse spent swimming in the target quadrant and in the three nontarget quadrants (right, left, and opposite quadrants) was measured, and the number of times the mouse crossed over the platform site was recorded.
Tissue processing for histology
For histology, on the day after the water maze test, animals in the vehicle-, C300, D-gal-, and D-gal+C300-treated groups (n=5 in each group) were randomly selected and were anesthetized with 30 mg/kg Zoletil 50® (Virbac, Carros, France) at 09:00–11:00 a.m. to avoid the interference of the stress hormone. The animals were perfused transcardially with 0.1 M phosphate-buffered saline (PBS; pH 7.4) followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB; pH 7.4). The brains were removed and postfixed in the same fixative for four hours. The brain tissues were dehydrated with graded concentrations of alcohol for embedding in paraffin. Sections 3 μm thick were serially cut using a microtome (Leica, RM2245; Leica Biosystems, Newcastle upon Tyne, United Kingdom), and they were mounted onto silane-coated slides.
Immunohistochemistry
In order to obtain accurate data, immunohistochemical staining was carefully conducted under the same conditions. Tissue sections were selected between 1.96 and 2.26 mm posterior to the bregma in reference to a mouse atlas for each animal. 45 The sections were placed in 400 mL jars filled with citrate buffer (pH 6.0) and heated in a microwave oven (Optiquick Compact, Moulinex, Ecully Cedex, France) operating at a frequency of 2.45 GHz on a 800 W power setting. After three heating cycles of five minutes each, slides were allowed to cool at room temperature and were washed in PBS. They were next incubated with rabbit anti-Ki67 (a marker for cell proliferation) antibody (1:1000; Abcam, Cambridge, United Kingdom), goat anti-DCX antibody (1:50; Santa Cruz Biotechnology, Santa Cruz, CA, USA), rabbit anti-pCREB (phosphorylated CREB, diluted 1:1000, Millipore, Temecula, CA, USA), or rabbit anti-BDNF (brain-derived neurotrophic factor, 1:1000; Chemicon International, Temecula, CA, USA) overnight, and subsequently exposed to biotinylated rabbit antigoat, or goat antirabbit IgG (diluted 1:200; Vector, Burlingame, CA, USA) and streptavidin peroxidase complex (diluted 1:200, Vector). Then, the sections were visualized by reaction with 3,3′-diaminobenzidine tetrahydrochloride (Sigma).
Measurements of Ki67-, DCX-, or pCREB-immunoreactive cells in all the groups were performed using an image analysis system equipped with a computer-based CCD camera (Olympus, DP71, Tokyo, Japan; software: Optimas 6.5, CyberMetrics, Scottsdale, AZ, USA). The Ki67-, DCX-, or pCREB-immunoreactive cells in each section of the dentate gyrus were counted using Optimas 6.5 software. The cell counts from all of the sections of all of the mice were averaged.
Measurement of BDNF levels
To confirm the effects of C300 and/or D-gal on BDNF levels, the remaining animals in each group (n=5) were sacrificed under anesthesia (Zoletil 50®) by decapitation on the day after the water maze test (9:00–11:00 a.m.), and hippocampal dentate gyri were dissected out using the PixCell II system (Arcturus Engineering, Mountain View, CA, USA). BDNF levels were measured with a Promega BDNF Emax immunoassay kit (Madison, WI, USA). Briefly, samples were weighed and 300 μL lysis buffer was added to each sample. Samples were sonicated for 30 sec and centrifuged (16,000 g) at 4°C for 20 min. The supernatant was stored at−20°C until analysis. All samples were assayed in duplicate, and absorbance was read on an enzyme-linked immunosorbant assay (ELISA) plate reader (Bio-Tek, Winooski, VT, USA). The concentration of each sample was calculated by a computer by plotting the absorbance values on a standard curve with known concentrations generated by the assay.
Statistical analysis
The data presented represent the means of the experiments performed for each experimental investigation. The differences among the means were statistically analyzed by a two-way analysis of variance followed by a Bonferroni's post-hoc method in order to elucidate the differences (D-gal×C300). P<.05 was considered statistically significant.
