
Abstract
Acquired immunodeficiency syndrome patients treated with red ginseng, which contains protopanxadiol and protopanaxatriol ginsenosides as its main constituents, have been reported to remain healthy for >20 years in the absence of highly active antiretroviral therapy. Of these ginsenosides, ginsenoside Rh1, a protopanaxatriol ginsenoside, is known to eliminate cytoprotective HIV-1-infected macrophages by inhibiting pyruvate dehydrogenase lipoamide kinase isozyme 1 (PDK-1) phosphorylation. In this study, we investigated the capacity of ginsenoside Rb1, a protopanaxadiol ginsenoside, to eliminate cytoprotective primary human macrophages. We found that ginsenoside Rb1 could also eliminate cytoprotective primary human macrophages infected with HIV-1 D3. Ginsenoside Rb1 inhibited lipopolysaccharide/cycloheximide-induced AKT and glycogen synthase kinase-3β phosphorylation in the D3-transduced macrophages, but not the phosphorylation of PDK-1 and phosphoinositide-3-kinase (PI3K). Furthermore, we also observed that a combined treatment with ginsenoside Rb1 and miltefosine synergistically abolished the cytoprotective CHME5 cells expressing HIV-1 tat. Based on these findings, we can conclude that ginsenoside Rb1 can eliminate cytoprotective macrophages infected with HIV-1 by inhibiting the AKT pathway.
Introduction
H
Red ginseng (the steamed root of Panax ginseng C.A. Meyer, Araliaceae), which contains ginsenosides as its main bioactive constituent, is frequently used for tumor, diabetes, inflammation, and stress in Asian countries. 11,12 Recently, most AIDS patients treated with red ginseng remain healthy for >20 years in the absence of highly active antiretroviral therapy (HAART). 13,14 Of these ginsenosides, ginsenoside Rh1, a protopanaxatriol ginsenoside, eliminated the cytoprotective phenotype of HIV-1-infected macrophages by inhibiting pyruvate dehydrogenase lipoamide kinase isozyme 1 (PDK-1) phosphorylation. 15
Therefore, we investigated the ability of ginsenoside Rb1, a protopanaxadiol ginsenoside, to eliminate the cytoprotective phenotypes of Tat-expressing CHME5 cells and D3-transduced human primary macrophages.
Materials and Methods
Reagents
LPS purified from Escherichia coli O26:B6, cycloheximide (CHX), propidium iodide solution (PI), RPMI-1640 medium, polybrene, Trypan blue, fetal bovine serum (FBS), and Dulbecco's modified Eagle's medium (DMEM) were purchased from Sigma (St. Louis, MO, USA). CD14 magnetic beads and LS columns were purchased from Miltenyi Biotec (Bergisch Gladbach, Germany). Antibodies for PI3K, Akt, p-Akt, GSK-3β, p-GSK-3β, and β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies for p-PDK-1, PDK-1, mTOR, p-mTOR, Bcl-2-associated death promoter (BAD), and p-BAD were purchased from Cell Signaling Technology (Beverly, MA, USA). Polyvinylidene difluoride membranes (PVDF) and enhanced chemiluminescence detection kit (Luminta™ Forte Western HRP substrate) were purchased from Millipore (Billerica, MA, USA).
Ginsenoside Rb1 was isolated from red ginseng according to the previously reported method of Choo et al. 16
Cell line
CHME5, a human fetal microglial cell line, was obtained from Dr David Mock (University of Rochester, NY, USA). Lipofectamine 2000 was used to transfect these cells with the plasmid pTat101 to establish a CHME5 cell line that stably expresses full-length Tat. The transfected cells were cultured for 15 days in a medium supplemented with 1 mg/mL of hygromycin, and then hygromycin-resistant cells were selected clonally. The control cells were transfected with an empty vector (pcDNA3.1/Hygro; Invitrogen, Carlsbad, CA, USA) and subjected to drug selection as indicated above. The parental CHME5 cells were maintained in the DMEM containing 10% FBS, 15 whereas the stable subline was maintained in a medium supplemented with 500 μg/mL of hygromycin. The presence of the tat was confirmed by PCR amplification of the tat DNA sequences using gene-specific primers, and the presence of a functional gene was confirmed by the activation of viral LTRs.
