
Abstract
The present study investigated the effect of Rubus coreanus Miquel (RCM) on scopolamine-induced memory impairments in ICR mice. Mice were orally administrated RCM for 4 weeks and scopolamine was intraperitoneally injected into mice to induce memory impairment. RCM improved the scopolamine-induced memory impairment in mice. The increase of acetylcholinesterase activity caused by scopolamine was significantly attenuated by RCM treatment. RCM increased the levels of acetylcholine in the brain and serum of mice. The expression of choline acetyltransferase, phospho-cyclic AMP response element-binding protein, and phospho-extracellular signal-regulated kinase was significantly increased within the brain of mice treated with RCM. The brain antioxidant enzyme activity decreased by scopolamine was increased by RCM. These results demonstrate that RCM exerts a memory-enhancing effect via the improvement of cholinergic function and the potentiated antioxidant activity in memory-impaired mice. The results suggest that RCM may be a useful agent for improving memory impairment.
Introduction
I
Memory is a complicated process that involves various neuronal pathways and neurotransmitters. It is well established that the cholinergic neuronal system plays a crucial role in memory. 1 Cholinergic deficits are neuropathological events that are constantly associated with memory impairment. Several studies revealed that cholinergic deficits are correlated with the severity of memory impairment in AD patients and aged humans. 2,3 It has been suggested that elevations in the levels of acetylcholine, a cholinergic neurotransmitter, might help to improve memory impairment observed in AD patients. 4 Acetylcholinesterase (AChE) plays a key role in the hydrolysis of acetylcholine, and thus it is responsible for regulating acetylcholine levels. 5,6 For this reason, the inhibition of AChE activity has been suggested as a strategy for improving memory impairment. Recently, many attempts have been made to reverse memory impairment and treat AD by increasing the availability of acetylcholine via AChE inhibitors. 7 However, the few currently approved AChE inhibitors for treating patients with memory impairment cause various side effects such as hepatotoxicity. 8 Therefore, it is necessary to seek safe and effective alternatives for patients with memory impairment.
Scopolamine is a tropane alkaloid drug that exhibits competitive antagonism at muscarinic acetylcholine receptors by interfering with cholinergic transmission in the central nervous system. 9 It causes memory impairments in healthy young subjects that parallel memory impairment seen in nondemented drug-free elderly subjects 10 and AD patients. 11 Thus, the scopolamine-treated animal model is very useful for investigating memory studies in which the cholinergic system is involved.
Rubus coreanus Miquel (RCM) is a type of raspberry from south-east Asian countries including Korea, and is used as a traditional herbal medicine for the treatment of various diseases, such as enuresis, asthma, and allergic diseases. 12 It has been reported that the bioactive constituents of RCM are flavonoids, tannins, triterpenosides, anthocyanins, and numerous polyphenolic compounds, which have antinociceptive, anti-inflammatory, antigastropathic, antirheumatic, antioxidant, and anticancer effects. 13 –17
Even though numerous biological activities of RCM have been reported, the effect of RCM on memory impairment has not been studied. In a preliminary in vitro experiment, we found that RCM exhibited the most potent inhibition of AChE activity. In this study, we explored the possibility that RCM functions as a memory-improving agent against scopolamine-induced memory deficit in mice. We showed that the administration of RCM significantly improved memory in memory-impaired ICR mice.
Materials and Methods
Preparation of lyophilized RCM
The cultivated berries (RCM), picked at a ripened stage in July 2011, were purchased in Hoengseong (Gangwon-do, Korea). The berries were picked, washed with water, and frozen at −20°C on-site. The frozen berries were lyophilized to dry powder using a freeze dryer (Lyoph-Pride; IlShinBioBase, Dongducheon, Korea) and stored at −20°C until use in experimental studies.
Animals and treatment
Four-week-old male ICR mice were purchased from Orient Bio, Inc. (Seongnam, Korea). The mice were acclimated to the laboratory conditions while providing free access to a commercial rodent diet (Superfeed Co., Wonju, Korea) and water. The mice were acclimated for 1 week prior to use and maintained throughout the study in a controlled environment: 24±2°C, 50±10% relative humidity, and a 12 h light/dark cycle. After acclimatization, the mice were randomly divided into four groups: (1) control group, (2) scopolamine group, (3) scopolamine +50 mg RCM/kg body weight/day group, and (4) scopolamine +100 mg RCM/kg body weight/day group. The mice were subjected to oral gavage with vehicle (saline) or RCM dissolved in saline (50 mg or 100 mg/kg body weight/day) for 4 week. For this period, the mice were fed on the AIN-76 diet (Research Diets, Inc., New Brunswick, NJ, USA) and water ad libitum. To induce memory impairment, all mice except the control group were intraperitoneally injected scopolamine (1 mg/kg body weight) 30 min prior to each test. All experiments were conducted in accordance with the protocols approved by the Animal Care and Use Committee of the Hallym University, Korea (ethical approval number: Hallym 2012–16).
Passive avoidance test
The step-through passive avoidance test was performed using the Gemini Avoidance System (Gemini San Diego, Inc., San Diego, CA, USA). For the training trial, the mice were initially placed in the light compartment and the door between the two compartments was opened 10 s later. When the mice entered the dark compartment, the door was closed automatically and an electrical foot shock (0.3 mA) was delivered for 2 s through the stainless steel rods. Twenty-four hours after the training trial, the mice were placed in the light compartment and the guillotine door was opened. The escape latency time for entering the dark compartment was measured. If the mice did not enter the dark compartment within 300 s, then the test was terminated and a ceiling score of 300 s was assigned.