Results
Effects of D-gal and/or C300 on spatial memory
In the vehicle-treated group, the mean escape latency was decreased within one day after initiation of the trials. In the C300-treated group, the mean escape latency was slightly shorter than in the vehicle-treated group throughout each day of the trial. However, no significant difference was observed in latency times between the vehicle- and C300-treated groups. In the D-gal-treated group, the escape latency was significantly longer during each day of the trial compared to that in the vehicle-treated group. In the D-gal+C300-treated group, the escape latency was significantly shorter throughout each day of the trial compared to that in the D-gal-treated group. However, it was longer during each day of the trial compared to that reported for the vehicle-treated group (Fig. 2A).
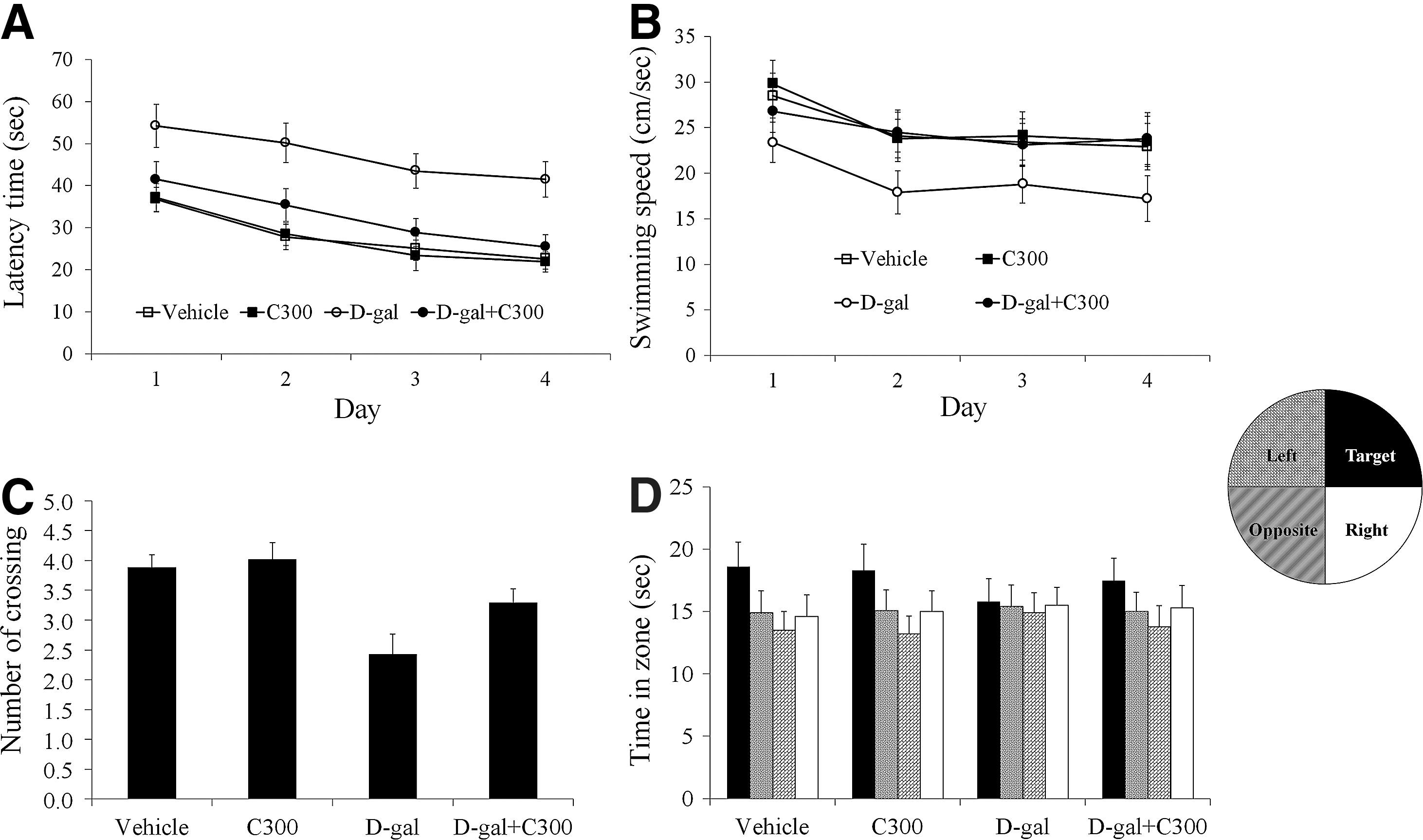
Spatial memory test using a Morris water maze in the vehicle-, C300-, D-gal-, and D-gal+C300-treated groups (n=10 per group).
