
Abstract
Inulin, a group of dietary fibers, is reported to improve the metabolic disorders. In the present study, we investigated the effects of chicory inulin on serum metabolites of uric acid (UA), lipids, glucose, and abdominal fat deposition in quail model induced by a purine-rich diet. In this study, 60 male French quails were randomly allocated to five groups: CON (control group), MOD (model group), BEN (benzbromarone-treated group), CHI-H (high-dosage chicory inulin-treated group), and CHI-L (low-dosage chicory inulin-treated group). The serum UA level was significantly increased in the model group from days 7 to 28, as well as triglyceride (TG) and free fatty acid (FFA) increased later in the experimental period. The abdominal fat ratio was increased on day 28. Benzbromarone can decrease UA levels on days 14 and 28. The high and low dosage of chicory inulin also decreased serum UA levels on days 7, 14, and 28. The abdominal fat ratio, activity, and protein of acetyl-CoA carboxylase (ACC) were decreased in chicory inulin-treated groups. The activities of xanthine oxidase (XOD) and fatty acid synthase (FAS) were increased in the model group and decreased in the benzbromarone and chicory inulin groups. This study evaluated a quail model of induced hyperuricemia with other metabolic disorders caused by a high-purine diet. The results indicated that a purine-rich diet might contribute to the development of hyperuricemia, hypertriglyceridemia, and abdominal obesity. Chicory inulin decreased serum UA, TG, and abdominal fat deposition in a quail model of hyperuricemia by altering the ACC protein expression and FAS and XOD activities.
Introduction
I
Inulin is a soluble, but indigestible, dietary fiber. It is a naturally occurring oligosaccharide belonging to a group of carbohydrates known as fructans. Inulin is widely distributed in some edible plants, including asparagus, garlic, chicory, leek, onion, and artichoke, as a storage carbohydrate. 11 Inulin is widely used as a food supplement and additive with applications in fruit preparations, milk desserts, yogurt, fresh cheese, baked goods, chocolate, ice cream, and sauces. Chicory (Cichorium intybus L.) inulin is a linear fructose polymer, in which the fructose units are joined by a β (2→1) glycosidic linkage. 12 There is little information about the absorption, distribution, metabolism, and excretion (ADME) of chicory inulin in animals or humans. Data show that inulin acts as a prebiotic and is not hydrolyzed by pancreatic enzymes, but are broken down by specific bacteria in the hindgut, to help the large bowel function and improve the health of the organism. Inulin consumption leads to the acidification of digest and an enhancement of short-chain fatty acid (SCFA) production in the hindgut, especially propionate and butyrate. After absorption, propionate is thought to inhibit hepatic lipogenesis and positively influence the blood lipid profile, stable blood glucose, and also reduce the incidence of various kinds of cancer in human population and laboratory rodent animals, 11 –14 whereas the decreased digest pH is probably responsible for the stimulation of mineral absorption and bone health in laboratory animals. 15 The inulin used in this study was blended Fibrulose F97 purchased from Cosucra. The inulin had molecular weights of 2.07×103–9.34×103, which was measured by HPLC at the China National Institute of Metrology.
Quail is a well-established laboratory research animal because of its appropriate body size and excellent reproductive performance. 16 Furthermore, the purine and lipid metabolic pathways in quail are similar to humans. 17 Therefore, quail might be suitable for developing a hyperuricemia model. However, there are few reports regarding the effects of chicory inulin on quails with hyperuricemia and lipid metabolism. Therefore, this study was conducted to examine the effects of chicory inulin on serum UA, blood lipids, glucose, and abdominal fat deposition in a quail model induced by a purine-rich diet.