Morris water maze test
A spatial memory test was performed by the method of Morris 18 using the EthoVision Maze test system (Noldus Information Technology, Wageningen, Netherlands). On the first day (day 1) of the experimental procedure, the mice performed single swim training for 60 s in the absence of the platform, to get used to the pool. On the days following, the mice were given two trial sessions each for four consecutive days. During each trial, the time taken to swim to the platform (escape latency) was recorded. This parameter was averaged for each session of trials and for each mouse. Once the mouse located the platform, it was permitted to remain on it for 10 s. If the mouse did not locate the platform within 120 s, then it was placed on the platform for 10 s and then removed from the pool (trial 1). The mouse was given a second trial (trial 2) with an intertrial interval of 20 min for four consecutive days. The point of entry for the mouse into the pool and the location of the platform for escape remained unchanged between trial 1 and trial 2, but it was changed each day thereafter.
Preparation of brain tissue samples
After the Morris maze water test, the mice were anesthetized by the intraperitoneal injection of 240 mg/kg body weight of 2,2,2-tribromoethanol (Avetin®; Sigma-Aldrich Co., St. Louis, MO, USA) 30 min after injection of scopolamine, and then the brains were removed. The cerebral cortex and hippocampus were dissected and removed from the brain. Each part of the brain was homogenized in ice-cold sodium phosphate buffer (100 μM, pH 7.4), respectively. The homogenates were centrifuged at 1000 g at 4°C for 5 min, and the supernatant was collected for assay. Protein concentrations were determined with a DC protein assay kit (Bio-Rad, Hercules, CA, USA).
Measurement of AChE activity
The activities of AChE were measured using Ellman's coupled enzyme assay. 19 In brief, the reaction mixture contained 0.06 mL of 100 mM sodium phosphate (pH 8.0), 0.02 mL of 20 mM 5,5′-dithio-bis(2-nitrobenzoic acid), and 0.1 mL of homogenate and was preincubated at 37°C for 5 min, and then 0.02 mL of 10 mM acetylthiocholine was added. After incubation at 37°C for 3 min, absorbance was measured at 410 nm. The activities of AChE were calculated as units per mg protein.
Measurement of acetylcholine levels
The levels of acetylcholine in the cerebral cortex, hippocampus, and serum were measured using EnzyChrom™ Acetylcholine Assay kit (BioAssay Systems, Hayward, CA, USA) according to the manufacturer's instructions.
Measurement of superoxide dismutase and glutathione peroxidase activities
The activities of superoxide dismutase (SOD) and glutathione peroxidase (GPx) in the cerebral cortex and hippocampus of the brain were estimated using their respective assay kits (Cayman Chemical Company, Ann Arbor, MI, USA) according to the manufacturer's instructions.
Immunohistochemistry
The mice were anesthetized by intraperitoneal injection of 240 mg/kg body weight of 2,2,2-tribromoethanol (Sigma-Aldrich Co.) and then perfused through the ascending aorta with normal saline followed by 4% paraformaldehyde (pH 7.4). The brain was removed from the skull, postfixed, and cryoprotected with 30% sucrose at 4°C. Coronal sections were cut at a thickness of 45 μm using a sliding microtome (SM2000R; Leica, Nussloch, Germany). The sections were immunostained using an Elite ABC kit (Vector Laboratories, Burlingame, CA, USA). In brief, the sections were incubated with antibodies against choline acetyltransferase (ChAT, 1:200 dilution; Millipore Corporation, Billerica, MA, USA), phospho-cyclic AMP response element-binding protein (phospho-CREB, 1:200 dilution; Cell Signaling Technology, Beverly, MA, USA), and phospho-extracellular signal-regulated kinase (phospho-ERK, 1:200 dilution; Cell Signaling Technology) in phosphate-buffered saline containing 0.5% bovine serum albumin for 24 h at 4°C. The sections were incubated with biotinylated secondary antibody, and then incubated for 90 min in ABC reagent to visualize immunoreactivity. Finally, the sections were incubated in a solution containing 3,3′-diaminobenzidine (Sigma-Aldrich Co.). The sections were mounted, air-dried, and dehydrated through graded ethanols; cleared in histoclear; and coverslipped. We observed each coronal section under a light microscope to examine the medial septum, gyrus, and hippocampus CA3 regions, individually. Images were captured at 200× magnification, and immune-positive cells were quantified with a Carl Zeiss AxioImager microscope and Image M1 Software (Carl Zeiss, Jena, Germany).
Statistical analysis
All measurements were performed by an independent investigator blinded to the experimental conditions. The data were expressed as the means±standard error of the means and analyzed via analysis of variance. The differences between the treatment groups were assessed via Duncan's multiple-range tests. Differences were considered significant at P<.05.
Results
Oral administration of RCM improves scopolamine-induced memory impairment in mice
We first evaluated memory functions using the passive avoidance test. The step-through latency in the scopolamine-treated group was markedly shorter than in the control group. The reduced step-through latency caused by scopolamine was significantly prevented by pretreatment with 50 and 100 mg/kg body weight/day of RCM for 4 weeks. However, no significant differences were observed between RCM-treated groups (Fig. 1).