The mean swimming speed in the C300-treated group was slightly faster than that in the vehicle-treated group, but in the D-gal-treated group, swimming speed was significantly slower than that in the vehicle-treated group. In the D-gal+C300-treated group, the swimming speed was significantly increased compared to that in the D-gal-treated group (Fig. 2B).
The platform crossings in the probe trial were similar between vehicle- and C300-treated groups. In the D-gal-treated group, there were significantly fewer platform crossings in the probe trial compared to the vehicle-treated group. In the D-gal+C300-treated group, the frequency of crossing over the platform site increased compared to that in the D-gal-treated group (Fig. 2C).
Spatial preference for the target quadrant was similar between vehicle- and D-gal-treated groups. Spatial preference was significantly decreased in the D-gal-treated group compared to that in the vehicle-treated group. In the D-gal+C300-treated group, spatial preference was significantly increased compared to that in the D-gal-treated group (Fig. 2D).
Effects of D-gal and/or C300 on cell proliferation
In each group, Ki67-immunoreactive nuclei were detected in the subgranular zone of the dentate gyrus. However, there were significant differences in the numbers of Ki67-immunorecactive nuclei between treatment groups. In the vehicle-treated group, the average number of Ki67-immunoreactive nuclei was 7.7 per section (Fig. 3A, E). In the C300-treated group, the mean number of Ki67-immunoreactive nuclei was significantly increased compared to the vehicle-treated group with 9.9 per section (Fig. 3B, E). In the D-gal-treated group, Ki67-immunoreactive nuclei were not clustered, and the average number of Ki67-immunoreactive nuclei was significantly decreased compared to those of the vehicle-treated group with 2.9 per section (Fig. 3C, E). In the D-gal+C300-treated group, the mean number of Ki67-immunoreactive nuclei was significantly increased compared to that in the D-gal-treated group and was 5.7 per section (Fig. 3D, E). However, the mean number of Ki67-positive nuclei in this group was less than that reported for the vehicle-treated group (Fig. 3D, E).
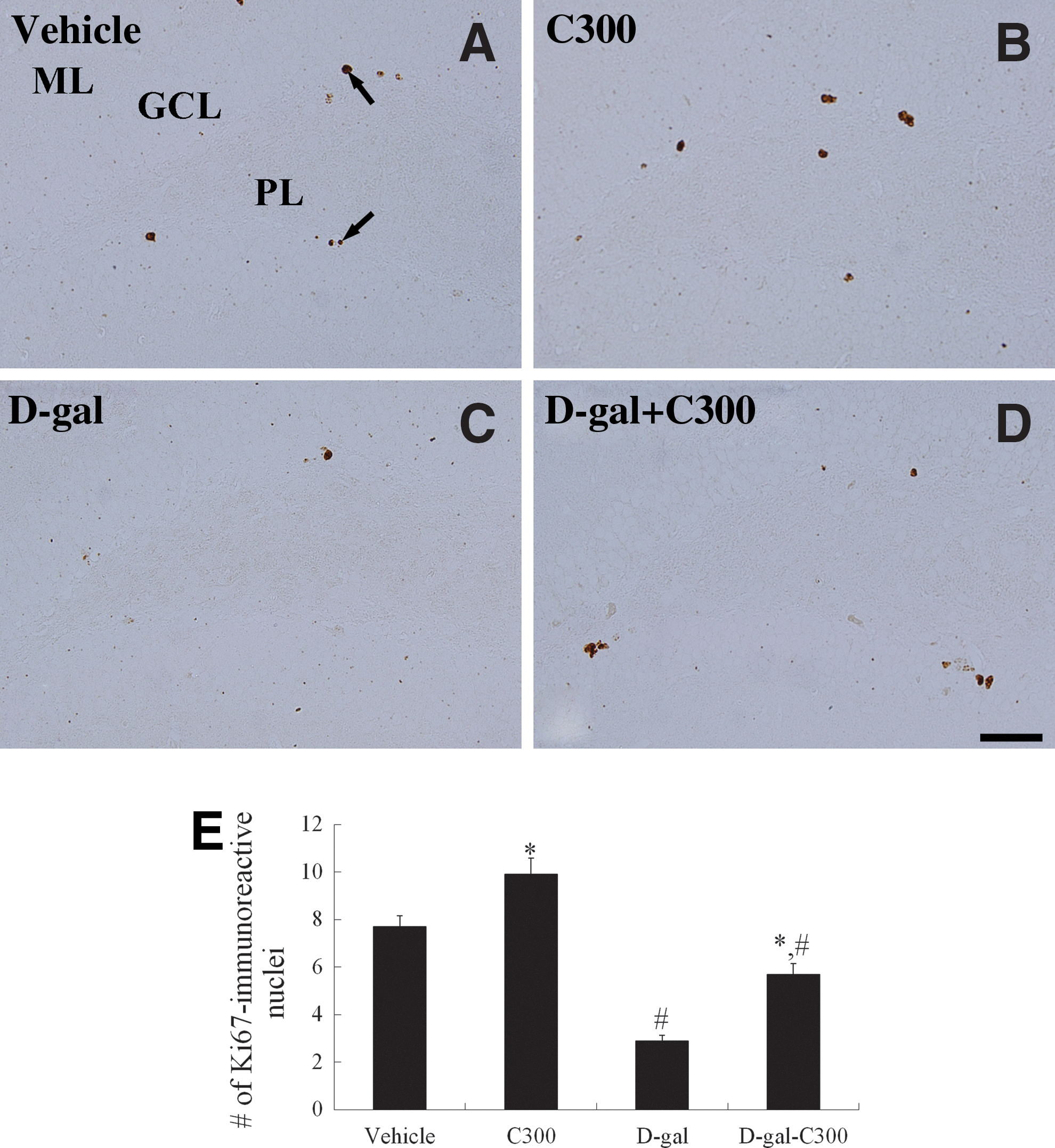
Immunohistochemistry for Ki67 in the dentate gyrus of the
Effects of D-gal and/or C300 on neuroblast differentiation