In this study, quails were fed with a purine-rich diet with added yeast extract (yeast extract is rich in purines and pyrimidines, which can increase the purine content of the diet). The UA level might be especially increased by a purine-rich diet in quails since they have a mutation in the urate oxidase (uricase) gene. In a previous study, we observed hyperuricemia and lipid metabolic disorders in quails induced by purine-rich diets. Benzbromarone, a nonpurine xanthine oxidase (XOD) inhibitor, can be used for the treatment of hyperuricemia and gout, and also can reduce fructose-induced hyperinsulinemia and hypertriglyceridemia. 5 Therefore, benzbromarone (clinically used as a drug) was used as a positive control for treating hyperuricemia and lipid metabolic disorders, as we aimed to observe the therapeutic effects of chicory inulin. The dosage of benzbromarone at 20 mg/kg and dosages of chicory inulin at 10 g/kg and 5 g/kg were used in the experiment according to our previous studies and the reference dosages used in rats and broiler chickens. 18 –20
Materials and Methods
Test products
Yeast extract was purchased from OXOID Co., Ltd. Benzbromarone tablets (50 mg) were purchased from Heumann Pharma GmbH. Chicory inulin used in the current study was a commercial product (Fibrulose F97; Cosucra). Chemical characteristics obtained from this product detected in our laboratory were the following: average molecular weight 1.38×103; degree of polymerization from 2 to 30; and free glucose less than 5%. UA, triglyceride (TG), and glucose kits were purchased from Beijing Leadman Group Co., Ltd. The acetyl-CoA carboxylase (ACC) primary antibody was purchased from Cell Signaling Technology. Free fatty acid (FFA), XOD, and fatty acid synthase (FAS) kits were obtained from Nanjing Jiancheng Bioengineering Co., Ltd. The Chicken Acetyl Coenzyme A Carboxylase ELISA kit was obtained from Life Sciences Advanced Technologies, Inc.
Equipment
Beckman CX4 automatic biochemical analyzer (HALI Beckman, Ltd.), Refrigerator Centrifuge (Jouan Ltd.), and 2000UV ultraviolet-visible spectrophotometer (Shanghai Spectrum Instruments Co., Ltd.). Upright microscope system (Leica DMRXA) and image analysis system (Q550CW). Microplate reader Sunrise™ Tecan Group Ltd.
Quails, diets, and management
All procedures for handling the quails were approved by the Animal Care and Ethics Committee in Beijing University of Chinese Medicine. Sixty male French quails obtained from a commercial breeding facility (Beijing Deling Quail Farm) were randomly divided into the following five groups: CON (control group fed with commercial quail feedstuff), MOD (model group fed a diet with added yeast extract powder), BEN (benzbromarone-treated group fed a diet with added yeast extract powder and treated with 20 mg/kg benzbromarone water solution by intragastric administration, using a ball-tip needle), CHI-H (high-dosage chicory inulin-treated group fed a diet with added yeast extract powder and treated with 10 g/kg chicory inulin water solution by intragastric administration, using a ball-tip needle), and CHI-L (low-dosage chicory inulin-treated group fed a diet with added yeast extract powder, and treated with 5 g/kg chicory inulin water solution by intragastric administration, using a ball-tip needle). Control group quails were fed commercial quail diets. The model group and other treated groups were fed the commercial diet with added yeast extract powder (OXOID Ltd.), 15% of the ratio in weight. Twelve quails in each group were housed in 90×80×40 cm3 cages, under standard conditions of temperature, humidity, and ventilation. Animals were freely provided feedstuff and water during the experimental period. Feedstuff was purchased from the Beijing Deling Quail Farm, the components of the commercial quail feedstuff are shown in Table 1.
Provided by Beijing Deling Quail Farm.
Feedstuff was provided twice per day and any remaining food in each cage was weighed, and feed intake for one quail was calculated by dividing the total number of quails in each cage. The mean of feedstuff intake was determined at weekly intervals.
Sampling and measurements
Body weight was recorded weekly. Blood samples were collected from jugular veins after a 12-h fasting. Blood samples were allowed to clot at room temperature for 60 min and then centrifuged at 1250 g for 10 min; serum was separated and stored at −20°C for the later analyses. At the end of the experimental trail, all quails were killed and the abdominal fat pad (from gizzard to bursa of Fabricius) was weighed.