The D3 vector pseudotyped with the VSV-G envelope protein was prepared by the method of Diamond et al. 17 The D3 vector system expresses eGFP and all HIV-1 viral proteins except the env and nef proteins.
Isolation of macrophages derived from primary human monocytes
Human monocytes were purified using the method of Jeong et al. 15 Briefly, peripheral blood mononuclear cells (PBMCs) from blood samples of voluntary male donors (age, 20–29 years) were harvested using Ficoll density gradients (Lymphoprep, Axis-Shield PoC AS, Oslo, Norway), and the monocytes were then purified from the PBMCs by immunomagnetic selection using magnetic beads conjugated with the anti-CD14 antibody (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer's instructions. The purified human monocytes were incubated in culture plates in the RPMI 1640 medium (Sigma) for 4 days in the presence of 5 ng/mL human recombinant GM-CSF (R&D Systems, Minneapolis, MN, USA) and then were incubated for an additional 3 days in the absence of GM-CSF to allow differentiation into macrophages.
Trypan blue staining assay
Primary human macrophages transduced with HIV-1 D3 and CHME5 cells expressing tat were subjected to chemical stress by treating them with 50 μg/mL LPS and 10 μg/mL CHX in the presence or absence of ginsenoside Rb1 (5, 10, 15, and 20 μM) for 48 h. The cells were then trypsinized, stained with Trypan blue solution, and assessed for viability.
PI/Flow cytometry assay
Primary human macrophages transduced with HIV-1 D3 (2×105 cells/well) were treated with 50 μg/mL LPS and 10 μg/mL CHX stress in the presence or absence of ginsenoside Rb1 (5, 10, or 20 μM) for 48 h. The cells were then trypsinized, stained with PI (1 μg/mL), and analyzed on a flow cytometer (Accuri Cytometers, Ann Arbor, MI, USA).
Immunoblot assay analysis
Primary human macrophages transduced with HIV-1 D3 (1×106 cells/well) were stimulated with LPS/CHX in the presence or absence of ginsenoside Rb1 and then lysed according to the method of Jeong et al. 15 The proteins in the supernatant of the lysate were separated by 6% to 12% SDS-PAGE and the bands were transferred onto PVDF. The membranes were blocked with 5% nonfat dried milk in 0.05% PBST and then probed with anti-PI3K, anti-p-AKT, anti-Akt, anti-p-GSK3β, anti-GSK3β, anti-p-PDK1, anti-mTOR, anti-p-mTOR, and anti-β-actin antibodies. After washing with PBST, the proteins were detected by incubation with HRP-conjugated secondary antibodies for 50 min. Bands were visualized using the HRP substrate (Millipore Corporation, Billerica, MA, USA).
Combined cytotoxic effects of ginsenoside Rb1 and miltefosine
The cytotoxic effects of cotreatment with ginsenoside Rb1 and miltefosine on the cytoprotective CHME5 cells expressing tat were evaluated by Trypan blue staining. The CHME5 cells were treated with 50 μg/mL LPS and 10 μg/mL CHX in the presence or absence of 10 μM ginsenoside Rb1 and miltefosine (0, 1, 2, 5, or 10 μM) or 5 μM miltefosine and ginsenoside Rb1 (0, 1, 2, 5, 10, or 20 μM) for 48 h. The cells were then trypsinized, stained with Trypan blue, and viability was assessed.
Statistical analysis
The data were calculated with mean values, and standard deviations (mean±SD) were determined from triplicate trials. Statistical significance of the results was evaluated by one-way ANOVA and differences were considered significant at P≤.05.
