
Abstract
Xanthoceraside, a saponin extracted from the husks of Xanthoceras sorbifolia Bunge, suppresses inflammation and oxidative stress. However, the antitumor properties of xanthoceraside as well as its mechanism of action remain unclear. Therefore, we proposed to investigate its potential anticancer property. In this study, the viability of cells was measured by the MTT assay. Cell cycle and mitochondrial membrane potential were measured by flow cytometry, and the expressions of procaspase-9, procaspase-3, Cyto.c, Apaf-1, Bcl-2, Bcl-xL, Bad, p53, and IGF-1R/Raf/MEK/ERK were tested by Western blotting. Xanthoceraside significantly inhibited the proliferation of human melanoma A375.S2 cells in a concentration- and time-dependent manner but did not impair the viability of normal cells (peripheral blood mononuclear cells). Further analysis revealed that xanthoceraside induced apoptosis by activating caspase-3 and caspase-9 in a time-dependent manner through the mitochondrial pathway but did not activate caspase-8 in the cells. In addition, xanthoceraside inhibited the expression of the insulin-like growth factor-1 receptor (IGF-1R), which is an important prosurvival, antiapoptotic signaling growth factor receptor that is frequently overexpressed in cancer cells and used as a therapeutic target for multiple cancers. Interestingly, xanthoceraside also decreased the expression of Raf, p-MEK, and p-ERK, the downstream effectors of IGF-1R. Taken together, these findings indicate that xanthoceraside induces apoptosis through a mitochondria-mediated apoptotic pathway, which is induced by the downregulation of IGF-1R/Raf/MEK/ERK cascades in A375.S2 cells.
Introduction
X
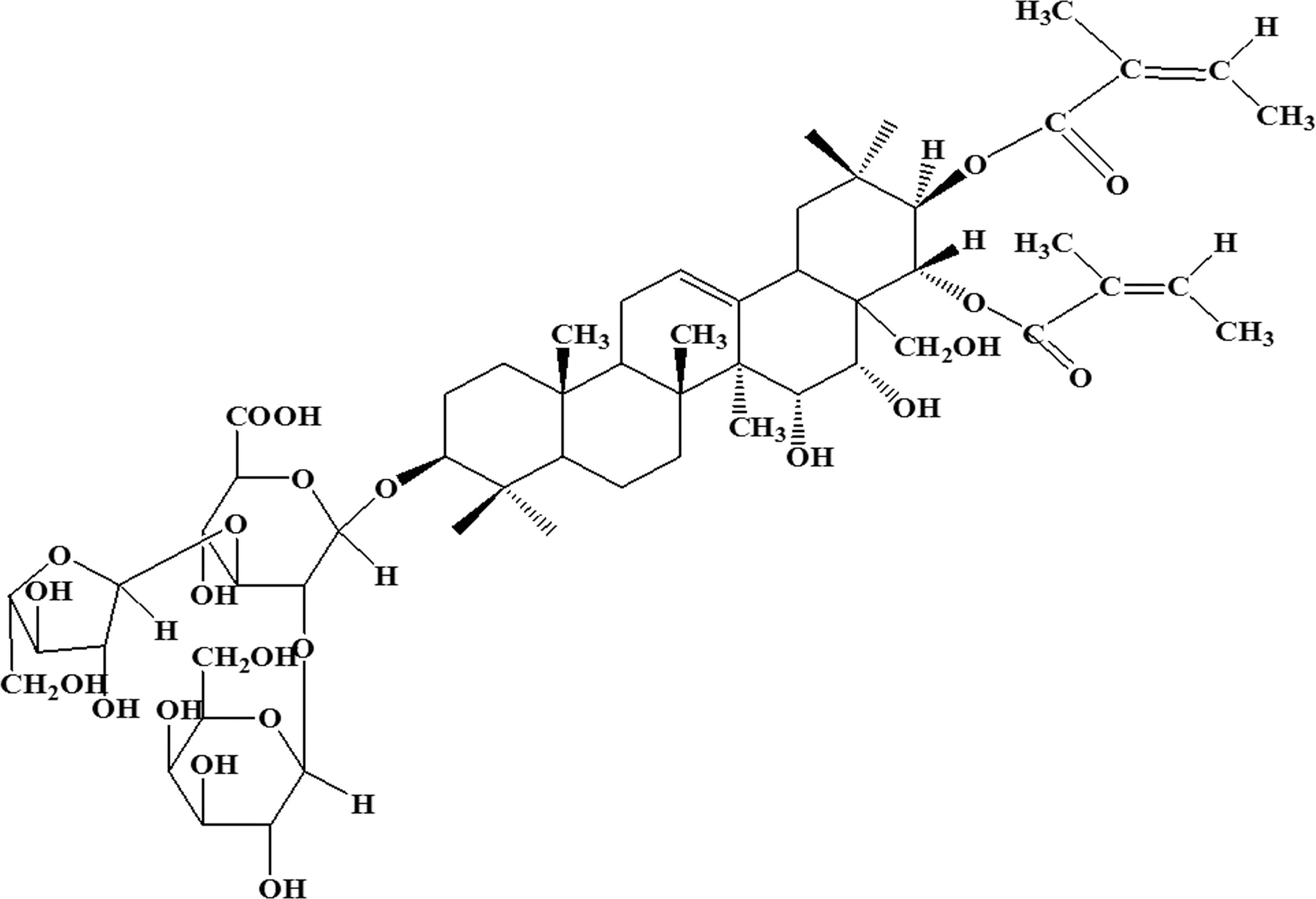
The chemical structure of xanthoceraside.
Melanoma is implicated in most skin cancer deaths, although it accounts for less than 5% of all the cases of skin cancer. It is the most aggressive type of skin cancer and the fifth and seventh most common cancer in men and women, respectively, in the United States. Among cancer patients under 40 years of age, the incidence of melanoma is second only to that of breast cancer for women and leukemia for men. 5 Despite years of research, the etiological mechanism of melanoma remains elusive although the insulin-like growth factor-1 receptor (IGF-1R) is considered to play a role.
