
Abstract
Ganoderma applanatum (GA) and related fungal species have been used for over 2000 years in China to prevent and treat various human diseases. However, there is no critical research evaluating the functionality of GA grown using submerged culture technology. This study aimed to evaluate the effects of submerged culture GA mycelium (GAM) and its active components (protocatechualdehyde [PCA]) on preadipocyte differentiation of 3T3-L1 cells. Mouse-derived preadipocyte 3T3-L1 cells were treated with differentiation inducers in the presence or absence of GAM extracts. We determined triglyceride accumulations, glycerol-3-phosphate dehydrogenase (GPDH) activities, and differentiation makers. PCA, the active component of GAM extract, was also used to treat 3T3-L1 cells. The MTT assay showed that the GAM extract (0.01–1 mg/mL) was not toxic to 3T3-L1 preadipocyte. Treatment of cells with GAM extracts and its active components significantly decreased the GPDH activity and lipid accumulation, a marker of adipogenesis, in a dose-dependent manner. Western blot analysis results showed that the protein expression levels of peroxisome proliferator-activated receptor γ (PPARγ), CCAAT/enhancer-binding protein α (C/EBPα), and sterol regulatory element-binding protein 1 (SREBP1) were inhibited by the GAM extract. In addition, adipogenic-specific genes such as perilipin, fatty acid synthase (FAS), fatty acid transport protein 1 (FATP1), and fatty acid-binding protein 4 (FABP4) decreased in a dose-dependent manner. Quantitative high-performance liquid chromatography analysis showed that the GAM extract contained 1.14 mg/g PCA. GAM extracts suppressed differentiation of 3T3-L1 preadipocytes, in part, through altered regulation of PPARγ, C/EBPα, and SREBP1. These results suggest that GAM extracts and PCA may suppress adipogenesis by inhibiting differentiation of preadipocytes.
Introduction
M
Obesity results from an imbalance between energy intake and expenditure and is mainly characterized at the cellular level by an increase in the size of adipocytes differentiated from preadipocytes, through the conversion of extra energy to lipids in adipose tissues. Researchers have investigated whether some food components exhibit the ability to suppress intracellular lipid accumulation.
3T3-L1 preadipocytes have been frequently used to study the differentiation of adipocytes in vitro. 3,4 As members of the CCAAT/enhancer-binding protein (C/EBP) family of transcription factors, C/EBPα, peroxisome proliferator-activated receptor γ (PPARγ), and sterol regulatory element-binding protein 1 (SREBP1) play critical roles in adiopogenesis. 5 During adipogenesis, C/EBPβ is rapidly induced by hormone signals and functions as a PPARγ transcriptional regulator. 6 PPARγ promotes the expression of a set of genes involved in the maturation of adipocytes. 7 Adipocyte differentiation and maturation are intimately related to the occurrence and development of obesity. 8 Adipocytes play a vital role in lipid homeostasis and energy balance by regulating triglyceride (TG) storage and free fatty acid release. Thus, many studies have been conducted to identify new bioactive compounds for obesity or weight control.
A number of compounds and extracts from natural sources exhibit the ability to suppress intracellular lipid accumulation. For example, tea catechins effectively inhibit adipogenesis in 3T3-L1 cells, 9 and in vivo studies have shown that oral administration of green tea decreases the weight of adipose tissue in rats and mice. 10,11 In addition, the same suppressive effects have been demonstrated with the mioga extract (Zingiber mioga Rosc.), 12 red yeast rice extract, 13 flavonoids, 14 and a mushroom extract 15 on 3T3-L1 and C3H10T1/2 B2C1 adipocyte differentiation.
Ganoderma applanatum (Pers.) Pat. (GA) (Order: Polyporales; Family: Ganodermataceae) and related species have been used for over 2000 years in China to prevent and treat various human diseases. 16 Many studies have reported that the major chemical constituents of GA and related mushroom species, such as polysaccharides, triterpenes, sterols, lectins, and some proteins, have beneficial properties for preventing and treating a variety of ailments. Remarkably, these ailments include very important diseases, such as bronchitis, hepatitis, hypertension, diabetes, tumor diseases, and immunological disorders. 17,18 In addition to its therapeutic effects, the terpenes isolated from GA also possess antioxidant capacity. 19
Artificial cultivation and a functional evaluation are required to utilize new mushroom species found in nature. Two representative methods for culturing mushrooms are solid state culture and submerged culture. In particular, submerged culture is an efficient method for producing polysaccharides and useful bioactive compounds from many mushrooms. 20 The influence of culture conditions on the submerged culture of mushroom has drawn much attention because the production of polysaccharides and triterpenoids from a culture broth containing mycelium is more efficient than that from fruiting bodies. 21 Several investigators have observed that both the culture medium and environmental conditions affect the production of useful components and also affect the physicochemical characteristics of the compounds. 22
In this study, GA was cultured under submerged conditions in complete media to elucidate the biological antiobesity activity of a liquid culture mycelium of the mushroom. The antiobesity effects of EtOH extracts were evaluated by confirming preadipocyte differentiation of 3T3-L1 cells in vitro, including a molecular level assay of relative enzymes.