Results and Discussion
We measured the cytotoxic effect of ginsenoside Rb1, a representative protopanaxadiol ginsenoside, against cytoprotective CHME5 cells that express HIV-1 tat by Trypan blue staining. We observed that ginsenoside Rb1 abolished the cytoprotective effect of the HIV-1 tat-expressing CHME5 cells more effectively than ginsenoside Rh1 did in a previous report (Fig. 1A, B). 15 To confirm this cytotoxicity, we also investigated the effect of ginsenoside Rb1 on cytoprotective primary human macrophages transduced with HIV-1 D3 by Trypan blue staining and PI/flow cytometry. Ginsenoside Rb1, in the presence of LPS/CHX, exhibited cytotoxicity in a dose-dependent manner. However, in the absence of ginsenoside Rb1, LPS/CHX did not induce cell death in the CHME5 cells expressing tat as well as in the primary human macrophages transduced with D3 at the tested concentrations (<40 μM) (Fig. 1C, D). Flow cytometry results also showed that ginsenoside Rb1 significantly induced cell death in transduced macrophages treated with LPS/CHX (Fig. 2). Furthermore, ginsenoside Rb1 showed no cytotoxicity against primary human macrophages that were not transduced with D3. We observed that treatment of D3-transduced primary human macrophages with LPS/CHX induced PI3K/AKT phosphorylation. However, treatment with LPS/CHX in the presence of ginsenoside Rb1 suppressed LPS/CHX-induced AKT, mTOR, and GSK3β phosphorylation, but did not influence PI3K and PDK1 phosphorylation (Fig. 3).
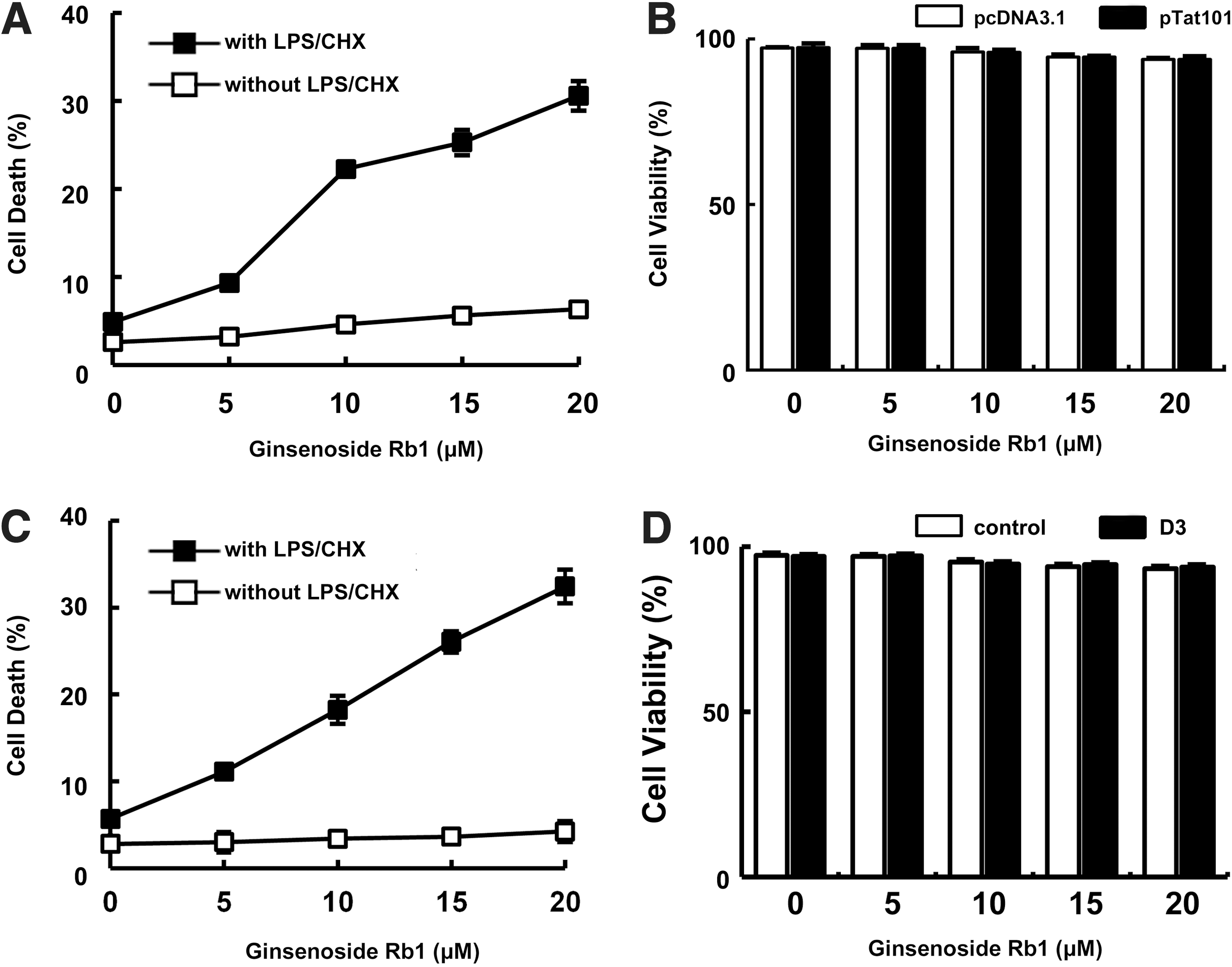
Cytotoxic effect of ginsenoside Rb1 against HIV-1 Tat-expressing cytoprotective CHME5 cells and D3-transduced human primary macrophages.
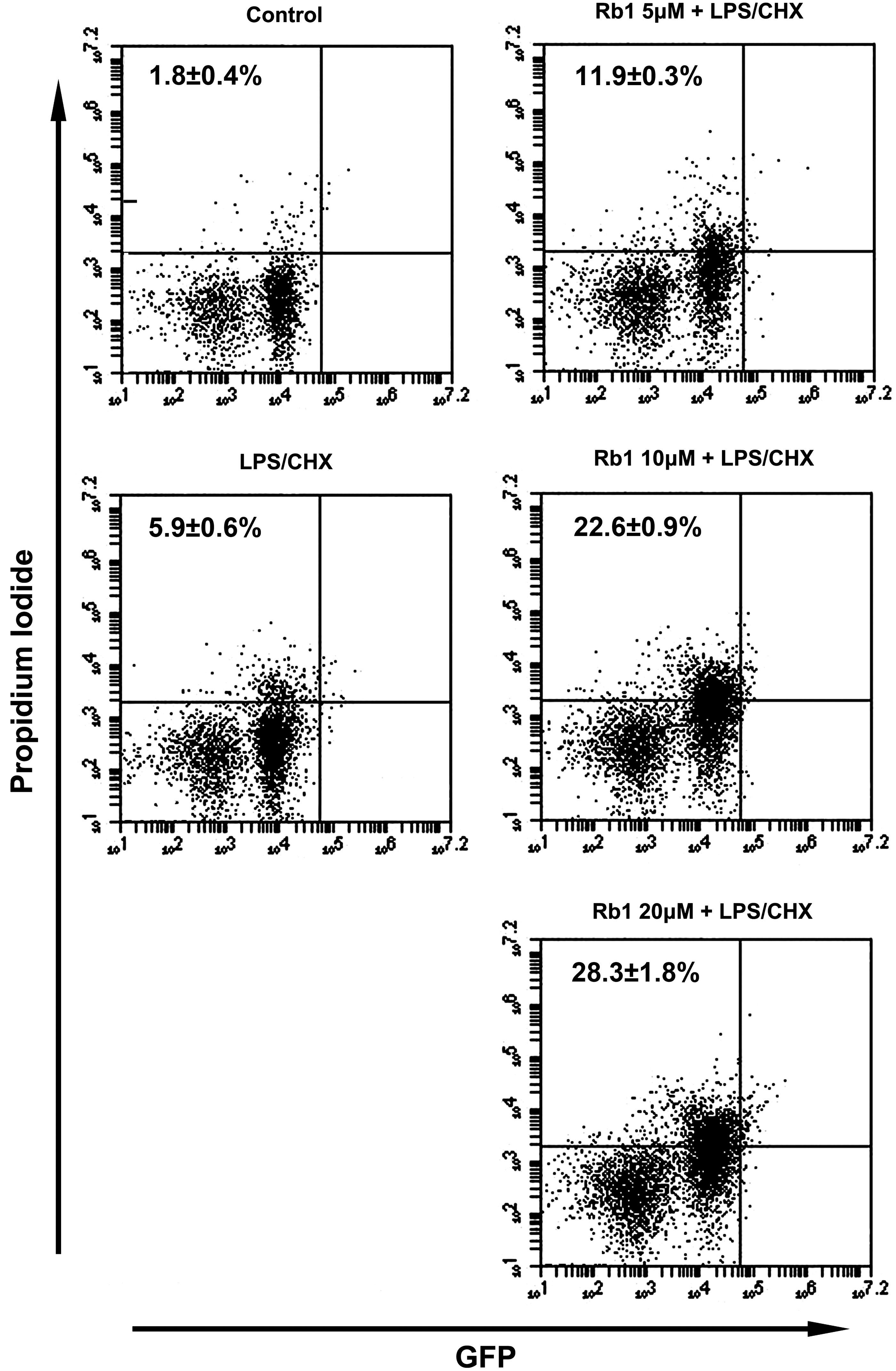