In each group, the soma of DCX-immunoreactive neuroblasts was observed in the subgranular zone of the dentate gyrus with dendrites extending into the molecular layer of dentate gyrus. However, there were significant differences in the number of DCX-immunoreactive neuroblasts with dendrites between the groups. In the vehicle-treated group, the average number of DCX-immunoreactive neuroblasts was 32.5 per section (Fig. 4A, B, I). In the C300-treated group, DCX-immunoreactive neuroblasts with dendrites were abundant compared to those in the vehicle-treated group. In this group, the average number of DCX-immunoreactive neuroblasts was 52.1 per section (Fig. 4C, D, I). In the D-gal-treated group, the average number of DCX-immunoreactive neuroblasts was significantly decreased compared to the vehicle-treated group with 12.6 per section (Fig. 4E, F, I). In the D-gal+C300-treated group, the mean number of DCX-immunoreactive neuroblasts was significantly increased compared to the D-gal-treated group with 30.1 per section (Fig. 4G–I). The mean number of DCX-immunoreactive neuroblasts was similar to that reported for the vehicle-treated group. However, there were fewer dendrites observed in the DCX-positive neuroblasts compared to the vehicle-treated group (Fig. 4I).

Immunohistochemistry for doublecortin (DCX) in the dentate gyrus of
Effects of D-gal and/or C300 on phosphorylation of CREB
In each treatment group, the pCREB-immunoreactive nuclei were mainly detected in the subgranular zone of the dentate gyrus. The average number of pCREB-immunoreactive nuclei was significantly different among the groups. In the vehicle-treated group, the average number of the pCREB-immunoreactive nuclei was 28.5 per section (Fig. 5A, E). In the C300-treated group, the average number of pCREB-immunoreactive nuclei increased compared to the vehicle-treated group with 39.5 per section (Fig. 5B, E). In the D-gal-treated group, pCREB-immunoreactive nuclei were significantly decreased compared to the vehicle-treated group. In this group, the average number of pCREB-immunoreactive nuclei quantitated was 8.5 per section (Fig. 5C, E). In the D-gal+C300-treated group, the average number of pCREB-immunoreactive nuclei was significantly increased compared to the D-gal-treated group with 26.8 per section (Fig. 5D, E).