Materials and Methods
Seed starter and culture conditions
The strain of GA was kindly provided by World Biotech Co. Ltd. (Kyungbuk, Korea). The stock culture was inoculated on a potato dextrose agar (PDA) slant and then incubated at 25°C for 10 days. The stock culture was maintained by monthly subculture and then the slants were stored at 4°C. 3T3-L1 fibroblasts were purchased from the Korean Cell Line Bank (KCBL, Seoul, Korea). GA was initially grown on a PDA medium and then transferred to a 50 mL seed culture medium (potato/dextrose broth) by punching out a portion (20 mm diameter). Fifty milliliters of the culture broth was aseptically homogenized at 9,809×g 3 min and inoculated 2% (v/v) into a culture medium with the following composition (g/L): glucose, 20; peptone, 2; yeast extract, 2; K2HPO4, 1; KH2PO4, 0.5; MgSO4, 0.5. The pH value was adjusted to 5 before sterilization. The submerged culture was carried out in a 5-L jar fermentor (Fermentec Co. Ltd., Cheongwon, Korea) containing 4 L of the medium at 25°C for 10 days.
Extraction of submerged culture mycelium
The total culture broth cultivated for 10 days was lyophilized and ground into powder. The powder, including mycelium, was extracted using 70% EtOH. The 70% EtOH (200 mL) was mixed with 10 g of powder and extracted with shaking at 25°C. The extract was filtered using filter paper (Advantec Co. Ltd., Tokyo, Japan) and then the residue was re-extracted three times. The filtrate was vacuum evaporated using a rotary vacuum evaporator (R-3000; Buchi Labortechik AG, Flawil, Switzerland). The GA mycelium (GAM) extract was freeze-dried, pulverized, and finally stored at −20°C until for analytical experiments.
Cell culture and differentiation
3T3-L1 preadipocytes were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% bovine calf serum (BCS) and 1% antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin) at 37°C in a 5% CO2 humidified incubator. Two days after confluence (day 0), the cells were induced to differentiate with a DMI mixture containing 1 mM dexamethasone, 0.5 mM 3-isobutyl-1-methyl-xanthine (IBMX), and 1 μg/mL insulin, in DMEM with 10% fetal bovine serum (FBS) for 2 days. The cells were then maintained in DMEM supplemented with 10% FBS and 1 μg/mL insulin for 2 days, which was refreshed every 2 days. During differentiation, the cells were treated with samples every other day for 8 days.
Cytotoxicity assay
Cytotoxicity was measured by a modified microculture tetrazolium (MTT) assay. 3T3-L1 preadipocytes were seeded in DMEM supplemented with 10% BCS into 96-well microtiter plates (1×104 cells per each well) and incubated at 37°C in a 5% CO2 incubator. After 24 h, vehicle or one of the five concentrations of samples (from 10 to 3000 μg/mL) in a serum-free DMEM was added to each well. After 24 h, 20 μL of MTT solution (5 mg/mL) was added to each well. Two hours later, the plate was centrifuged for 5 min at 221×g, the medium was then removed, and the resulting formazan crystals were dissolved with 150 μL DMSO. The optical density was measured at 570 nm using a microplate reader SpectraMax M2 (Molecular Devices, Sunnyvale, CA, USA).
Oil Red O staining
Adipocyte differentiation was estimated by staining the cultured cells with Oil Red O. Briefly, the cells were washed with phosphate-buffered saline (PBS) twice, fixed with 4% formaldehyde at room temperature for 20 min, and stained with Oil Red O for 30 min. Cells were then washed with 60% isopropanol three times to remove unbound dye and photographed. For quantification analysis, stained Oil Red O was eluted with isopropanol and quantified by measuring the optical absorbance at 510 nm.
Measurement of TG content
The cellular TG content was determined spectrophotometrically using a TG assay kit (Cleantech TG-S; Asan Pharm Co., Ltd., Whasung, Korea). Briefly, the cells were washed gently twice with PBS and lysed in a lysis buffer (1% Triton X-100 in PBS). Twenty microliters of the cellular lysate were mixed with 3 mL of the enzyme solution supplied and incubated for 10 min at 37°C. The absorbance at 550 nm was measured within 60 min. The protein concentration was determined by using a Bradford protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA).
Measurement of glycerol-3-phosphate dehydrogenase activity
The glycerol-3-phosphate dehydrogenase (GPDH)-specific activity was measured with a GPDH activity assay kit (Takara, Kyoto, Japan). Briefly, cells were carefully washed twice with ice-cold PBS on 8 days of the differentiation period, and an enzyme extraction buffer was added, and cells were collected by scraping with a cell scraper. After centrifugation at 14,240×g for 5 min at 4°C, the absorbance reduction of supernatant at 340 nm for 5 min was measured to estimate the rate of NADH oxidation during the GPDH-catalyzed reduction of dihydroxyacetone phosphate. One unit of enzyme activity corresponded to the oxidation of 1 nM NADH/min and the results were expressed as units per mg protein, and the protein concentration was determined by using a Bradford protein assay kit.