Immunohistochemical analysis
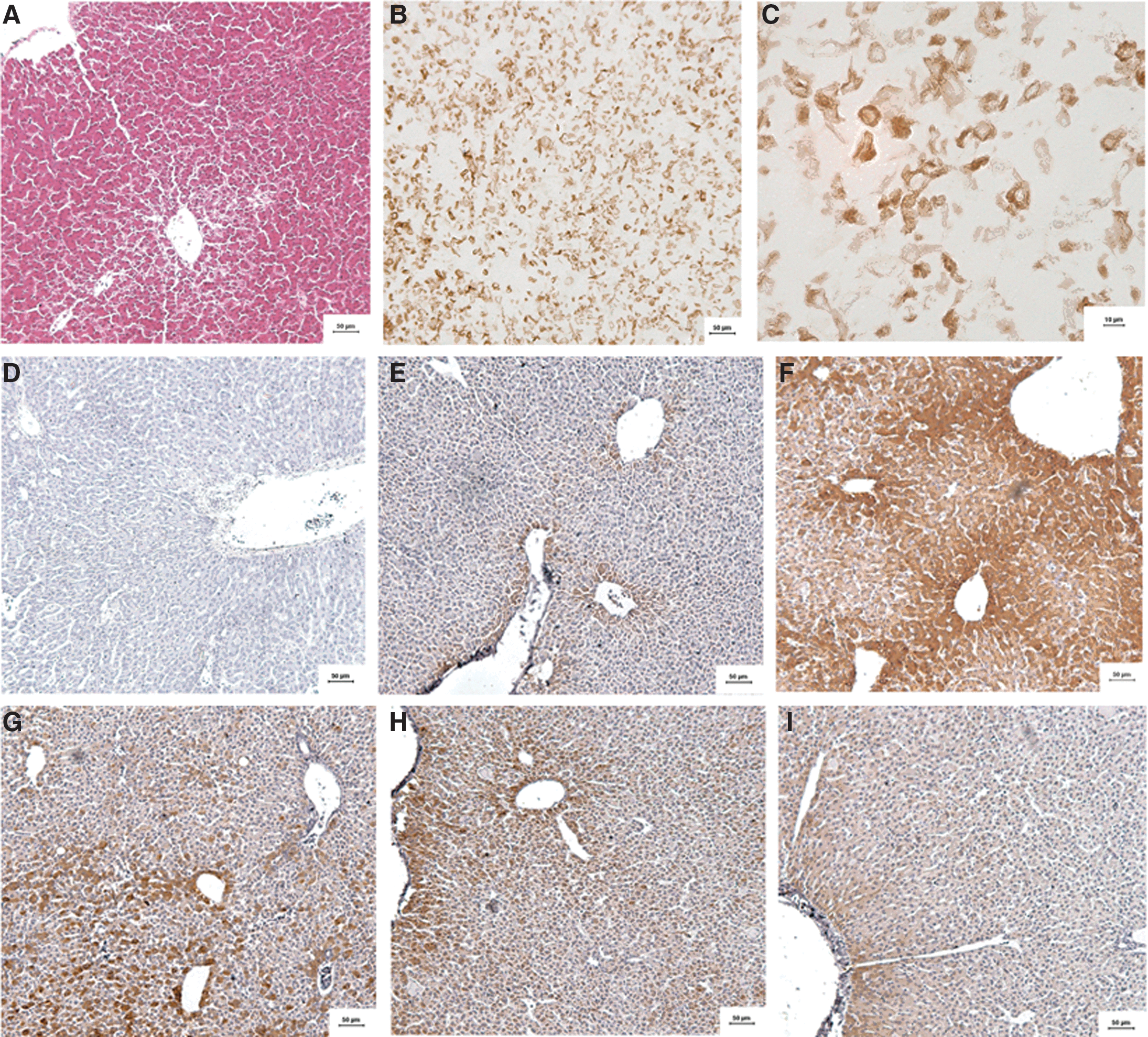
The liver tissues, fixed in 4% polyoxymethylene, were processed by routine histological procedures. Five-μm paraffin sections were prepared for immunohistochemical analysis. Protein expression of ACC in liver sections was detected by using anti-rat antibodies of p-ACC (Phospho-Acetyl-CoA Carboxylase [Ser79] Antibody #3661; Cell Signaling Technology). The sections were rinsed in TBST, boiled in 10 mM Tris/1 mM EDTA (pH 9.0) for 20 min, and then cooled to room temperature for 30 min to unmask antigen. With antigen unmasking, the sections were immersed in 0.3% hydrogen peroxide for 5 min. The slides were then incubated with specific antibodies at 4°C overnight and then rinsed with TBST thrice for 5 min. Sections were then incubated with the appropriate unbiotinylated secondary antibody (Zhongshan Goldenbridge) for 30 min at 37°C. Slides were then treated with a solution-containing DAB kit (Zhongshan Goldenbridge), incubated for 3 min, and rinsed under running water. As a positive control, 3T3-L1 culture cell sections (Cell Signaling Technology) were used for detecting the primary antibody; as a negative control, Phosphate Buffer Saline (PBS) was used instead of the primary antibody.
All measurements were performed with the automated upright microscope system (Leica DMRXA) and image analysis system (Q550CW). Five random images from each section obtained from five rats were randomly captured. The positive immunostained area in a total area under an image field of each section was calculated.