In cancer cells, IGF-1R is a frequently overexpressed prosurvival, antiapoptotic signaling growth factor receptor, which has been reported as a potential therapeutic target for various types of cancer. 6 The mechanisms that downregulate or inhibit IGF-1R may contribute to the antitumor effect. 7,8 The IGF-1R signaling pathway mainly includes the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) pathway and the serine/threonine kinase (Raf)/MAP kinase kinase (MEK)/extracellular signal-regulated kinase (ERK) pathway. 9 –12 In the Raf/MEK/ERK pathway, activated Raf phosphorylates MEK, and MEK subsequently phosphorylates ERK 1/2 at its tyrosine and threonine sites. 13 ERK has many downstream targets, including transcription factors (such as NF-κB) as well as other proteins involved in cell cycle regulation. 14 The Raf/MEK/ERK pathway is mostly involved in cell growth modulation. 9,12 In melanomas, genes that express Raf proteins (ARAF, BRAF, and CRAF) are found to be mutated with BRAF mutations being most common with a rate from 50% to 70% among all the incidences. 15 Inhibition of BRAF results in cellular apoptosis and the mechanisms include the dephosphorylation of Bad, the translocation of Bmf, the upregulation of Bim, and the downregulation of Mcl-1. 16 –18
In the present study, we focus on the antitumor effects of xanthoceraside in human melanoma A375.S2 cells. Furthermore, we investigate the cellular and molecular mechanisms of xanthoceraside-induced apoptosis in A375.S2 cells.
Materials and Methods
Xanthoceraside and extraction procedure
Xanthoceraside was provided by the Department of Natural Products Chemistry, Shenyang Pharmaceutical University (Shenyang, China). The preparation procedure is described briefly: after grinding the husk of Xanthoceras Sobifolia into particles, the particles were extracted with solvents, passed through a macroporous resin, eluted by a solvent, and dried to yield total saponins. Then, the total saponins were dissolved in water, extracted by normal butyl alcohol, dried to obtain a brown powder, and chromatographed through a silica gel column. The gradient elution with l00:(35–60) chloroform:methanol was refined and recrystallized. White raphide was obtained as the xanthoceraside. 19
Materials
Xanthoceraside was dissolved in dimethyl sulfoxide (DMSO) and RPMI-1640 medium at a concentration of 1 mM and stored at −20°C. The final concentration of DMSO in all assays was not greater than 0.1%. 3-(4,5-Dimetrylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), propidium iodide (PI), and rhodamine 123 (Rho 123) were purchased from Sigma Chemical (St. Louis, MO, USA). The cell lysis buffer for Western blotting, the BCA protein assay kit, and enzymatic activity assay kits for caspase-3, caspase-8, and caspase-9 were obtained from Beyotime Biotechnology (Shanghai, China). The rabbit polyclonal antibodies against procaspase-9, procaspase-3, Cyto.c, Apaf-1, Bcl-2, Bcl-xL, Bad, p53, IGF-1R, Raf, p-MEK, MEK, p-ERK, ERK and the mouse polyclonal antibodies against β-actin and horseradish peroxidase-conjugated secondary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell lines and culture
Human melanoma A375.S2 cells were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were routinely cultured in the RPMI-1640 medium containing 10% fetal bovine serum, 100 U/mL of streptomycin, and 100 U/mL of penicillin in a humidified cell incubator with an atmosphere of 5% CO2 at 37°C. Fresh peripheral blood mononuclear cells (PBMC) were obtained from healthy volunteers after separation through Ficoll-Hypaque density sedimentation.
Cell viability assay
The viability of cells was measured by the MTT assay. The cells were planted in 96-well flat-bottom plates at a density of 1.5×104 cells per well. After 24 h in a CO2 incubator at 37°C, the cells were treated with various concentrations of xanthoceraside and cultured for 0, 6, 12, 18, and 24 h. Subsequently, 15 μL of MTT (5 mg/mL) was added to each well. Four hours later, the medium was removed and 150 μL of DMSO was added to each well. The absorbance was measured at 540 nm using an enzyme-linked immunosorbent assay (ELISA) reader (Thermo Fisher Scientific, Ashville, NC, USA). The percentage of cell viability was determined as follows:
cell viability (%)=[A540(xanthoceraside) − A540(blank)]/[A540(control) − A540(blank)]×100%.
Cell cycle analysis
A375.S2 cells were seeded in six-well plates at a density of 4.5×105 cells per well and incubated with 10 μM of xanthoceraside in the humidified cell incubator with an atmosphere of 5% CO2 at 37°C for 6, 12, 18, and 24 h. The collected cells were fixed with 500 μL of cold phosphate-buffered saline (PBS) and 10 mL of 70% ethanol at −20°C overnight, washed twice with PBS, and incubated with 1 mL of PI staining solution (PI 50 mg/L and RNase A 1 g/L) at 4°C for 30 min. The percentage of cells at different phases of the cell cycle or sub-G1 DNA content was measured by flow cytometry (Becton Dickinson, Franklin Lakes, NJ, USA), and the data were analyzed with multicycle DNA content and cell cycle analysis software. 20
DNA fragmentation assay
Fragmented DNA was isolated using a DNA extraction kit (Beyotime) according to the manufacturer's instructions. The eluants containing DNA pellets were electrophoresed in a 1.5% agarose gel at 80 V for 1.5 h. Digital images of the gel were obtained by an ultraviolet gel documentation system.
Enzymatic activity analysis
We quantified the enzymatic activity of caspase-3, caspase-8, and caspase-9 by commercial assay kits according to the manufacturer's protocols. Briefly, A375.S2 cells were lysed in a lysis buffer in an ice bath for 30 min. The lysates were centrifuged at 20,000 g for 10 min, and 30 μg/10 μL of protein was incubated with 80 μL of reaction buffer and 10 μL of colorimetric tetrapeptides, Ac-DEVD-pNA (acetyl-Asp-Glu-Val-Asp p-nitroanilide) for caspase-3, Ac-IETD-pNA (acetyl-Ile-Glu-Thr-Asp p-nitroanilide) for caspase-8, and Ac-LEHD-pNA (acetyl-Leu-Glu-His-Asp p-nitroanilide) for caspase-9, at 37°C for 2 h. The optical density of the reaction mixture was measured at 405 nm using an ELISA reader (Thermo Fisher Scientific).