Western blot analysis
Cells were carefully washed twice with ice-cold PBS on 8 days of the differentiation period, solubilized in the lysis buffer containing 50 mM Tris-HCl (pH 7.5), 5% β-mercaptoethanol, 2% sodium dodecyl sulfate (SDS), 1% NP-40, 2 mM phenylmethylsulfonyl fluoride, 10 μg/mL leupeptin, 1 mM Na3VO4, 50 mM NaF, and 10 mM EDTA. After cell lysates were centrifuged at 10,000 g for 20 min at 4°C, the protein concentration was determined by using a Bradford protein assay kit (Bio-Rad Laboratories), and 20 μg of protein was subjected to 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to the polyvinylidene difluoride membrane. The membrane was blocked with 5% skim milk in TBST containing 0.1% Tween 20 at room temperature for 1 h and then incubated overnight at 4°C with primary antibodies, PPARγ (Cell Signaling Technology, Danvers, MA, USA; 1/1000), C/EBPα (Cell Signaling Technology; 1/1000), and SREBP1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA; 1/1000), in 5% skim milk in TBST, followed by incubation with secondary antibody conjugated to horseradish peroxidase, antirabbit IgG (Cell Signaling Technology; 1/2000) for 1 h at room temperature. To visualize reaction complexes, the enhanced chemiluminescence system (ECL; Amersham Pharmacia Biotech, Piscataway, NJ, USA) was used and reaction complexes were exposed to X-ray film.
Quantitative real-time reverse transcription polymerase chain reaction
Total RNA was isolated from the 3T3-L1 adipocytes using the Tri reagent (Molecular Research Center, Inc., Cincinnati, OH, USA) according to the manufacturer's instructions. The purity of the RNA was measured at A260 nm/A280 nm using Nanodrop 1000 (Thermo Fisher Scientific, Inc., Franklin, MA, USA) with ratio values of ∼2.0. The cDNAs were synthesized from 2 μg of RNA using high-capacity RNA-to-cDNA reverse transcriptase kit (Bioneer, Inc., Alameda, CA, USA) in a total of 20 μL of reaction volume. Reverse transcription was performed with sample incubation at 42°C for 1 h, followed by 99°C for 5 min. Reverse transcription polymerase chain reaction (RT-PCR) amplifications were performed from 1 μL cDNA diluted at 1:20 using each of the gene-specific primer sets. The oligonucleotide sequence of the forward (sense) and reverse (antisense) primers used for amplification are as in Table 1. Each primer set was used at a concentration of 100 μM in final volume of 14 μL using SYBR® premix Ex Taq (Takara Biotechnology Co. Ltd., Tokyo, Japan). All PCRs were performed using a fluorometric thermal cycler (Thermal Cycler Dice Real Time System; Takara Biotechnology Co. Ltd.). The ΔΔCT method was used to measure relative quantification, where values were normalized to the reference gene (control-insulin+). Individual CT values are means of triplicate measurements, with repeatability measurement of <0.5. Separate control experiments were used to demonstrate that the efficiency of target and reference amplifications were equal.
High-performance liquid chromatography analysis of protocatechualdehyde content in GAM extract
Samples were filtered through a 0.45 μm membrane filter (Millipore, Billerica, MA, USA) and analyzed by means of high-performance liquid chromatography (HPLC). The chromatographic determination was carried out on a Knauer K-501 prominence manufactured by Knauer Company (Berlin, Germany), with a Knauer K-2301 UV/VIS detector. μBondapack C18, 4.6×300 mm, and a 10 μm column was used (Waters, Milford, MA, USA). Gradient elution was performed using water containing 2% acetic acid and acetonitrile at a total flow rate and injection volumes were 1.0 mL/min and 20 μL, respectively. Initially, elution was started with a gradient of 10% acetonitrile changing to 70% in 30 min and finally to 10% in 40 min. The chromatograms at 254 nm were analyzed and compared. The protocatechualdehyde (PCA) was used as commercial products (Sigma-Aldrich, St. Louis, MO, USA).
Statistical analyses
All experiments were performed in triplicate, and all data are expressed as the mean±standard deviation. Statistical analyses were performed using SPSS 20 (SPSS, Inc., Chicago, IL, USA). The level of significance was assessed by variance followed by the Tukey's Honestly Significant Difference (HSD) test. *P<.05, **P<.01, and ***P<.001 were considered statistically significant.
Results
Effect of GAM extract on 3T3-L1 preadipocyte cell toxicity
The cytotoxicity of a GAM extract was examined using 3T3-L1 preadipocyte cells to assess whether it could be useful as an antiobesity agent. Cells were seeded in a 96-well plate and cultured under various concentrations (0, 0.01, 0.1, 0.5, 1, 2, and 3 mg/mL) of GAM extract for 24 h. Cell toxicity was then assessed using the MTT assay. The GAM extract, up to 1 mg/mL, was confirmed to have no significant cytotoxicity against preadipocyte cells, whereas the extract ≥2 mg/mL decreased cell viability (Fig. 1A). Therefore, we concluded that the GAM extract (≤1 mg/mL) did not affect viability of 3T3-L1 preadipocytes.