Analytical methods
The inulin commercial product was assayed by HPLC at the China National Institute of Metrology. Serum UA, TGs, and glucose (Glu) were measured by Beckman CX4 automatic biochemical analyzer. The activities of XOD and FAS were detected by the colorimetric method. Abdominal fat pad was weighted and abdominal fat ratios were calculated as abdominal fat pad weight (g) divided by body weight (g).
Statistical analysis
Values are expressed as mean and standard deviation (
Results
Growth performance
Birds were in good health throughout the entire experimental period. Body weight and dietary intakes were recorded. There was no significant difference in body weight among the five groups during the experimental period. There was a nonsignificant tendency for a lower dietary intake in quails receiving inulin compared with the control group.
Blood serum metabolites
Effect of chicory inulin on serum UA, TGs, FFA, total cholesterol, and glucose levels
The serum UA levels of each group over the experimental period are shown in Table 2. Compared with the control group, the levels of serum UA were significantly higher in the model group from 7 to 28 days (P<.05 and P<.01). Compared with the model group, the levels of UA were significantly decreased in the benzbromarone-treated group on days 14 and 28; the levels of UA were significantly decreased in the CHI-H- and CHI-L-treated group from 7 to 28 days (P<.05 and P<.01), and there were no statistically significant differences between the CHI-H and CHI-L groups. Serum was obtained from jugular veins in the morning after a 12-h fasting every 7 days, which was used for clinical chemistry assays. Compared with the control group, the levels of serum TGs were unchanged in the model group from 7 to 14 days and significantly increased from 21 to 28 days. Compared with the model group, the levels of TG were significantly decreased in the benzbromarone-treated group on day 21; the levels of TG were significantly decreased in the CHI-H-treated group from 21 to 28 days and the levels of TG were significantly decreased in the CHI-L-treated group on days 7, 21, and 28 (Table 2).
Values are mean±SD.
Means with different superscript lowercase letters (a,b,c) in the same column are significantly different (P<.05).
BEN, model group treated with benzbromarone; CHI-H, model group treated with chicory inulin 10 g/kg; CHI-L, model group treated with chicory inulin 5 g/kg; CON, control group; MOD, model group; TG, triglyceride; UA, uric acid.
The serum FFA levels were significantly increased on days 7 and 28. Compared with the model group, the levels of FFA were significantly decreased in the benzbromarone-, CHI-H-, and CHI-L-treated groups on days 7 and 28 (Table 3).
Groups are the same as Table 1. Values are mean±SD.
Means with different superscript lowercase letters in the same column are significantly different (P<.05).
There were no significant differences in serum total cholesterol or glucose among any of the groups, results not shown.
Effect of chicory inulin on abdominal fat deposition
In this study, abdominal fat deposition is expressed as abdominal fat pad weight (g) and abdominal fat ratio (%). The weight of abdominal fat pad tended to be higher in the model group on day 28 (P=.0596) but was not significant; abdominal fat ratios were increased in the model group on day 28. Compared with the model group, the weight of abdominal fat pad and abdominal fat ratios were significantly decreased in the benzbromarone-, CHI-H-, and CHI-L-treated groups on day 28 (P<.05 and P<.01, Table 4).
Groups are the same as Table 1. Values are mean±SD.
Means with different superscript lowercase letters in the same column are significantly different (P<.05), Unpaired Student's t-test.
Purine metabolic enzyme XOD and lipid metabolic enzymes FAS, ACC
There were no significant differences in serum XOD activities in CON, MOD, BEN, and CHI-L groups, but were decreased in the CHI-H group on day 7. The activities of serum XOD were increased in the model group on day 28. Compared with the model group, the activities of XOD were significantly decreased in the benzbromarone-, CHI-H-, and CHI-L-treated groups on day 28 (P<.01, Table 5).
Groups are the same as Table 1. Values are mean±SD.
Means with different superscript lowercase letters in the same column are significantly different (P<.05).
The liver fatty acid synthase (FAS) activities were increased in the model group on day 28. Compared with the model group, liver FAS activities were significantly decreased in CHI-H- and CHI-L-treated groups on day 28 (P<.05 and P<.01). The activities of liver ACC were increased in the model group on day 28 compared with the control group. There were no significant differences in ACC activities among the benzbromarone-, CHI-H-, and CHI-L-treated groups (Table 6).
Groups are the same as Table 1. Values are mean±SD.
Means with different superscript lowercase letters in the same column are significantly different (P<.05).
ACC, acetyl-CoA carboxylase; FAS, fatty acid synthase.