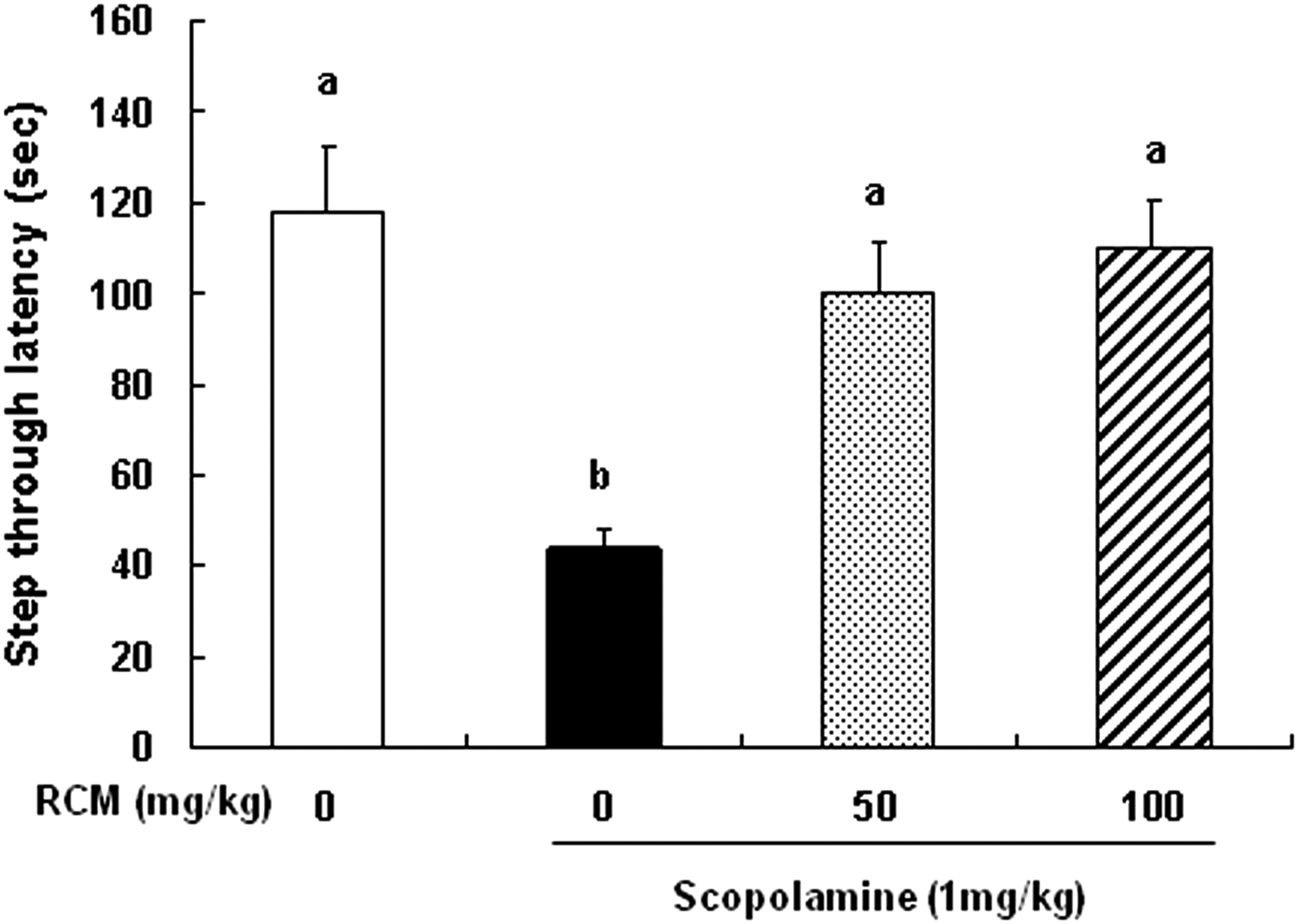
Effect of Rubus coreanus Miquel (RCM) on the scopolamine-induced memory deficit in the passive avoidance test. The mice were subjected to oral gavage with vehicle (saline) or RCM (50 or 100 mg/kg body weight/day) for 4 weeks. Memory deficit was induced by scopolamine injection (1 mg/kg body weight, i.p.) 30 min prior to the training trial. Twenty-four hours after the training trial, the mice were again placed in the light compartment. The latency to enter the dark compartment was measured. Each bar represents the mean±standard error of the mean (SEM, n=10). Means without a common letter differ, P<.05.
We next investigated the effect of RCM on scopolamine-induced spatial memory impairments using the Morris water maze test. The normal control mice rapidly learned the location of the submerged hidden platform, so the escape latencies gradually decreased from 120±0 s on the first day to 56.9±4.8 s on the fourth day. The scopolamine-treated mice exhibited longer escape latencies throughout training than the normal control mice. The administration of RCM significantly reduced the escape latencies extended by scopolamine injection. However, no statistically significant difference occurred between the groups receiving 50 and 100 mg/kg body weight/day of RCM (Fig. 2).