Flow cytometric analysis with Rho123
The mitochondrial membrane potential (MMP) was measured by flow cytometry using the cationic lipophilic green fluorochrome Rho123. A375.S2 cells were harvested, washed twice with PBS, incubated with 1 μM Rho123 at 37°C for 30 min, and washed twice with PBS. The intensity of fluorescence was determined by flow cytometry with an excitation wavelength of 480 nm with an FL-1 filter (Becton Dickinson).
Western blotting
As reported previously, 13 the protein levels were quantified and resolved (30 μg) in 10% or 12% polyacrylamide gels and subsequently transferred to PVDF membranes by electroblotting. The membranes were blotted with primary antibodies followed by the horseradish peroxidase-conjugated secondary antibody. The blots were visualized using an enhanced chemiluminescence kit. The intensity of each band was normalized to β-actin.
Statistical analysis
All the presented data and results were confirmed in at least three independent experiments. All the data are expressed as the mean±standard deviation. The Western blot data were analyzed with Quantity One 4.6.2. Statistical comparisons were performed using one-way analysis of variance followed by Fisher's Least Significant Difference (LSD) multiple comparisons, where appropriate, using SPSS software. A P value less than .05 was considered to be statistically significant.
Results
Effects of xanthoceraside on A375.S2 cell viability
The effect of xanthoceraside on cell viability was examined by the MTT assay. With concentrations ranging from 4 to 10 μM and duration from 6 to 24 h, xanthoceraside inhibited the growth of A375.S2 cells in a concentration- and time-dependent manner (Fig. 2A) and the IC50 was 5.71 μM at 24 h, but did not obviously impair the viability of normal PBMC (P=.31) (Fig. 2B), which indicated its selective cytotoxicity against tumor cells. Based on the results of the MTT assay, we found that 10 μM xanthoceraside significantly inhibited the growth of A375.S2 cells at all time points. Thus, in the following experiments, a single dose of 10 μM was used for different time points.

Effects of xanthoceraside on the cell cycle in A375.S2 cells
Cell cycle and apoptosis were further evaluated by determining the number of cells in the sub-G1 phase. The A375.S2 cells were treated with 10 μM xanthoceraside for 0, 6, 12, 18, and 24 h. As shown in Figure 3, compared with the control group, xanthoceraside treatment from 12 to 24 h, increased the percentage of cells in the sub-G1 phase from 15.01%±2.91% to 30.67%±4.86% and 42.07%±2.81%, respectively. This arrest at sub-G1 phase suggested that xanthoceraside induced apoptotic cell death in the cells in a time-dependent manner.

Effects of xanthoceraside on the cell cycle in A375.S2 cells. The cells were treated with 10 μM xanthoceraside for 6, 12, 18, and 24 h, and the DNA content was analyzed by fluorescence flow cytometry after PI staining (mean±SD, n=3). PI, propidium iodide.
Effects of xanthoceraside on DNA fragmentation in A375.S2 cells
To determine whether xanthoceraside treatment induced DNA fragmentation, DNA was isolated from treated and untreated A375.S2 cells and separated by agarose gel electrophoresis. A375.S2 cells were treated with 10 μM xanthoceraside for 0, 6, 9, 12, 18, and 24 h. DNA fragmentation was observed at 24 h, which was indicated by the appearance of a typical ladder pattern at ∼180 bp intervals (Fig. 4, lane 7), whereas in untreated cells, no DNA fragmentation was observed (Fig. 4, lane 2). These results reconfirmed that xanthoceraside induced apoptosis in A375.S2 cells.
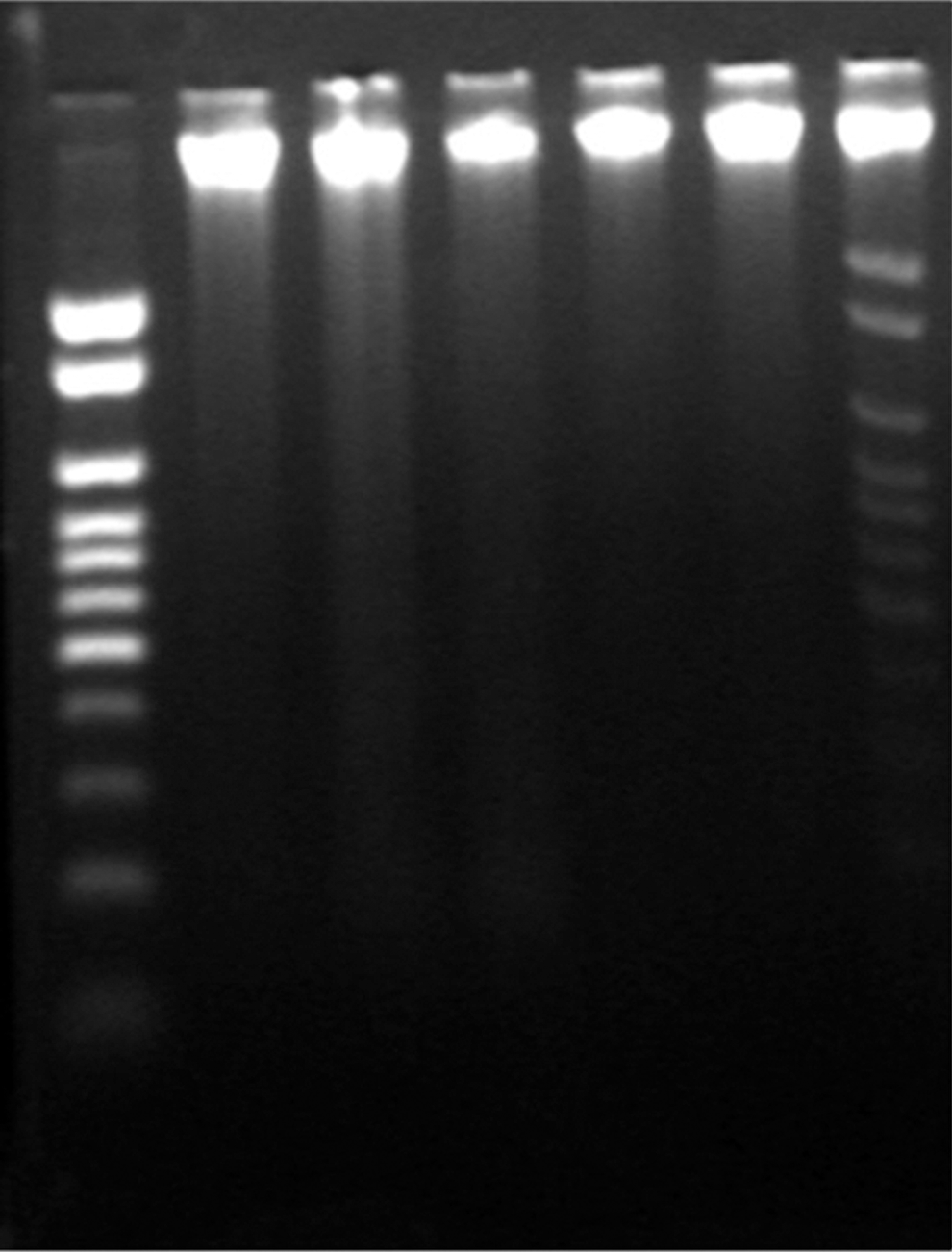
Effects of xanthoceraside on DNA fragmentation. A375.S2 cells were treated with 10 μM xanthoceraside for 0, 6, 9, 12, 18, and 24 h. The fragmented DNA was observed by agarose gel electrophoresis.