Liver ACC activities and ACC protein expression
There was no immunoreactivity in the sections set aside for negative control. ACC positivity was indicated by cytoplasmic golden brown staining. In the control group, only a small area was positive for immunostaining. ACC immunostaining was strongly presented in the portal area of liver tissue, and moderate positivity in the wall of the portal veins. ACC protein expression in the model group was significantly higher than that in the control group (P<.01), whereas an evaluation of ACC protein expression in chicory inulin-treated (CHI-H and CHI-H) and benzbromarone-treated groups was significantly lower compared with the model group (P<.01). Details are shown in Table 7 and Figure 1A–I.
Values are mean±SD.
Means with different superscript lowercase letters in the same column are significantly different (P<0.05).
Discussion
The similarity of metabolic characteristics between human and avian species
Quail (Coturnix japonica [Coja]), a common avian species, belongs to the same order (Galliformes) and family (Phasianidae) as the chicken. Quails have a long history as laboratory research animals, because of their appropriate body size and excellent reproductive performance. 16 The evolutionary distance between chickens and humans provides high specificity in detecting functional elements, both noncoding and coding. 17 About 60% of chicken protein-coding genes have a single human ortholog. Chicken and human 1:1 ortholog pairs exhibit the sequence conservation with a median amino acid identity of 75.3%. 17 UA is the oxidation (breakdown) product of purine metabolism. In most mammals, the enzyme uricase further oxidizes UA to allantoin. In contrast, in humans and avians, UA is the final product of purine metabolism due to the loss of uricase activity by various mutations of its gene, which lead to humans and birds having higher UA levels than other species. 1,2,21 UA metabolism in men and chickens is influenced by the amount of purine and protein in the diet. 8 –10,22,23 In addition, chicken apoAI is capable of transforming phospholipid vesicles into discoidal bilayer structures, similar in both size and apolipoprotein content to those produced for human apoAI under the same conditions. 24 Lipid-free chicken and human apoAI were equivalent in their ability to deplete cholesterol from this pool, as were intact chicken high-density lipoprotein (HDL) and human HDL. 25 Above all, there are several advantages to using quails as a metabolic disease model, especially for the model of purine and lipid metabolic disorders.
Correlationships among UA, blood lipids, and abdominal obesity
One common feature in patients with metabolic syndrome is an elevated UA; recent evidences supported a primary role for UA in mediating metabolic syndrome. 26,27 Epidemiological and laboratory studies have revealed a strong relationship between UA and abdominal obesity, hyperlipidemia, hypertension, insulin resistance, type 2 diabetes mellitus, ischemic heart disease, and cerebral apoplexy. 28 –32 The studies in our laboratory demonstrated that quails develop hyperuricemia at days 7 and 28 when fed with purine-rich diets. The serum TG and FFA concentrations increased later in the experimental period. In addition, the accumulation of abdominal fat increased by 73.40% in the model group. Our findings indicated that increased serum concentrations of UA, TG, and abdominal fat accumulation were present in the quail model. Furthermore, there are some other explanations about the pathology of hyperuricemia and hyperlipemia. The classical literature indicates that lifestyle is the main reason for hypertriglyceridemia in gout patients. 33 Matsuura et al. report that the synthesis of fatty acids in the liver is related to the de novo synthesis of purines, thus accelerating UA production. 30 Samant et al. report that chronic hyperuricemia can inactivate nitric oxide (NO), which leads to hyperlipidemia in diabetic patients. 34 Benzbromarone is an inhibitor of reabsorption of UA, although it does not meditate the enzymes of FAS and ACC and the level of serum TG was significantly decreased with decreasing UA. Above all, the features are similar to those in humans of metabolic syndrome (abdominal obesity and hypertriglyceride are consistently linked to hyperuricemia). 35 This study was consistent with the literature of previous epidemiological studies. 30,32,35
The medicinal function of chicory inulin
Chicory inulin is a kind of dietary fiber characterized by linear β (2→1) fructans. Inulin resists hydrolysis by intestinal digestive enzymes, so it is classified as a nondigestible carbohydrate. A series of animal studies demonstrated that inulin affects the metabolism of lipids primarily by decreasing triglyceridemia because of a decrease in the number of plasma VLDL particles, a suppressed triglyceridemia and only a relatively slight decrease in cholesterolemia, mostly in (slightly) hypertriglyceridemic conditions. 36 Inulin also affects gastrointestinal functions not because of its physicochemical properties, but rather because of its biochemical and physiological attributes. 12 In the current study, we demonstrated that chicory inulin significantly suppressed the level of serum UA, TG concentrations, and abdominal fat deposition in quails fed a purine-rich diet. The results indicated that chicory inulin was able to significantly reverse hyperuricemia, hypertriglyceridemic, and abdominal fat accumulation. Therefore, chicory inulin is a candidate agent for treating metabolic syndrome.