Immunohistochemistry for phosphorylated CREB (pCREB) in the dentate gyrus of
Effects of D-gal and/or C300 on BDNF immunoreactivity and protein levels
In the vehicle-treated group, BDNF immunoreactivity was detected in a few cells in the dentate gyrus (Fig. 6A). In this group, the mean BDNF protein level in the dentate gyrus homogenates was 86.5 pg/mg (Fig. 6E). In the C300-treated group, BDNF immunoreactivity was prominently observed in the dentate gyrus (Fig. 6B), and the average BDNF protein level was reported as 128.4 pg/mg protein (Fig. 6E). In the D-gal-treated group, BDNF immunoreactivity was almost undetectable in the dentate gyrus (Fig. 6C), and the average BDNF protein level was markedly decreased (41.8 pg/mg) when compared to that in the vehicle-treated group (Fig. 6E). In the D-gal+C300-treated group, BDNF immunoreactivity was detected in the dentate gyrus (Fig. 6D), and the mean BDNF levels were significantly higher than that in the D-gal-treated group, but the BDNF protein level was significantly decreased (63.3 pg/mg) compared to that of the vehicle-treated group (Fig. 6E).
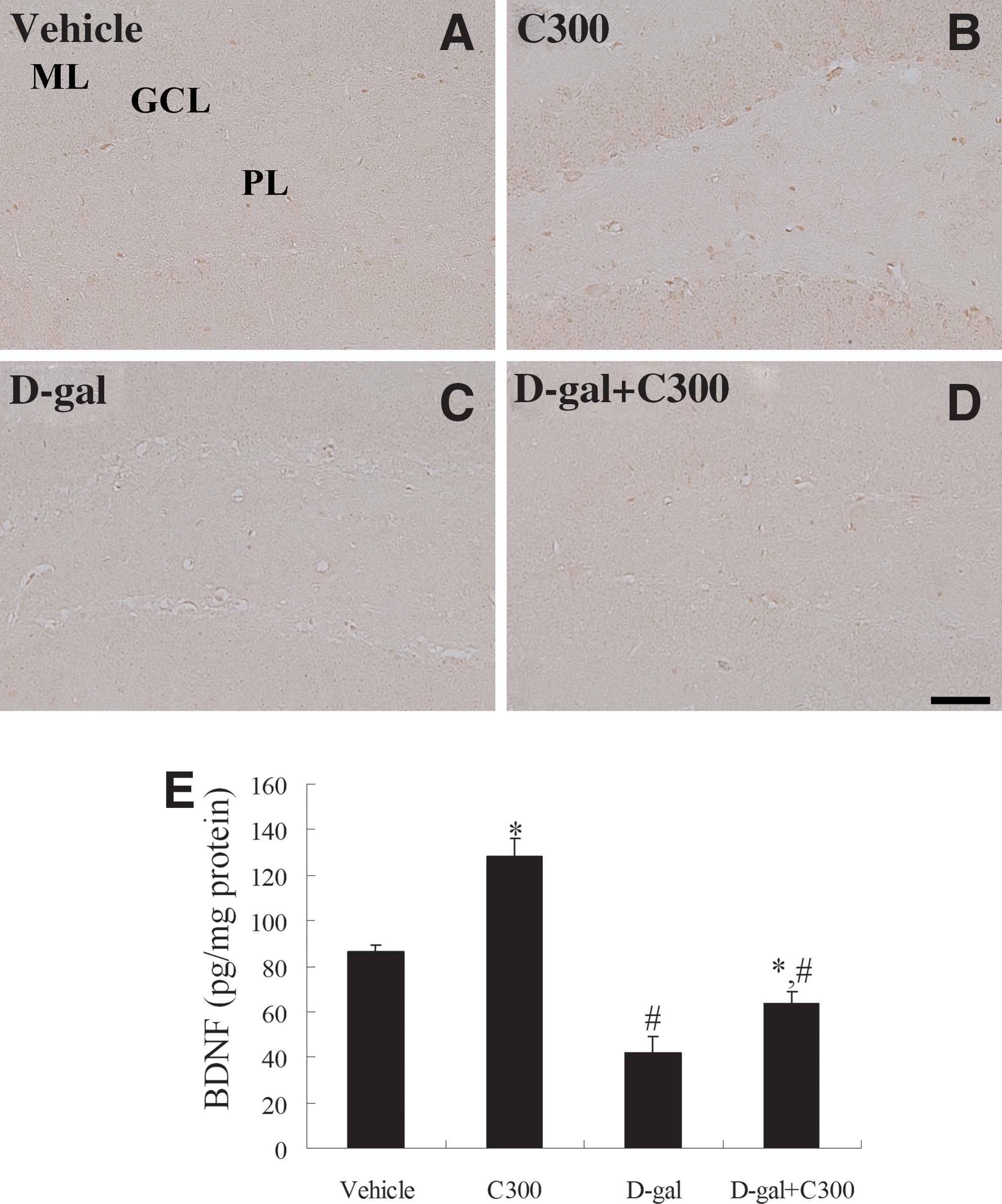
Immunohistochemistry for brain-derived neurotrophic factor (BDNF) in the dentate gyrus of
Discussion
Chronic systemic exposure to D-gal in mice gives rise to the acceleration of senescence and includes shortened life-span, 3 cognitive deficits, 46 increased ROS, and suppressed antioxidant enzyme activity. 2,3 In the present study, we found that curcumin extract protected against memory impairment in the D-gal induced aged mice. The administration of C300 ameliorated the D-gal-induced increased latency times and decreased swimming speed, platform crossings, and spatial preference for the target quadrant compared to those reported in the counterpart control mice. This result is supported by previous studies that have shown that C. longa extract significantly ameliorates memory deficits in D-gal-induced senescence mice, 28 prevents decreases in acetylcholinesterase activity in aged rats 26,27 and mice with Alzheimer's disease 47 and chronic unpredictable stress condition. 