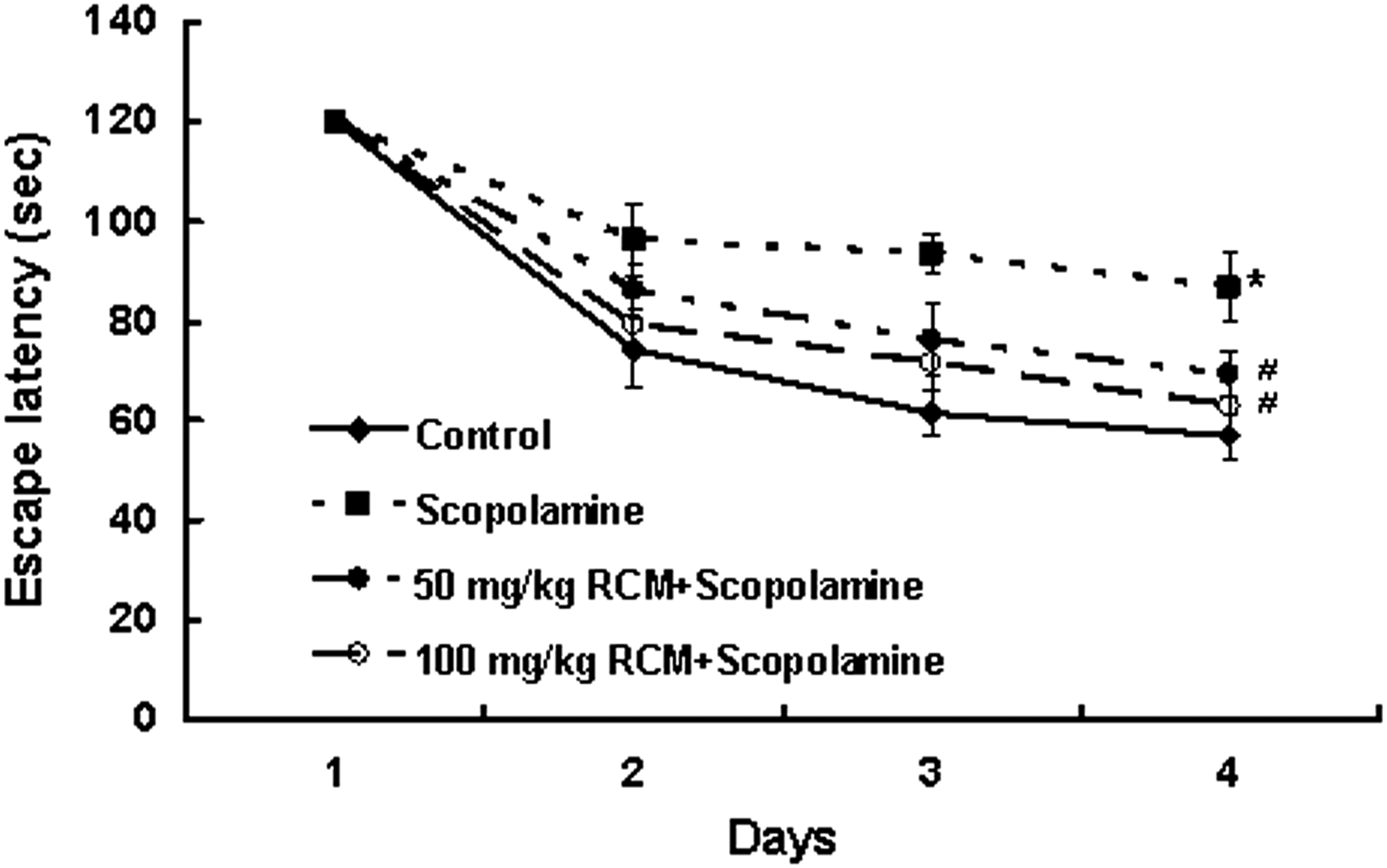
Effect of RCM on the scopolamine-induced memory deficit in the Morris water maze test. The mice were subjected to oral gavage with RCM and memory deficit was induced with scopolamine as described in Figure 1. Mice were given two sessions of trials each day for four consecutive days. The swimming time required for a mouse to escape to the platform was recorded in each day. Data represent the mean±SEM (n=10). *Significantly different from control group, P<.05. #Significantly different from scopolamine-treated group, P<.05.
RCM administration causes the alteration of AChE activities and acetylcholine levels in mice
To understand the memory-improving mechanism of RCM, we estimated AChE activities in the cerebral cortex, hippocampus, and serum of mice. AChE activities were increased by the injection of scopolamine in the cerebral cortex, hippocampus, and serum as compared with the vehicle-injected control group. The treatment with RCM significantly attenuated the enhancement of this enzyme activity induced by scopolamine injection in the cerebral cortex, hippocampus, and serum, respectively (Table 1).
The mice were subjected to oral gavage with RCM for 4 weeks and induced memory deficit with scopolamine as described in Figure 1. The mice were euthanized 30 min after scopolamine injection. The brains were removed and the sera were prepared. Values represent the mean±SEM (n=5). Means without a common letter differ, P<.05.
RCM, Rubus coreanus Miquel; BW, body weight; SEM, standard error of the mean.