Effects of xanthoceraside on the activation of caspase-3, caspase-8, and caspase-9
To elucidate the molecular events involved in xanthoceraside-induced apoptotic cell death in A375.S2 cells, the levels of caspase-3, caspase-8, and caspase-9 were measured by using the enzymatic activity analysis. To quantify the activities of caspase-3, caspase-8, and caspase-9, lysates were equalized with a protein from the cells that were treated with xanthoceraside using DEVD-pNA, IETD-pNA, and LEHD-pNA, respectively, as the fluorogenic substrates. Xanthoceraside treatment significantly activated caspase-3 and caspase-9 in a time-dependent manner (P<.01). However, caspase-8 was not activated by the xanthoceraside treatment (6 h: P=.40, 12 h: P=.38, 24 h: P=.45) (Fig. 5A). Xanthoceraside significantly decreased the levels of procaspase-9 and procaspase-3 from 6 to 24 h (Fig. 5B). In addition, we found that xanthoceraside treatment increased the levels of p53, which is a typical marker of apoptosis at 12 and 24 h (P<.01) (Fig. 5B).
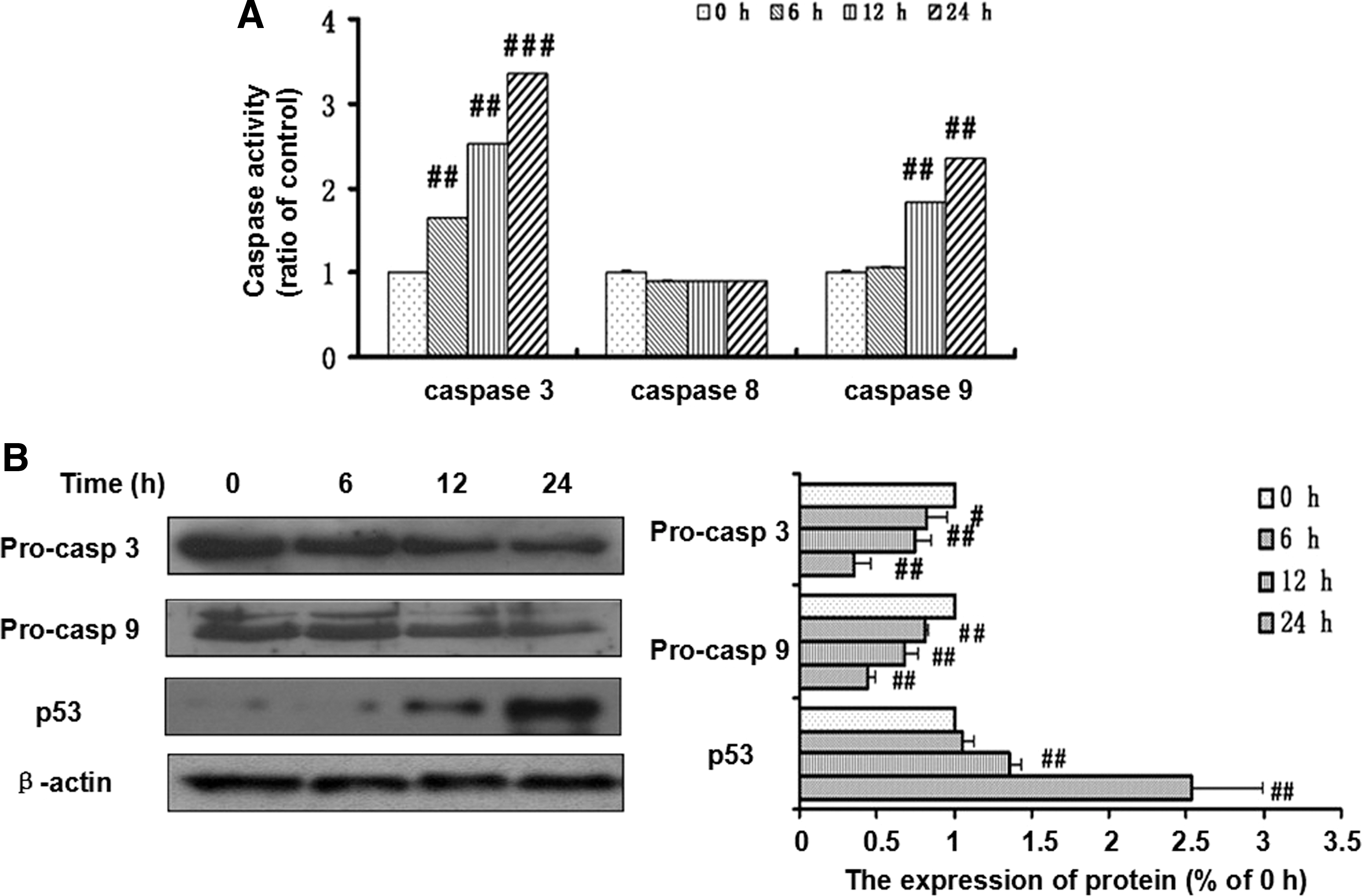
Effects of xanthoceraside on the activation of caspase-3, caspase-8, and caspase-9 in A375.S2 cells.