The role of XOD, FAS, and ACC in the metabolic disorders
In this study, we observed that the model quails had the pathologic features of hyperuricemia, hypertriglyceridemia, and abdominal fat accumulation. The mechanisms underlying the suppressive effects of chicory inulin on UA, TG, and abdominal fat accumulation might be related to the purine and lipid metabolic enzymes. XOD is an important enzyme in the purine metabolism pathway, which catalyzes the oxidative hydroxylation of the natural purine, hypoxanthine to xanthine, and finally to UA, which is excreted in the urine. 37,38 FAS is the enzyme that catalyzes the de novo synthesis of long-chain fatty acids from acetyl-coenzyme A (CoA) and malonyl-CoA in cells, the process uses reduced nicotinamide adenine dinucleotide phosphate (NADPH) as a cofactor. 39 Malonyl-CoA is catalyzed by the enzyme ACC, the rate-limiting step in the process of de novo fatty acid synthesis, and large amounts of NADPH (serving as electron donor) are consumed. 40,41 NADPH is produced in the pentose phosphate pathway, which also regulates UA production. 42 Several studies have demonstrated ACC might be a potential target for treating metabolic syndrome. 41,43
Abdominal fat accumulation associated with the overproduction of UA is another link to metabolic syndrome, which has been suggested by several investigators. Increased ACC protein expression and activity contribute to abdominal fat accumulation, which provides excessive FFA in the portal vein and accelerates the overproduction of very low-density lipoprotein, which causes hypertriglyceridemia. This also accelerates the de novo purine synthesis by NADPH produced in the pentose phosphate pathway, which increases UA production. 44,45
Therefore, we can see that ACC plays an important role in the process of fatty acid synthesis and UA synthesis as well. The activity of the rate-limiting enzyme ACC might alter the consumption of NADPH; on the other hand, UA production might be accelerated through the pentose phosphate pathway as the reduction of NADPH. Accordingly, we developed the hypothesis that the increased activity of ACC contributed to the mechanisms underlying the biochemical changes in UA, TG, and abdominal fat accumulation. In this study, we observed that ACC activities were elevated and protein overexpression in the quail model fed a purine-rich diet. Furthermore, ACC inhibitory activities and hypouricemic effects of chicory inulin were observed during the experimental period. We also found that chicory inulin significantly decreased the activities of the purine metabolic enzyme, XOD, and lipid metabolic enzyme, FAS, in the quail model fed a purine-rich diet. To our knowledge, these results suggest for the first time that the ACC activity and protein expression are correlated with UA and TG levels and abdominal fat deposition in quail and involved in the regulation of purine and lipid metabolism by chicory inulin.
In conclusion, this study demonstrates that a purine-rich diet induces hyperuricemia in quails. Chicory inulin lowered serum UA and TG concentrations and decreased abdominal fat deposition by altering the purine metabolic enzyme XOD and lipid metabolic enzyme FAS. ACC might play an important role in the development of hyperuricemia, hypertriglycemia, and abdominal obesity.
Footnotes
Acknowledgments
The authors thank Mr. Masaoki Tsudzuki for his mail with the information of quail characters and Mr. Xudong Jia for his technical support on the preparation of paraffin sections. This work was supported, in part, by the Research and Experimental Center in Beijing University of Chinese Medicine and China National Institute of Metrology. This research was supported by the Major State Basic Research Development Program of China (973 Program) No. 2007CB512605, the Creative Research Team Program of Beijing University of Chinese Medicine 2011-CXTD-014, and the Research Fund for the Doctoral Program of Higher Education of China, 20120013130002, 20130013120001.
Author Details
Zhijian Lin: Responsible for execution of experimental procedures and primary preparation of the manuscript; Bing Zhang: Responsible for obtaining funding, designing, and supervision of the study, and assisted with manuscript preparation; Xiaoqing Liu: Responsible for study implementation and assisted with measurement of metabolic enzymes; Rui Jin: Responsible for analysis of data and assisted with manuscript preparation; Wenjing Zhu: Responsible for assisting with execution of experimental procedures.
Author Disclosure Statement
The authors declare that they have no competing interests. All authors read and approved the final manuscript.