Scopolamine treatment significantly reduced the levels of acetylcholine in the cerebral cortex, hippocampus, and serum of mice. The administration of RCM significantly prevented this reduction of acetylcholine in the cerebral cortex, hippocampus, and serum of scopolamine-treated mice (Table 2).
The mice were subjected to oral gavage with RCM for 4 weeks and induced memory deficit with scopolamine as described in Figure 1. The mice were euthanized 30 min after scopolamine injection. The brains were removed and the sera were prepared. Values represent the mean±SEM (n=5). Means without a common letter differ, P<.05.
RCM administration prevented the scopolamine-lowered expression of ChAT, phospho-CREB, and phospho-ERK in brain
The expression of ChAT, phospho-CREB, and phospho-ERK in the brain was analyzed using immunohistochemistry. In the medial septum region, the numbers of ChAT-immunoreactive neurons significantly decreased in the scopolamine-treated group as compared with the vehicle-treated control group. The administration of RCM prevented the losses of ChAT-immunoreactive neurons in the medial septum region (Fig. 3). The phosphorylation of CREB in the dentate gyrus region was decreased in the mice treated with scopolamine, and this decrease was suppressed by RCM administration in a dose-dependent manner (Fig. 4). Additionally, the treatment with scopolamine inhibited the phosphorylation of ERK in the hippocampus CA3 region. The administration of RCM also prevented scopolamine-induced decrease in ERK phosphorylation in the hippocampal CA3 region in the mouse brains (Fig. 5).
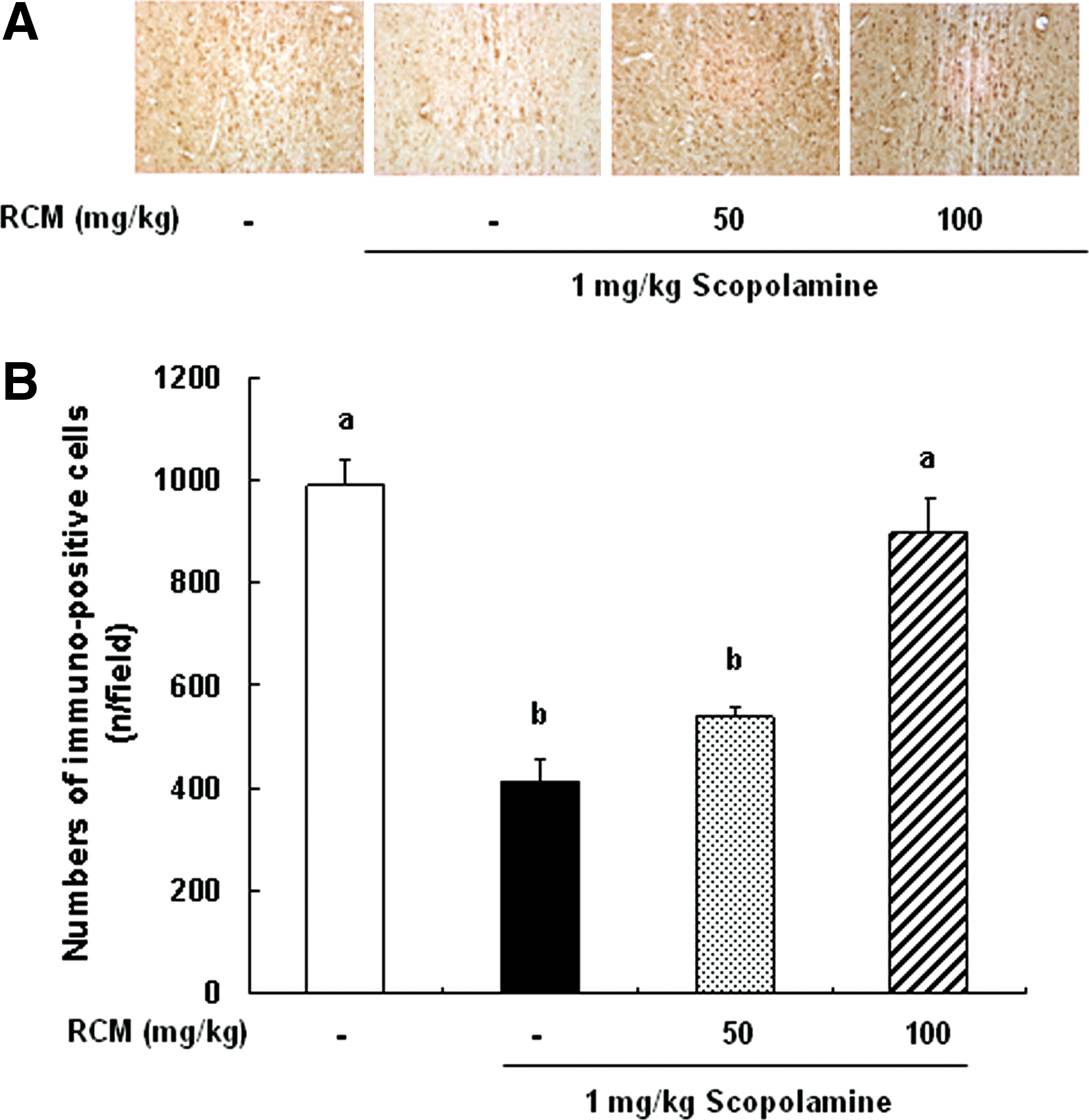
Effect of RCM on the expression of choline acetyltransferase (ChAT) in medial septum of brain. The mice were subjected to oral gavage with RCM and memory deficit was induced with scopolamine as described in Figure 1. The mice were perfused 30 min after scopolamine injection and the brains were removed. Brain tissues were stained with antibody raised against ChAT, and then immunostained using the Elite ABC kit.