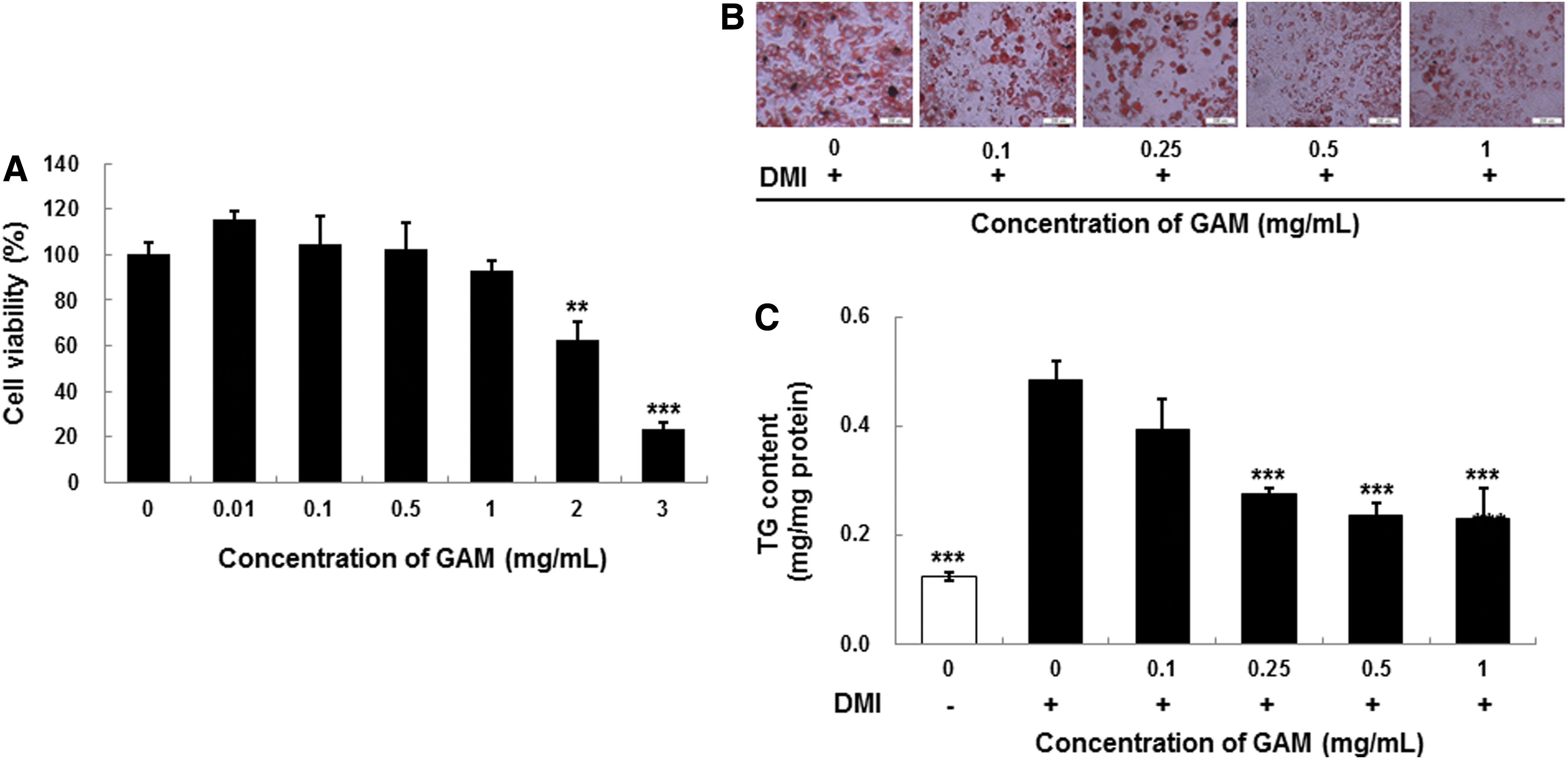
Cell viability
Effect of GAM extract on lipid accumulation assessed by Oil Red O staining
A differentiation medium containing insulin, dexamethasone, and IBMX was used to induce 3T3-L1 preadipocyte differentiation in the presence or absence of GAM extract to examine whether they inhibited adipocyte differentiation. The GAM extract was added to the differentiation medium to observe their effects on 3T3-L1 preadipocyte differentiation. Lipid accumulation and the development of the adipocyte phenotype were assessed by Oil Red O staining in 8 days. As shown in Figure 1B, the GAM extract strongly inhibited preadipocyte differentiation in 3T3-L1 cells during the 8 days. The GAM extract inhibited differentiation of adipocytes in a dose-dependent manner compared with that in control cells. These results show that GAM extract suppressed adipocyte differentiation, indicating a suppression of adipogenesis in 3T3-L1 cells.
Effect of the GAM extract on TG content
As shown in Figure 1C, GAM extract effectively prevented TG accumulation during differentiation of 3T3-L1 preadipocytes in a dose-dependent manner compared with that in control cells. TG content decreased markedly to 69% of that observed in control cells in 3T3-L1 cells incubated with 1 mg/mL GAM extract for 8 days.
Effect of the GAM extract on GPDH activity
GPDH, which is involved in the conversion of glycerol to TG during adipocyte differentiation, was measured after treatment with the GAM extract as an indicator of adipocyte differentiation. The GAM extract decreased GPDH activity in a dose-dependent manner compared with control cells (Fig. 2). GPDH activity markedly decreased to 84% of that observed in control cells in 3T3-L1 cells incubated with 1 mg/mL GAM extract for 8 days.