Cytotoxic effect of ginsenoside Rb1 against D3-transduced human primary macrophages using PI/flow cytometry assay. For the PI/flow cytometry assay, D3-transduced human macrophages were treated with and without 50 μg/mL LPS and 10 μg/mL cycloheximide (LPS/CHX) in the absence or presence of ginsenoside Rb1 for 48 h.
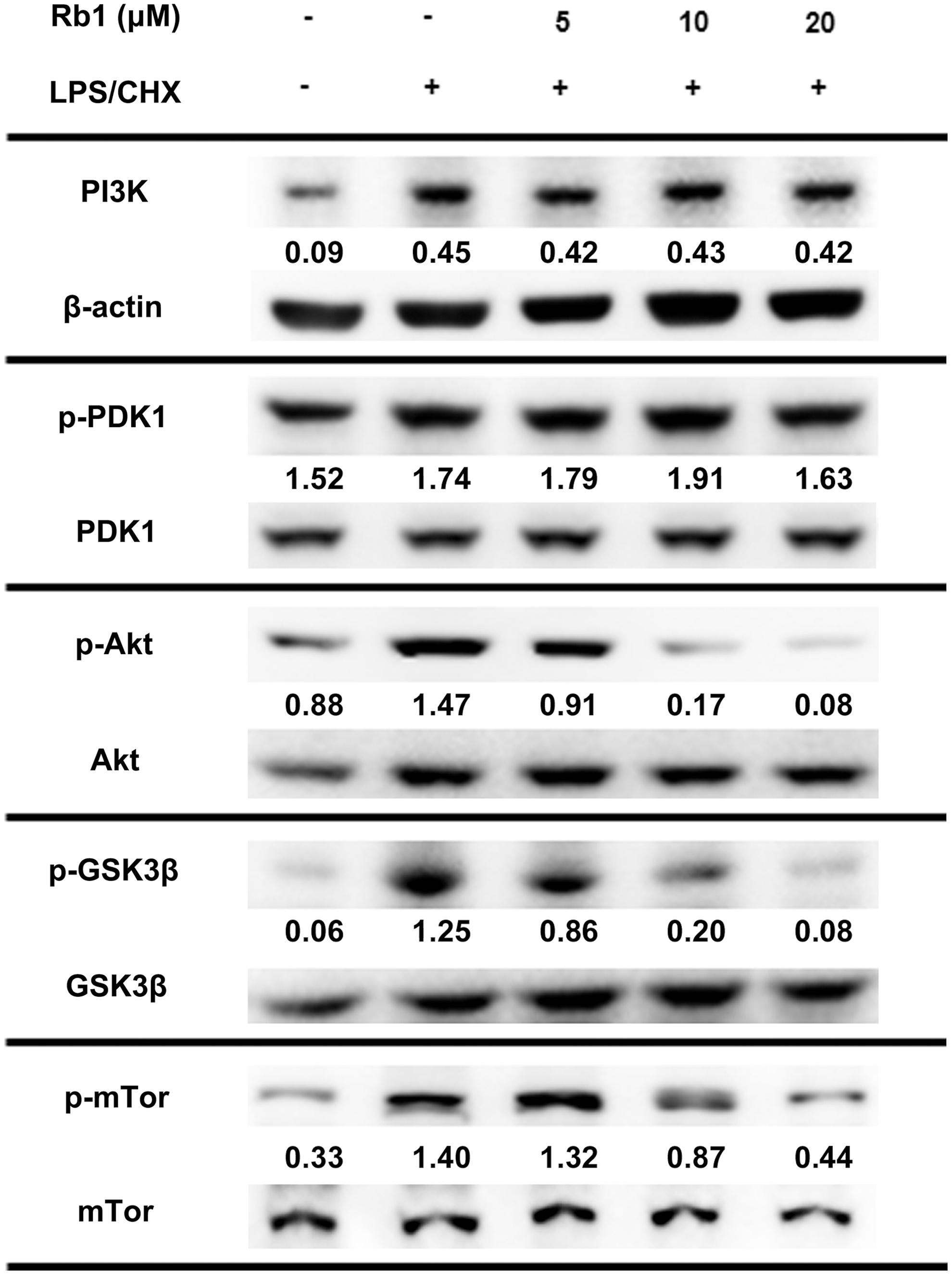
Effect on PI3K, AKT, and GSK3β phosphorylation in D3-transduced human primary macrophages using immunoblotting. Normal control was treated with vehicle alone. For the immunoblot assay, D3-transduced human primary macrophages were treated with and without LPS/CHX in the absence or presence of ginsenoside Rb1 (5, 10, and 20 μM) for 90 min. Immunoblotting for PI3K, p-PDK1, PDK1, p-Akt, Akt, p-GSK3β, GSK3β, mTOR, p-mTOR, and β-actin was performed for their lysates.
In humans, miltefosine eliminates HIV-1-infected cytoprotective macrophages by inhibiting the PI3K/Akt pathway, 1 but can cause severe side effects, such as nausea, vomiting, and teratotoxicity. 18,19 Therefore, to reduce the dose of miltefosine, we investigated the combined effect of miltefosine and ginsenoside Rb1 on the transduced CHME5 cells expressing tat. When cytoprotective Tat-transduced CHME5 cells were treated with various concentrations of ginsenoside Rb1 and a fixed concentration of miltefosine (5 μM), the cytotoxicity of ginsenoside Rb1 increased in a dose-dependent manner (Fig. 4). Similarly, the cytotoxicity of ginsenoside Rb1 (10 μM) also increased in a miltefosine dose-dependent manner. However, ginsenoside Rb1 (10 μM) in the presence of miltefosine showed no cytotoxicity against primary human macrophages that were not transduced with D3.