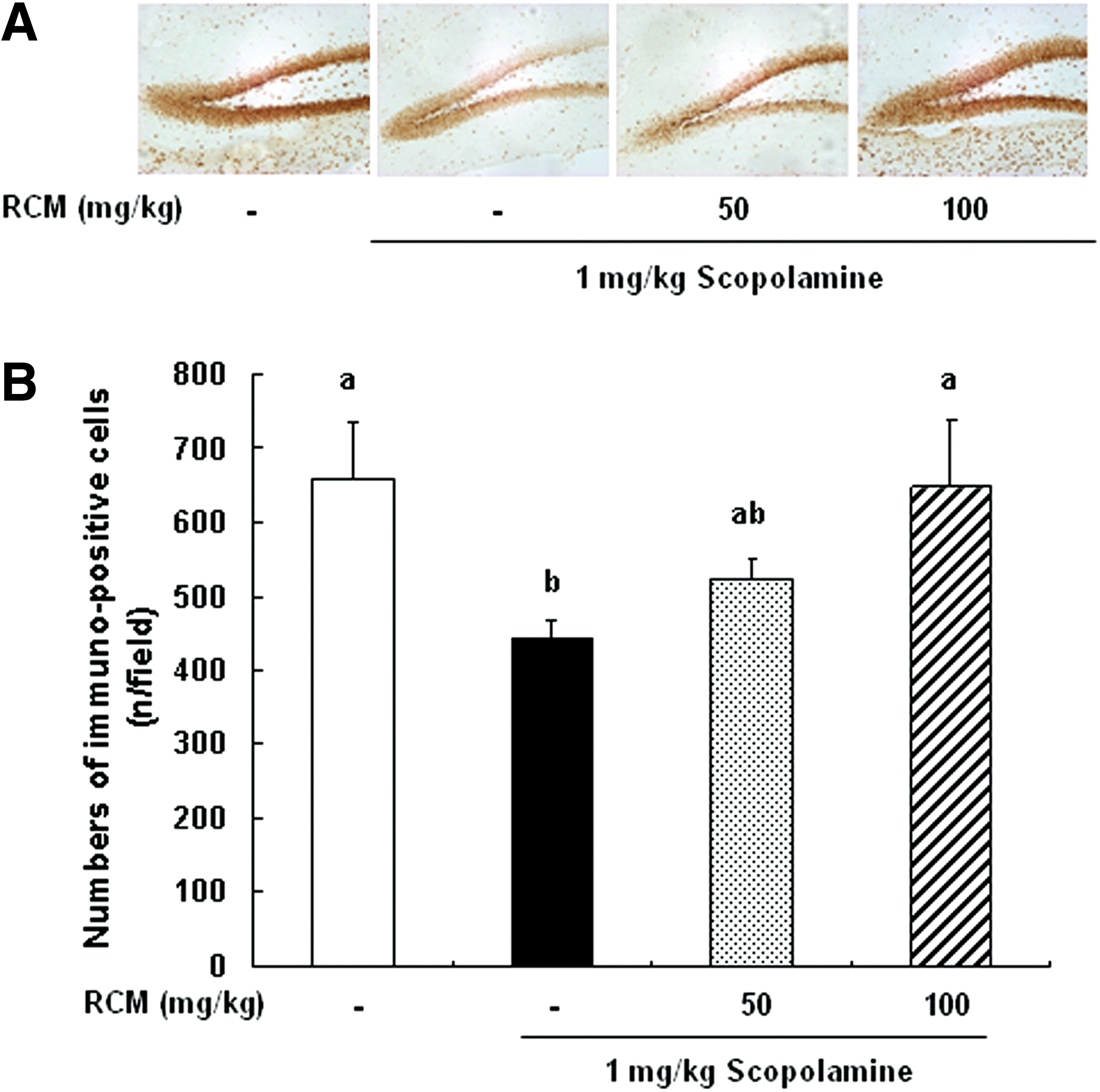
Effect of RCM on the phosphorylation of cyclic AMP response element-binding protein (CREB) in the dentate gyrus of brain. The mice were subjected to oral gavage with RCM and memory deficit was induced with scopolamine as described in Figure 1. The mice were perfused 30 min after scopolamine injection and the brains were removed. Brain tissues were stained with antibody raised against phospho-CREB, and then immunostained using Elite ABC kit.