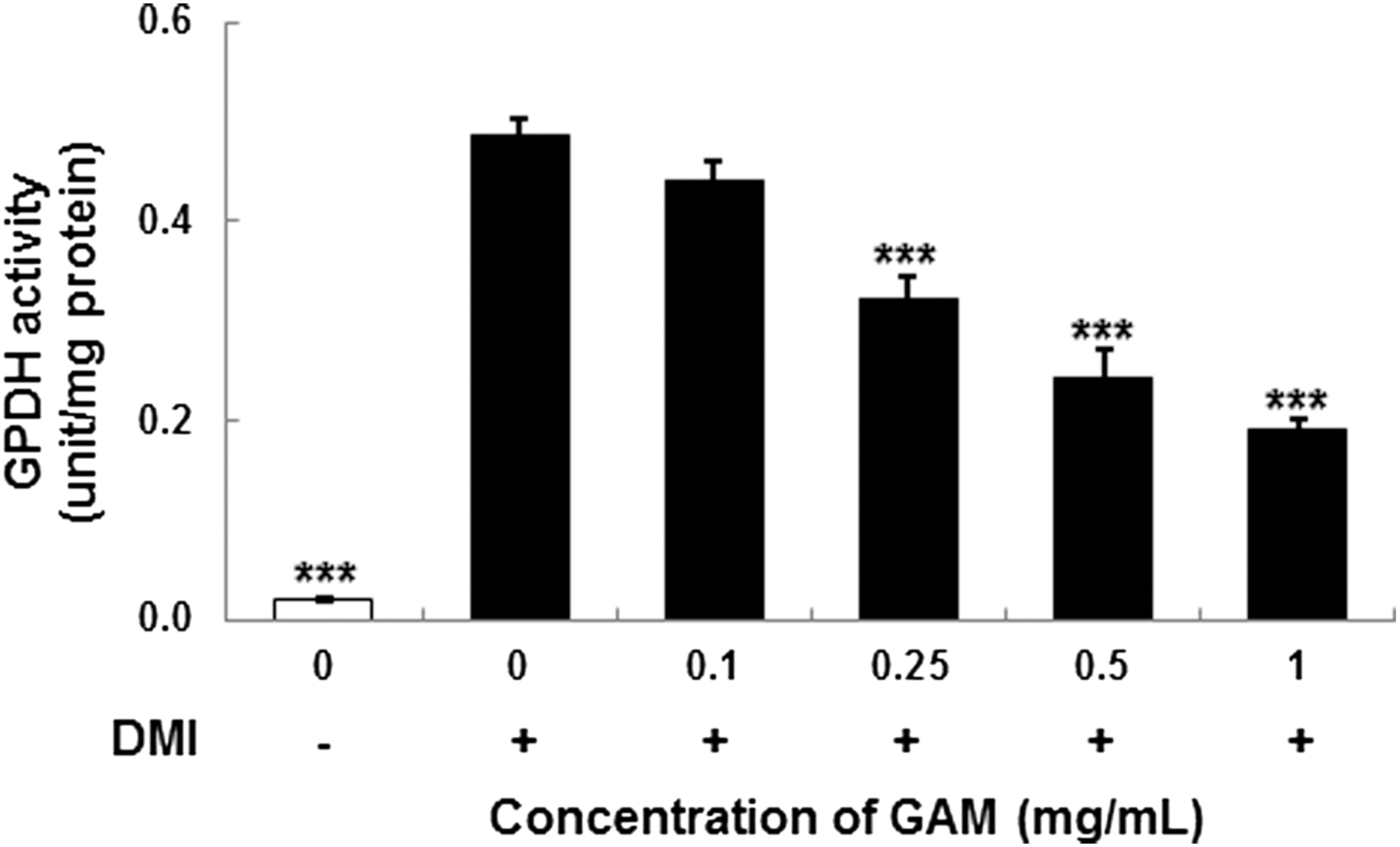
The glycerol-3-phosphate dehydrogenase (GPDH) activity of mature 3T3-L1 adipocytes with treatment of GAM extract at various concentrations. DMI was dexamethasone, IBMX, and insulin. Values are expressed as mean±SD (n=3). ***P<.001 versus control (0+).
Effect of the GAM extract on adipogenic transcription factor protein expression
The effect of GAM extract on the expression of PPARγ, SREBP1, and C/EBPα was evaluated by western blot analysis. As seen in Figure 3, differentiated adipocytes had substantially increased levels of the PPARγ, SREBP1, and C/EBPα compared to preadipocytes. Moreover, treatment of adipocytes with GAM extract (0–1 mg/mL) decreased the expression of PPARγ (Fig. 3A), SREBP1 (Fig. 3B) and C/EBPα (Fig. 3C) in a dose-dependent manner, suggesting that the GAM extract inhibits adipocyte lipogenesis through PPARγ, SREBP1, and C/EBPα-regulated genes.