Effects of xanthoceraside on the mitochondrial pathway in A375.S2 cells
Because of the activation of caspase-9 with xanthoceraside treatment, we believed that the mitochondrial pathway might be involved in xanthoceraside induced apoptosis. Therefore, we examined the MMP using the Rho 123 probe. As shown in Figure 6, 10 μM xanthoceraside induced a marked decrease of the MMP, as determined by the flow cytometric analysis. Because the Bcl-2 family members play a vital role in the regulation of the mitochondrial apoptotic pathway, the levels of Bcl-2, Bcl-xL and Bad were detected by Western blot analysis. The expression level of Bad was significantly increased at 24 h, but the expression levels of Bcl-2 and Bcl-xL did not change after xanthoceraside treatment (Bcl-2—6 h: P=.79, 12 h: P=.49, 24 h: P=.54; Bcl-xL—6 h: P=.81, 12 h: P=.27, 24 h: P=.47) (Fig. 7). Simultaneously, the amount of cytochrome c (Cyto. c) in the cytosol of the cells was significantly increased, suggesting that Cyto.c was released from the mitochondria. Xanthoceraside also increased the level of Apaf-1 at 24 h (P<.01). These results clearly indicated that xanthoceraside-induced apoptosis in A375.S2 cells was mediated by the mitochondrial pathway.
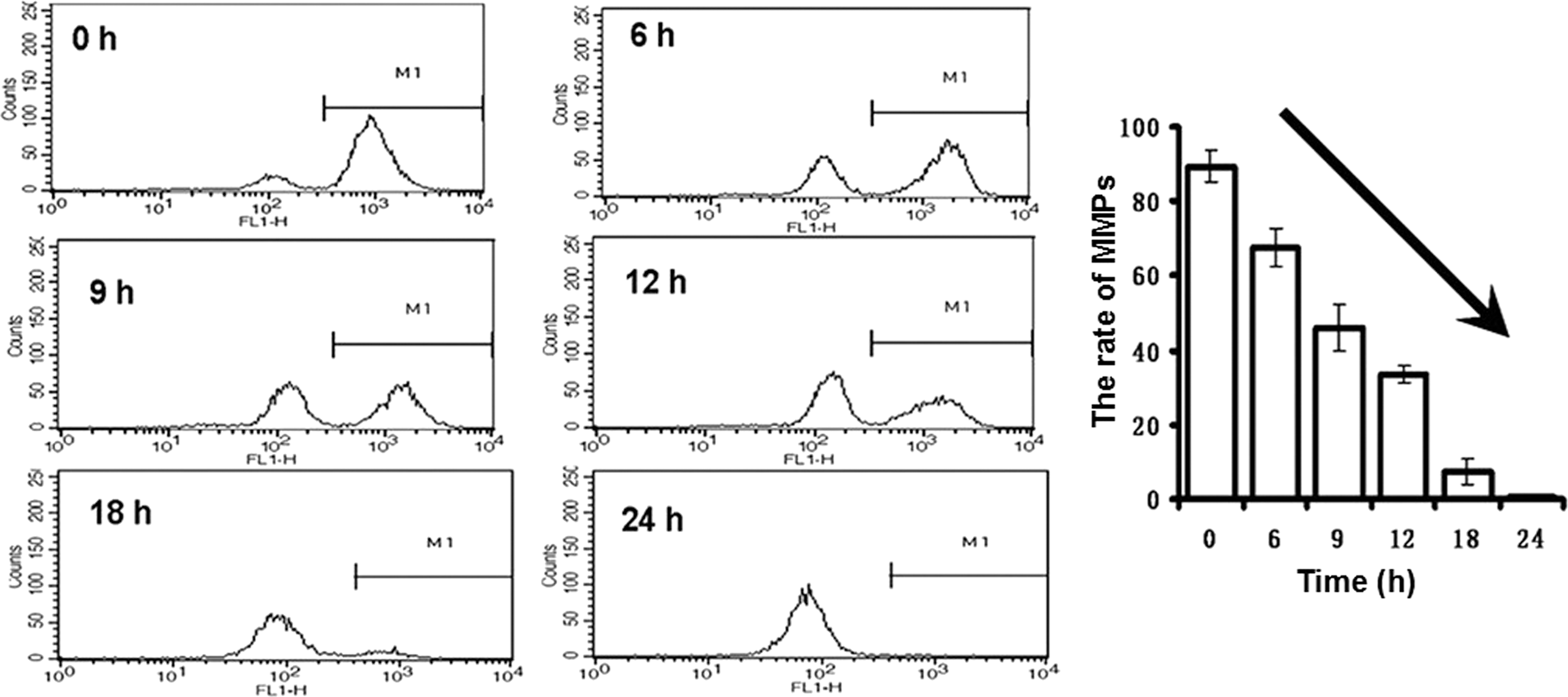
Xanthoceraside induces apoptosis through a mitochondria-mediated pathway. Cells were stained with 1 μM Rho123 to detect the change of MMP using flow cytometry (mean±SD, n=3). MMP, mitochondrial membrane potential.