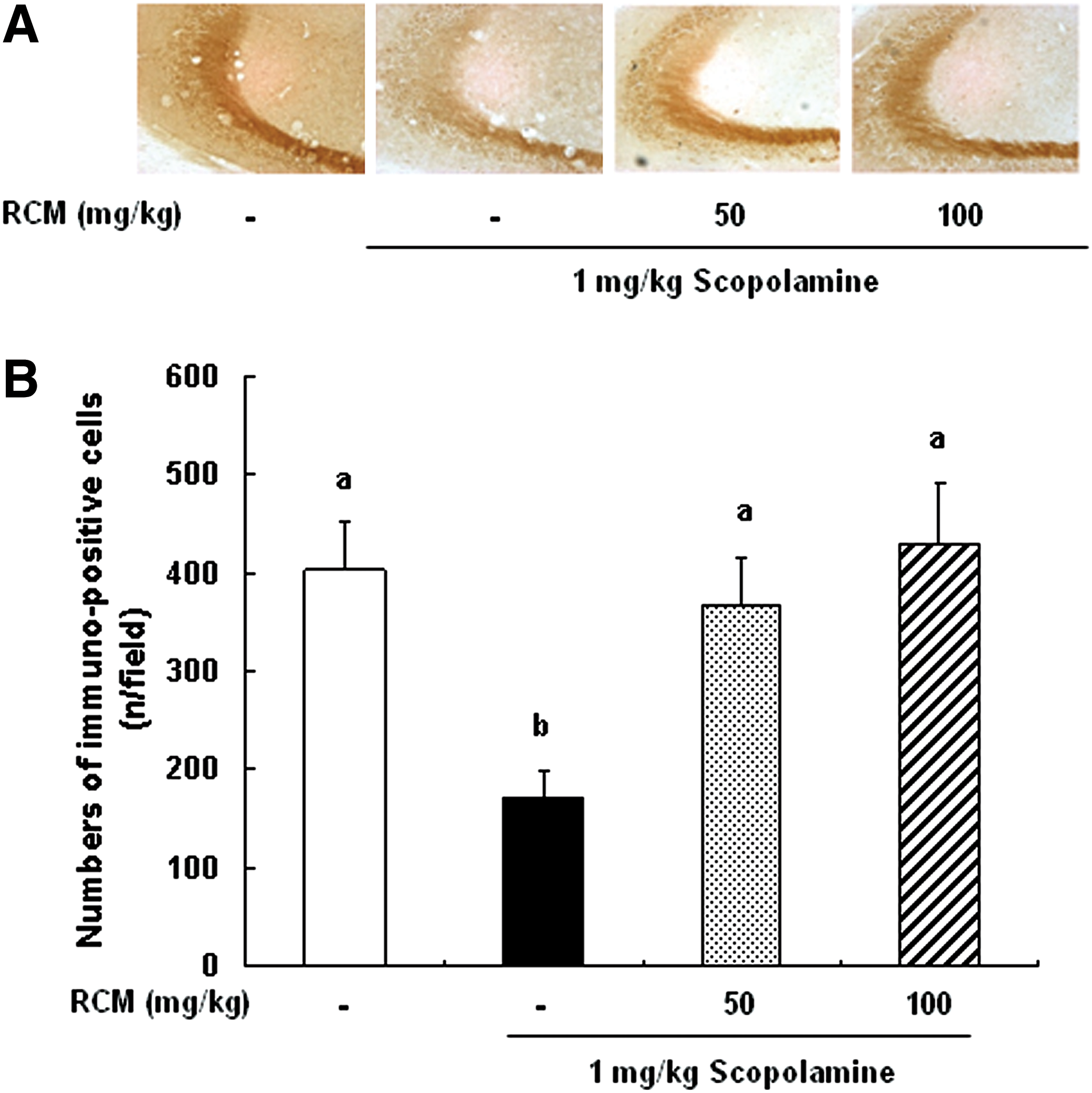
Effect of RCM on the phosphorylation of extracellular signal-regulated kinase (ERK) in the brain CA3 region. The mice were subjected to oral gavage with RCM and memory deficit was induced with scopolamine as described in Figure 1. The mice were perfused 30 min after scopolamine injection and the brains were removed. Brain tissues were stained with antibody raised against phospho-ERK, and then immunostained using the Elite ABC kit.
RCM administration exhibits antioxidant activity on scopolamine-induced oxidative stress
We measured the enzyme activities of SOD and GPx in the brains of mice. The treatment with scopolamine resulted in a significant decrease in SOD activity in the cerebral cortex and hippocampus. The administration of RCM significantly mitigated the decreases in SOD activities induced by scopolamine in the cerebral cortex and hippocampus (Table 3). The GPx activities were also significantly decreased by the treatment of scopolamine in the cerebral cortex and hippocampus. The decreased GPx activities induced by scopolamine were restored by the treatment with RCM in the cerebral cortex and hippocampus (Table 3).
The mice were subjected to oral gavage with RCM for 4 weeks and induced memory deficit with scopolamine as described in Figure 1. The mice were euthanized 30 min after scopolamine injection. The brains were removed. The activities of SOD and GPx in the cerebral cortex and hippocampus of brain were estimated using the relevant assay kit. Values represent the mean±SEM (n=5). Means without a common letter differ, P<.05.
SOD, superoxide dismutase; GPx, glutathione peroxidase.
Discussion
Convincing evidence suggests that RCM exhibits various beneficial health effects. 13 –17 However, to the best of our knowledge, the effects of RCM on memory have yet to be assessed. In the current study, we investigated whether RCM protects against memory impairment in mice. The results presented here demonstrate that the oral administration of RCM improves scopolamine-induced memory deficit. As RCM has been demonstrated to potently improve memory impairment in this animal model, future studies will be necessary to determine whether RCM effectively enhances memory in humans.