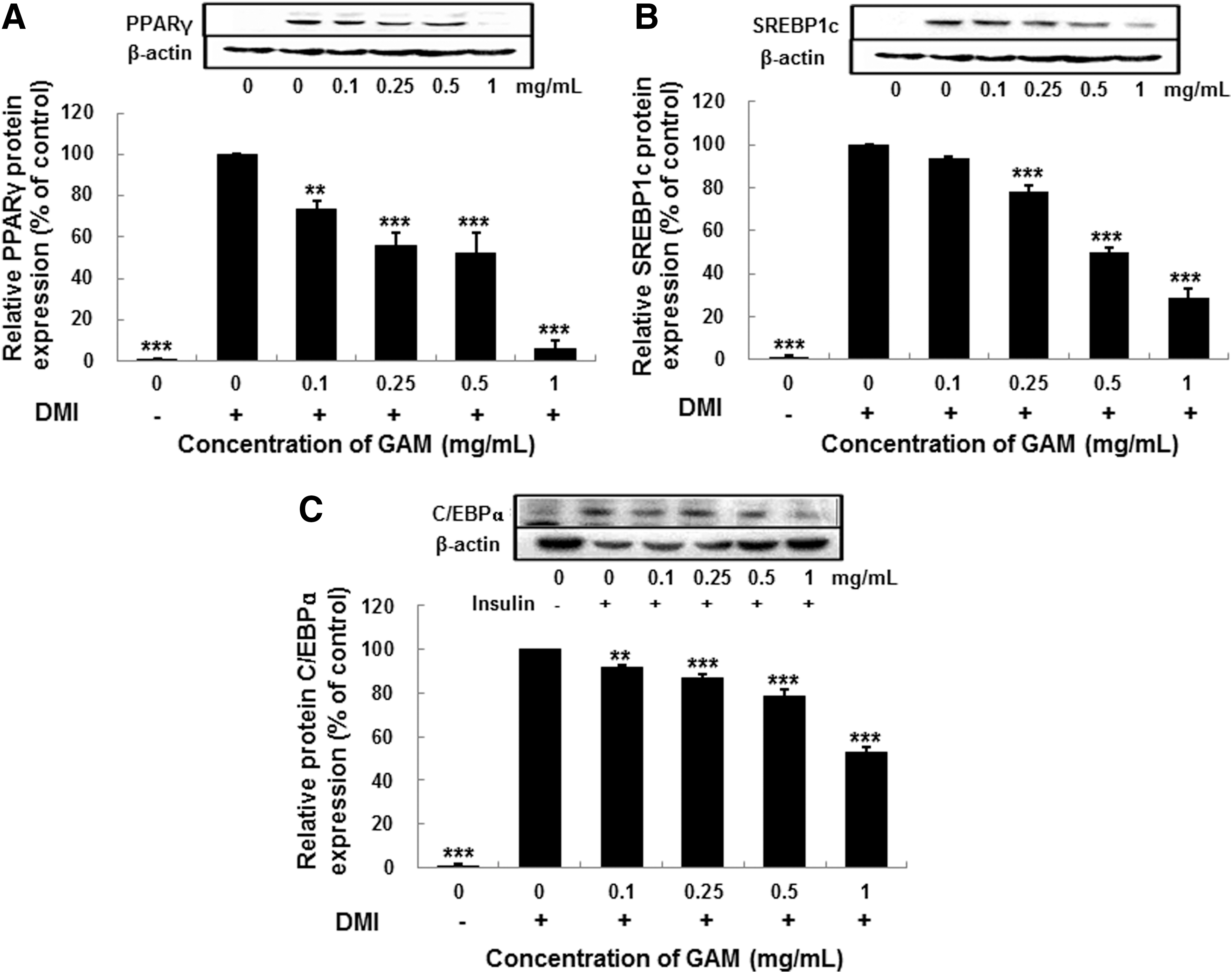
The effect of GAM extract on protein expression of peroxisome proliferator-activated receptor γ (PPARγ)
Effect of the GAM extract on perilipin, FAS, FATP1, and FABP4 mRNA expression
As shown above, GAM extract treatment inhibits the expression of PPARγ, SREBP1, and C/EBPα, which regulate genes responsible for the synthesis and transport of fatty acids. Thus, we determined whether GAM extract also suppresses expression of perilipin, fatty acid synthase (FAS), fatty acid transport protein 1 (FATP1), and fatty acid-binding protein 4 (FABP4), which function as lipid storage, fatty acids synthesis, and transport. 3T3-L1 cells were differentiated and treated with GAM extract (0–1 mg/mL) and expression of perilipin, FAS, FATP1, and FABP4 evaluated by real-time RT-PCR. As seen in Figure 4, GAM extract treatment significantly inhibited the expression of perilipin, FAS, FATP1, and FABP4 in a dose-dependent manner. Therefore, our data suggest that GAM extract suppresses expression of proteins involved in lipid synthesis and lipid transport.

The effect of GAM extract on the levels of adipogenic transcription factors mRNA expression by real-time PCR in 3T3-L1 cells. DMI was dexamethasone, IBMX, and insulin. Values are expressed as mean±SD (n=3). *P<.05, **P<.01, ***P<.001 versus control (0+).
HPLC analysis of the active components in the GAM extract
The HPLC profile of the GAM extract was compared with commercial standard markers. The HPLC chromatogram showed peaks at between 2.4–7.5 min and 10.0–15.0 min with a major peak at 12.6 min. The presence of PCA in the GAM extract at 4.97 min was confirmed by comparing its retention time and UV spectra with that of a PCA reference standard (Fig. 5). It was confirmed that the GAM extract contained 1.41 mg/g of PCA in extract (data not shown).