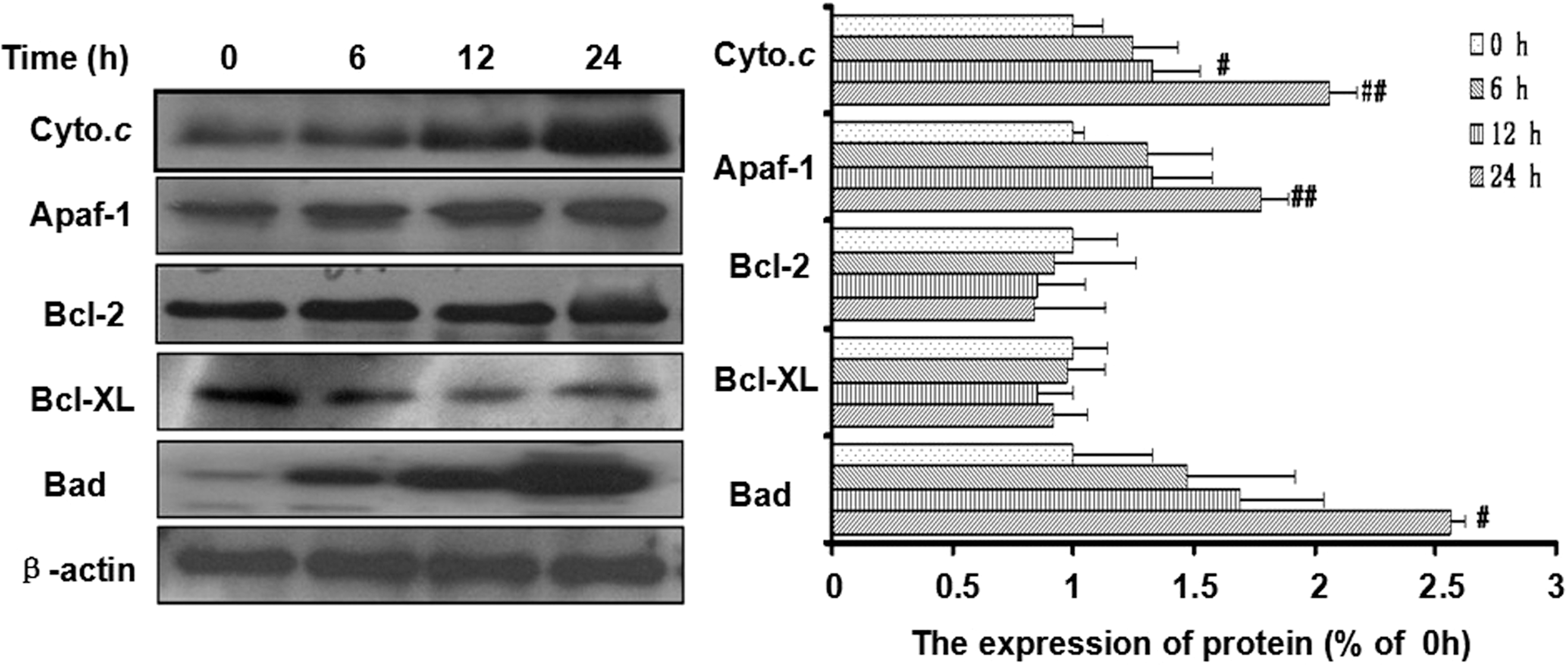
Xanthoceraside induces apoptosis through a mitochondria-mediated pathway. Xanthoceraside (10 μM) significantly increased the levels of Cyto.c (in the cytosol) in a time-dependent manner and the levels of Apaf-1 and Bad at 24 h, but xanthoceraside did not change the levels of Bcl-2 and Bcl-xL, as determined by Western blotting. The protein levels were analyzed by Quantity One 4.6.2 (mean±SD, n=3). # P<.05 and ## P<.01 when compared with 0 h.
Effects of xanthoceraside on the IGF-1R/RAF/MEK/ERK signaling pathway in A375.S2 cells
In melanoma, the activation of the IGF-1R/RAF/MEK/ERK signal transduction pathway contributes to tumor progression. Therefore, we investigated the effect of xanthoceraside on the IGF-1R/RAF/MEK/ERK signaling pathway in A375.S2 cells. Western blot analysis showed that xanthoceraside treatment significantly decreased the expression levels of IGF-1R and Raf at 12 and 24 h (P<.01). Xanthoceraside also significantly decreased the phosphorylation levels of MEK at 24 h and ERK in a time-dependent manner (p-MEK: P<.01; p-ERK: P<.001), but did not change the levels of total MEK and ERK (MEK—6 h: P=.93, 12 h: P=.91, 24 h: P=.97; ERK—6 h: P=.76, 12 h: P=.74, 24 h: P=.99) (Fig. 8). These results indicated that the IGF-1R/RAF/MEK/ERK pathway might be involved in the xanthoceraside-induced apoptosis.
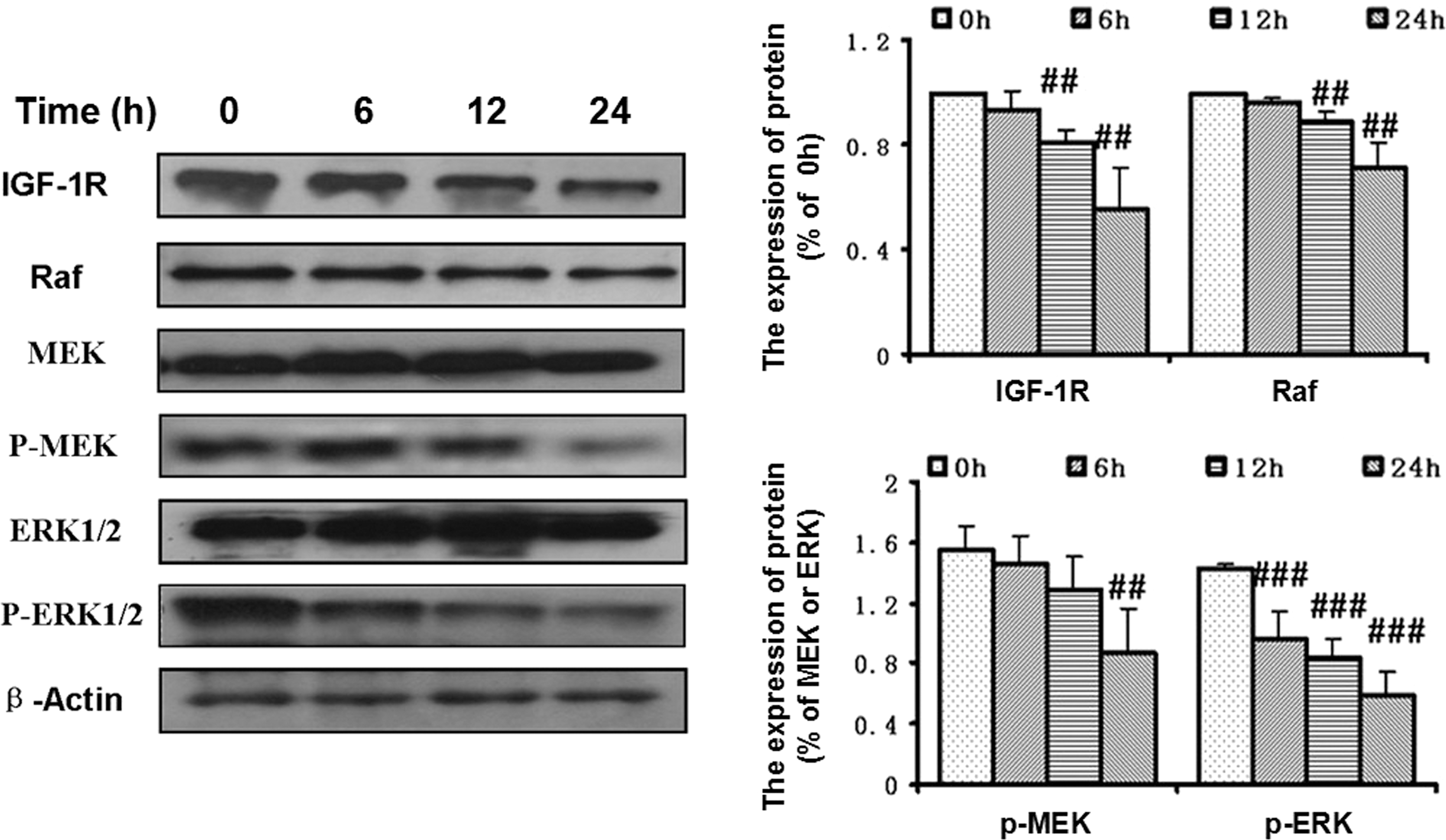
Xanthoceraside inhibits the IGF-1R-RAF-MEK-ERK signaling pathway. Protein levels of IGF-1R, Raf, MEK, p-MEK, ERK, p-ERK, and β-actin were analyzed by Western blot using specific antibodies. Xanthoceraside (10 μM) significantly decreased the expression levels of IGF-1R and Raf and the phosphorylation levels of p-MEK and p-ERK. The protein levels were analyzed by Quantity One 4.6.2 (mean±SD, n=3). ## P<.01 and ### P<.001. IGF-1R, insulin-like growth factor-1 receptor.
Discussion
Xanthoceraside shows anti-inflammatory and antioxidant activities. 3,21 Recently, antitumor effects of xanthoceraside have been revealed. Chan investigated the cytotoxicity of xanthoceraside against several kinds of human cancer cell lines and the IC50 values ranged from 14.5±1 to 130±14 μg/mL. 4 Ji et al. showed the antiproliferative effect of xanthoceraside on human breast cancer MCF-7 cells. 22 In this study, we assessed the cytotoxic effect of xanthoceraside on human melanoma A375.S2 cells. We found that xanthoceraside inhibited the growth of the A375.S2 cells, thereby indicating its antitumor potential. Interestingly, the viability of normal PBMC was less affected even at the highest concentration of xanthoceraside, 10 μM, which was highly toxic to the tumor cells, suggesting its selective cytotoxicity against tumor cells. We found that xanthoceraside significantly inhibited tumor growth in H22 tumor-bearing mice (Qing Jiao, Jingzhuo Tian, Jinjin Pan, Qian Xu, and Libo Zou, unpublished data), which indicated that xanthoceraside was biologically available. However, the pharmacokinetics and metabolism of xanthoceraside are still unknown. Therefore, further pharmacokinetics studies will be needed to identify the bioavailability and metabolites of xanthoceraside to fully understand its therapeutic potential.