29 It has reported that the protective effect of curcumin on memory deficit is mediated by prevention of oxidative stress. Chronic curcumin oral treatment significantly improved the colchicine-induced cognitive impairment in mice by decreasing lipid peroxidation. 48,49 In addition, curcumin reversed ethanol-induced memory deficit via suppression of nitric oxide synthase/nitric oxide pathway. 50 Curcumin shows neuroprotective effects against ethanol-, aluminum-, and iron-induced neurotoxicity in rodents. 51,52 In the okadaic acid-induced impairment model, oral administration of curcumin significantly improves the memory function as assessed by both Morris water maze and passive avoidance tests. 53 Under normal aging and increased oxidative stress conditions, memory function declines due to decreases in BDNF. 54,55 On the other hand, enhanced BDNF improved learning and memory impairments. 56,57 It has been reported that BDNF is closely related to learning and memory by directly examining its role in a variety of learning paradigms in rodents. 58,59 In the present study, we found that C300 administration enhanced BDNF expression in the hippocampus (Fig. 7). This result may suggest that enhanced BDNF expression by curcumin may prevent memory impairment induced by the D-gal aging model.
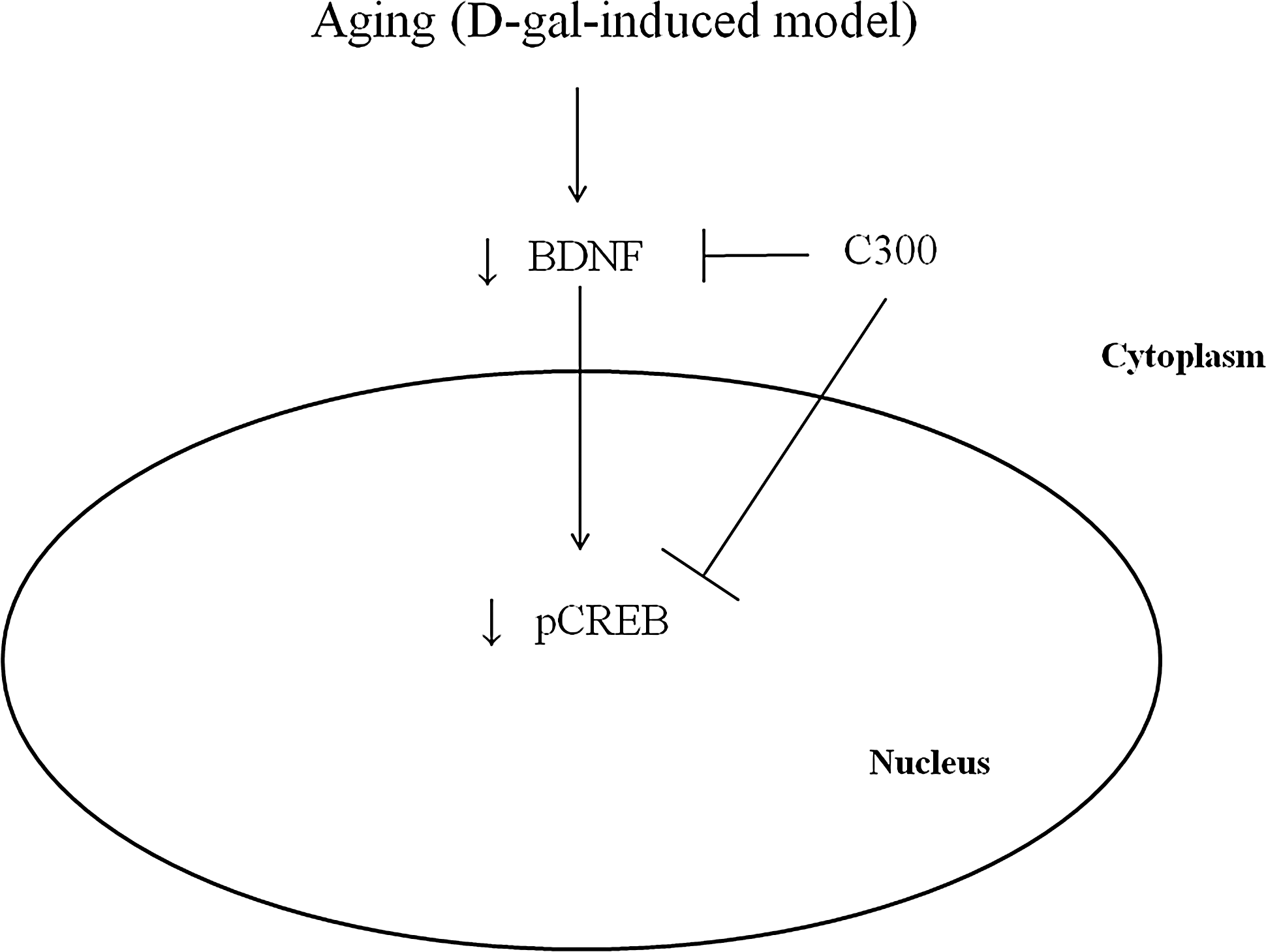
A proposed mechanistic diagram of C300 action in this study.
Numerous studies have reported that aging decreases the proliferation of progenitor cells in the dentate gyrus of the hippocampus. 2,33 –39 In this study, we confirmed that the administration of D-gal significantly decreased cell proliferation and neuroblast differentiation as seen in the aging process. The administration of C300 significantly increased cell proliferation and neuroblast differentiation in adult mice. It has been reported that the administration of curcumin significantly increases embryonic neural stem cell viability and proliferation. 60 In the D-gal-induced aging model, C300 significantly mitigated the D-gal-induced decrease in cell proliferation and neuroblast differentiation. It has been reported that curcumin ameliorates the stress-induced impairment of neurogenesis in rats. 61 However, the detailed mechanism involved in the enhanced neurogenesis observed by the treatment with curcumin remains unknown.