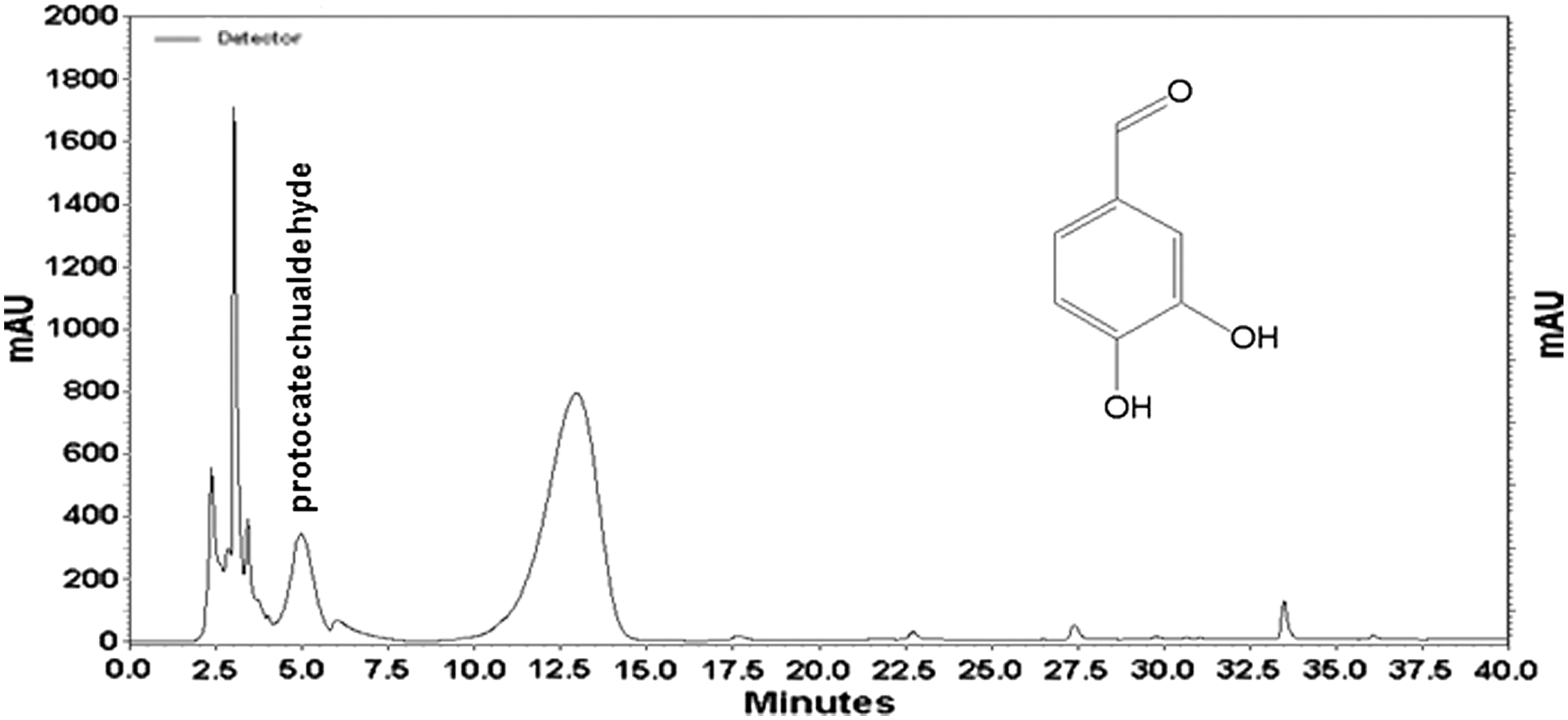
High-performance liquid chromatography chromatogram of GAM extract showing the presence of protocatechualdehyde (PCA). Inset: structure of PCA (3,4-dihydroxybenzaldehyde).
Effect of the PCA on lipid accumulation, TG content, and GPDH activity
To confirm whether PCA inhibits adipocyte differentiation among active components, differentiation medium containing insulin, dexamethasone, and IBMX was used to induce 3T3-L1 preadipocyte differentiation in the presence or absence of 1, 10, and 100 μM PCA. The PCA treatment decreased differentiation of adipocytes in a dose-dependent manner, with the most effective dosage being 100 μM of PCA. Control adipocytes had many lipid droplets (Fig. 6B); however, lipid accumulation was inhibited by the treatments with 10 and 100 μM PCA throughout the differentiation period, and lipid accumulation was progressively decreased by 100 μM PCA. Additionally, TG contents and GPDH activity were decreased during 8 days of differentiation in cells treated with PCA as compared to those in control cells.