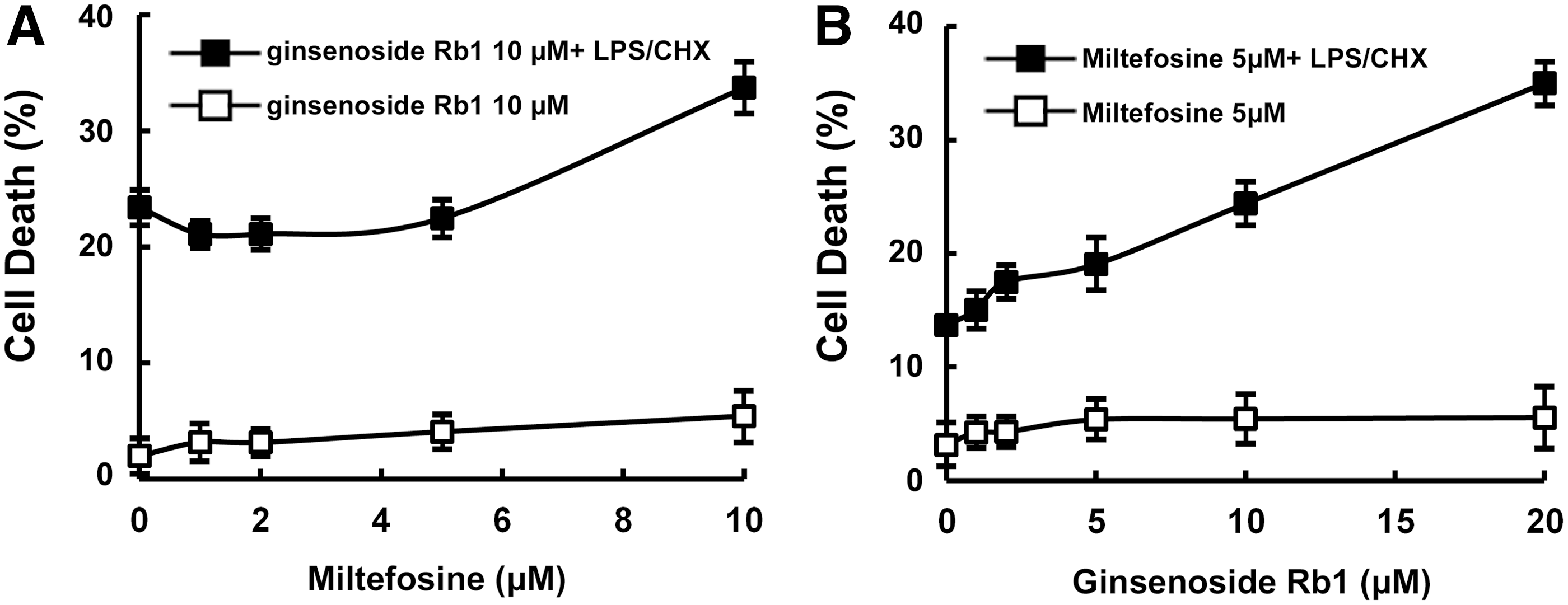
Combined cytotoxic effects of miltefosine and ginsenoside Rb1 against HIV-1 Tat-expressing cytoprotective CHME5 cells.
Summary
In the present study, we found that ginsenoside Rb1 eliminated the cytoprotective CHME5 cells expressing HIV-1 tat and primary human macrophages transduced with HIV-1 D3, which showed enhanced cell survival under LPS/CHX stress. 1,2 Furthermore, ginsenoside Rb1 potently inhibited HIV-1 tat- or D3-induced AKT pathways in CHME5 cells expressing HIV-1 tat and primary human macrophages transduced with HIV-1 D3. These results suggest that ginsenoside Rb1 eliminates cytoprotective HIV-1-infected macrophages in vivo through inhibition of the AKT pathway. The action mode of ginsenoside Rb1 was unique in that ginsenoside Rh1 abolishes cytoprotective tat-transduced CHME5 cells by inhibiting PDK-1 phosphorylation. In addition, ginsenoside Rb1 in combination with miltefosine, which is effective for HIV-1 dementia or Leishmania infection, 20 synergistically eliminated cytoprotective macrophages expressing HIV-1 tat. These results suggest that a combination therapy with miltefosine and ginsenosides may be more beneficial than miltefosine alone. These results, in part, agree with the finding of Cho et al. that most patients treated with red ginseng remain healthy for >20 years in the absence of HAART. 13 Taken together, these findings suggest that ginseng could be an effective complementary medicine for AIDS patients.
Finally, ginsenoside Rb1 may exert anti-HIV-1 effects by shortening the lifespan of HIV-1-infected macrophages and by protecting neurons in HIV-1-infected brains.
Footnotes
Acknowledgment
This study was supported by a grant from the World Class University Program through the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology (R33-2008-000-10018-0).
Author Disclosure Statement
No competing financial interests exist.