Apoptosis is an essential process for both the development and maintenance of tissue homeostasis as first described by Kerr et al. 23 Inefficient apoptosis is considered as one of the most important hallmarks of tumorigenicity. 24 Therefore, induction of apoptosis plays a key role in cancer therapy. DNA fragmentation and ladder formation, one hallmark of apoptosis was clearly observed from our experiments. Xanthoceraside also increased the percentage of cells in the sub-G1 phase. All these results confirm that xanthoceraside induced apoptosis in A375.S2 cells. It has been well established that apoptosis in cells is induced through the activation of either the mitochondrial (intrinsic) pathway or the death receptor (extrinsic) pathway. 25 Caspase-8 is the initiator caspase of the extrinsic pathway. Fas-associated protein with death domain (FADD) recruits caspase-8 through homotypic interactions of death effector domains and results in caspase-8 activation and further activated the executioner caspases. 26 Caspase-9 has been specifically linked to the mitochondrial pathway. It is activated during the progression of Cyto.c release from mitochondria and the activation of Apaf-1. Our enzymatic activity analysis results imply that xanthoceraside activated caspase-3 and caspase-9 but not caspase-8, suggesting that xanthoceraside likely induced apoptosis through the mitochondrial pathway, but not through the death receptor pathways. The mitochondrial pathway is initiated by many intracellular stresses, such as radiation, oxidative stress, and nutrient deprivation. 27 Specifically, the discontinuity of the outer mitochondrial membrane results in the redistribution of Cyto.c to the cytosol followed by subsequent depolarization of the inner mitochondrial membrane. The release of Cyto.c further promotes caspase activation by binding to Apaf-1 and thus activates caspase-9. 28,29 The Bcl-2 family proteins, including the antiapoptotic Bcl-2 and Bcl-xL and the proapoptotic Bax and Bad, have effects on the stabilization of the MMP. 30 In our study, we found that xanthoceraside increased the expression of Bad, but did not change the expression of Bcl-2 and Bcl-xl, and caused a decrease in the MMP in the A375.S2 cells. The decrease in the MMP triggered the release of Cyto.c from the mitochondria to the cytoplasm, leading to the activation of effector caspases to promote apoptosis. p53, the guardian of the genome, is another main sensor in apoptosis. Activated p53 induces the expression of multiple transcriptional targets, including proapoptotic members of the Bcl-2 family. 31 We found that xanthoceraside treatment significantly upregulated the expression of p53. These results clearly demonstrate that xanthoceraside induces apoptosis through the mitochondrial pathway in A375.S2 cells.