Presently, several medicines, such as tacrine and donepezil, have been used to treat AD patients and aged humans with memory deficit. 7 However, these medicines have been shown to cause some unwanted side effects. 8 Accordingly, it is important to find novel agents that are relatively safe and that can effectively improve memory. RCM used in this study was a lyophilized powder of a completely ripened fruit. This RCM is expected to show low or no toxicity or undesirable side effects because humans have a long history of consuming this completely ripened fruit. Therefore, this RCM may be a novel and safe candidate for the development of a memory-improving agent.
The cholinergic system plays a major role in the regulation of memory processes. The cholinergic deficit is a major feature that is associated with memory loss. Cholinergic agonists can facilitate memory, whereas cholinergic antagonists can impair memory. 20 Scopolamine reduces cholinergic function, thereby causing memory impairment. 9 In this study, we found that scopolamine significantly impaired the memory of mice, and that the oral administration of RCM for 28 successive days significantly reversed scopolamine-induced memory impairment in mice. These results suggest that RCM ameliorates scopolamine-induced memory impairment, which may function as cholinergic agonists.
Acetylcholine is considered to be one of the important neurotransmitters involved in the memory process. 20 It was reported that the hippocampal acetylcholine release was increased in the training of spatial memory performance. 21,22 The decrease of acetylcholine in specific brain regions resulted in memory deficit. 23 Acetylcholine is biosynthesized by ChAT, and is hydrolyzed by AChE. The expression and activation of AChE and ChAT regulate the dynamic concentration of acetylcholine in the cholinergic synapses in the brain, and then played a crucial role in the memory process. 5,24,25 AChE inhibitors 7,24 and ChAT activators 25 increase the acetylcholine levels in the brain, and hence reverse memory deficits. Many studies have reported that scopolamine increased AChE activity in the brain. 24 –26 In this study, we noted that scopolamine markedly increased AChE activities in the hippocampus and serum, and these increases were suppressed via the oral administration of RCM. Losses of ChAT immunoreactivity in scopolamine-induced memory-impaired mice were significantly restored by the RCM administration. In addition, the acetylcholine levels that were reduced by scopolamine treatment increased in the cortex, hippocampus, and serum. These results indicate that the memory-enhancing effect of RCM might be due to the inhibition of AChE and activation of ChAT, leading to increased levels of acetylcholine.
CREB, a transcription factor, functions as a molecular switch to control the consolidation of short-term memory into long-term memory. 27 Further, CREB appears to play a universal role in memory function. 28 There has been an increasing interest in investigating CREB's involvement under pathological conditions of memory impairment, in particular AD. 29,30 The activation of CREB by phosphorylation can be regulated by several kinases, including ERK 31 and protein kinases A 32 and B. 33 These signaling pathways play important roles in memory. 34 Several studies demonstrate that the CREB-mediated pathway in memory is one of the most attractive targets for the agents aimed at restoring and protecting memory abilities under pathological situations. 35,36 In the present study, we found that scopolamine reduced the phosphorylation of CREB and ERK in the hippocampus. The treatment with RCM significantly prevented the scopolamine-induced reduction of CREB and ERK phosphorylation. These results demonstrate that RCM exerts a memory-enhancing effect via the regulation of CREB and ERK activation. However, we did not determine the effect of RCM on the other pathway involving CREB and ERK upstream/downstream in this study. Therefore, future studies will be necessary to further establish the effect of RCM on the CREB-mediated pathway.
Clinical studies have reported strong evidence that oxidative stress is involved in the pathogenesis of AD. 37,38 Memory impairment in the scopolamine-induced animal model is associated with the altered status of brain oxidative stress. 39,40 Schroeter et al. 41 reported that supplementation with antioxidants delayed the development of AD and attenuated neuronal cell death induced by oxidative stress. RCM has been reported to exert a strong antioxidant ability. 15 Several studies showed that scopolamine inhibits the activities of SOD and GPx in the brain. 26,42 In this study, the treatment with scopolamine resulted in a significant decrease in both SOD and GPx activities in the cortex and hippocampus. The administration of RCM restored the activities of SOD and GPx in the cortex and hippocampus. Our results suggest that the memory-improving effect of RCM may be due in part to the antioxidant action of RCM. It has been reported that antioxidants attenuated oxidative-stress-induced neuronal cell death. Since RCM exhibited antioxidant activity via the increase of SOD and GPx activities, RCM may have the potential to protect against neurotoxicity. However, the present results did not address the question of whether RCM attenuates scopolamine-induced neurotoxic effects in the brain. To answer this question, additional experiments will be necessary to investigate whether scopolamine induces neurotoxicity and, if it does, whether RCM protects against scopolamine-induced neurotoxicity in the brain.
In conclusion, these results showed that the oral administration of RCM improves memory in scopolamine-induced memory-impaired mice. We further demonstrated that the memory-improving effects of RCM may be attributed to the inhibition of AChE activity, the increase of ChAT expression, the activation of CREB and ERK, as well as the antioxidant activity. These results indicate that RCM may prove to be a safe and effective agent for the improvement of memory.
Footnotes
Acknowledgments
This work was supported by the Ministry of Knowledge Economy through the Center for Efficacy Assessment and Development of Functional Foods and Drugs at Hallym University, Korea.
Author Disclosure Statement
The authors declare that they have no competing interests.