It has been reported that curcumin can reverse the decreased levels of monoamines, including serotonin, noradrenalin, and dopamine, in the rat brain due to olfactory bulbectomy 62 and exposure to conditions of stress. 63 –65 Activation of serotonin receptors coupled to cAMP production and CREB activation can induce transcription of the BDNF gene. 66 BDNF plays a central role in brain development and plasticity by opposing neuronal damage and promoting neurogenesis and cell survival. 67,68 BDNF-tyrosine kinase B signaling leads to phosphorylation and activation of transcription factors such as CREB, which induces gene expression and long-lasting synaptic changes. 69,70 In the present study, we observed that the administration of C300 significantly increased BDNF immunoreactivity and protein levels in the dentate gyrus. In addition, the administration of C300 significantly increased pCREB immunoreactivity in the dentate gyrus of adult and aging mice. This result is supported by a previous study demonstrating that the administration of curcumin increases hippocampal neurogenesis in chronically stressed rats via upregulation of the 5-HT1A receptor and BDNF in the hippocampus. 61 In addition, curcumin protects neurons from chronic stress, depression, and glutamate excitotoxicity by activating BDNF levels. 71 –75 In a depression-like model induced by olfactory bulbectomy, curcumin significantly ameliorated the hyperactivity in an open field test as well as reducing BDNF levels in the whole brain. 76
In conclusion, the present study indicates that the administration of C300 mitigates D-gal-induced memory impairment, as well as the reduction of cell proliferation and neuroblast differentiation, through the activation of CREB and BDNF in the dentate gyrus.
Footnotes
Acknowledgments
This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning (2011-0013933).
Author Disclosure Statement
No competing financial interests exist.