IGF-1R plays an important role in tumor progression, resistance to apoptosis, and resistance to antitumor agents. 32 IGF-1R can activate the MAPK pathways and is frequently overexpressed in human melanoma cells. To further explore the apoptotic mechanism triggered by xanthoceraside, we attempted to identify the effect of xanthoceraside on the expression of IGF-1R in A375.S2 cells. Western blot analysis showed that 10 μM xanthoceraside treatment decreased the expression of IGF-1R in a time-dependent manner. The Ras/Raf/MEK/ERK is the main downstream signaling pathway of IGF-1R and modulates the function of numerous substrates involved in the regulation of cell survival, proliferation, and invasion. In human melanoma, the Ras/Raf/MEK/ERK signaling pathway is constitutively activated through multiple mechanisms. In humans, BRAF, a gene that encodes Raf protein, is mutated in 50% to 70% of all melanomas. 33 In 2011, a therapeutic agent for BRAF V600E mutation-positive metastatic melanoma, PLX4032 (Zelboraf®), inhibits the MEK/ERK pathway, and cell growth in BRAF V600E melanoma was approved. 34 These findings have focused efforts on developing inhibitors to target the RAF/MEK/ERK pathway. ERK1/2 regulates the cellular activity by acting on more than 100 substrates in the cytoplasm and nucleus, including transcription factors (such as NF-κB) and cell cycle kinases. 35 Investigators have reported that ERK1/2 activation lead to Bcl-2 transcriptional upregulation and Bcl-2 downregulation and also inhibited p-ERK1/2. 36,37 In this study, we observed that 10 μM xanthoceraside treatment decreased the expression of Raf, p-MEK, and p-ERK in a time-dependent manner and had no effect on MEK and ERK in A375.S2 cells. Xanthoceraside did not change the expression of Bcl-2, therefore we confirmed that there was no cooperative effect of ERK1/2 and Bcl-2 on xanthoceraside cytotoxicity. These results suggest that the positive effect of xanthoceraside on induction of apoptosis in A375.S2 cells may be associated with the suppression of the IGF-1R/Raf/MEK/ERK signaling pathways.
In conclusion, this is the first study to clearly demonstrate the effect of xanthoceraside to induce apoptosis in A375.S2 cells. These findings also indicate that xanthoceraside activates the caspase-dependent apoptosis through the mitochondria-mediated apoptotic pathway. This apoptosis is induced by the downregulation of IGF-1R/Raf/MEK/ERK cascades (Fig. 9). The effects of xanthoceraside on A375.S2 cells reported in this study warrant further investigations on xanthoceraside for the treatment of melanoma.
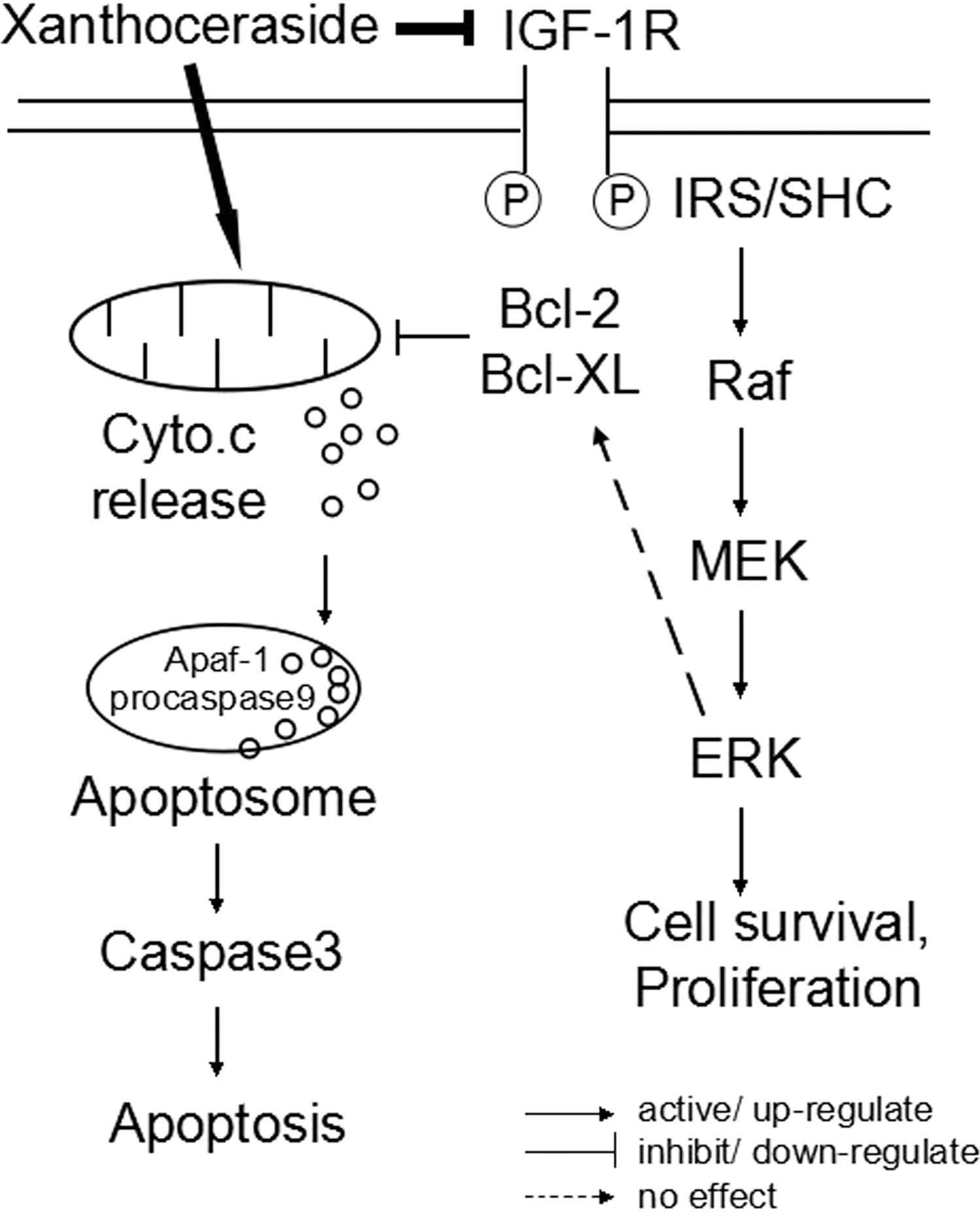
The proposed signaling cascade and the potential action of xanthoceraside.
Footnotes
Acknowledgment
The research described in this article was partially supported by the Foundation for the Research of Science and Technology of the Department of Education of Liaoning province (No. L2010526).
Author Disclosure Statement
No competing financial interests exist.