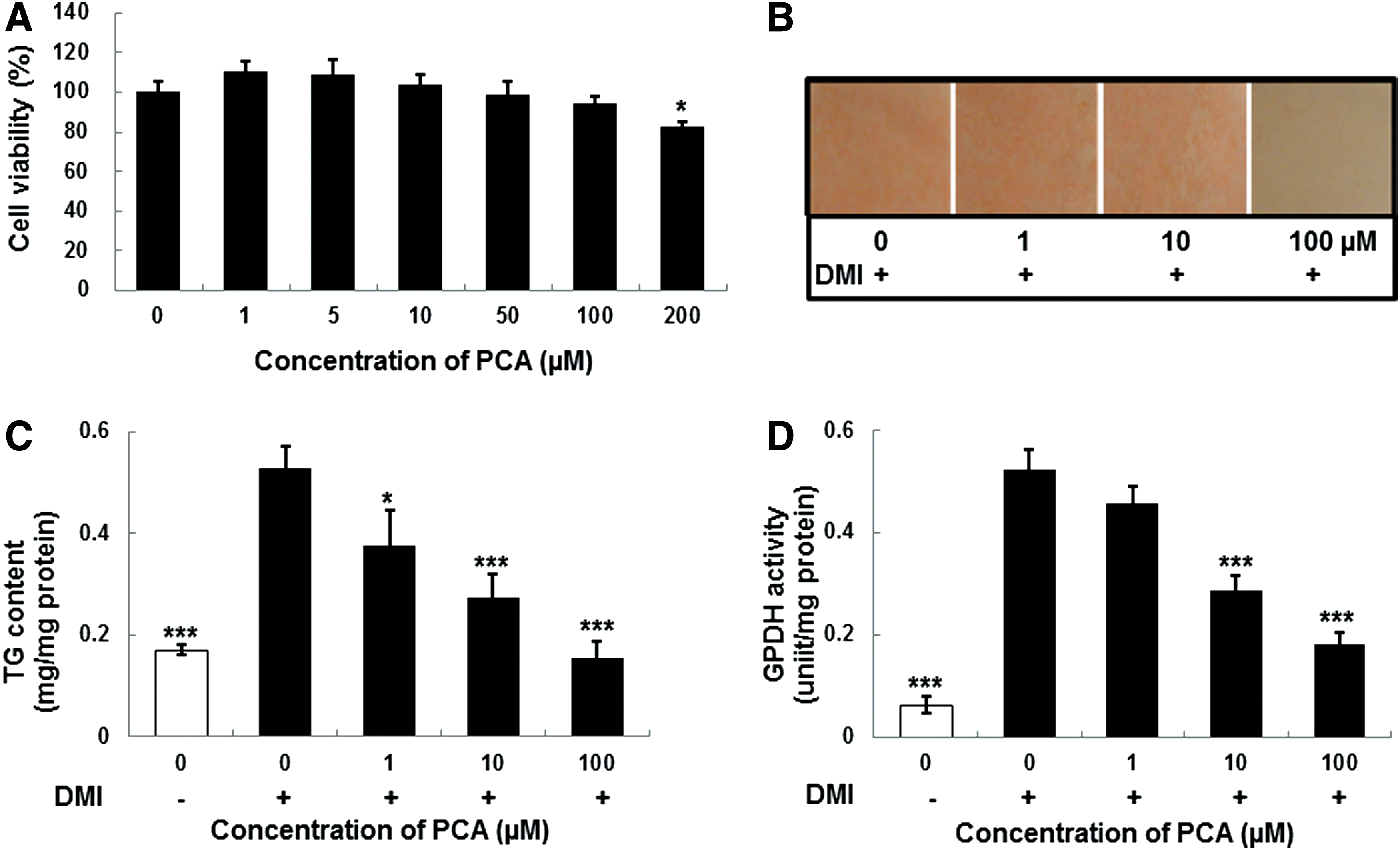
The effect of PCA on cell viability
Discussion
Obesity is the excess accumulation of fat in the fat cells of the body that greatly increases the risk for obesity-associated diseases. 23 Several studies have described that adipocyte differentiation and the amount of fat accumulation in adipocytes are associated with the occurrence and development of obesity. Therefore, inhibiting adipocyte differentiation is one of the strategies for treating obesity. Recent studies have attempted to investigate the beneficial effects of natural products on obesity. In the present study, we demonstrated for the first time that a GAM extract inhibited the differentiation of 3T3-L1 preadipocytes into mature adipocytes by reducing TG accumulation and GPDH activity. Moreover, GAM extracts significantly inhibited the expression of regulatory proteins such as PPARγ, SREBP1, and C/EBPα. Furthermore, it inhibited mRNA expression of modulator proteins such as perilipin, FAS, FATP1, and FABP4 during adipocyte lipid metabolism. Also, PCA, which is a known active component, was confirmed to reduce TG accumulation and GPDH activity during adipocyte differentiation.
Evans et al. 24 suggested the possible mechanisms related to antiobesity actions, including reducing the conversion of glucose and free fatty acids into TG, increasing the oxidation of glucose and/or fatty acids, and increasing lipolysis. We investigated the effects of a GAM extract on lipid accumulation in 3T3-L1 adipocytes by measuring the TG content and used Oil Red O staining as an indicator of lipid accumulation. The reduction in TG content during adipocyte differentiation was more effective when cells were treated with the GAM extract compared to that of the control. Treatment with GAM extract reduced the absorbance values of Oil Red O eluted solution in the cytoplasm of treated cells in a concentration-dependent manner, indicating that the GAM extract inhibited adipogenesis during adipocyte differentiation. 3T3-L1 cells store energy as accumulating TG and dissipate energy as heat, suggesting that the GAM extract enhanced energy expenditure of 3T3-L1 cells.
Cytosolic GPDH appears to play an important role in the conversion process. This enzyme occupies a central position in the TG synthesis pathway where it branches from the glycolytic pathway. 25 GAM extract significantly decreased the GPDH activity in a dose-dependent manner (Fig. 2). Katsuhiko et al. 26 reported that the GPDH activity and TG accumulation decrease when a water soluble extract from the maitake mushroom is added continuously to BAT cells.
Adipocyte differentiation includes a serious of programmed changes in the expression of specific genes. It is well known that adipogenesis can be induced through the action of several enzymes, such as FAS, acetyl-CoA carboxylase (ACC), acyl-CoA synthase (ACS), and glycerol-3-phosphate acyltransferase. Expression of these genes is regulated by transcription factors, such as PPARγ, C/EBPα, and SREBP1, which are critical adipogenesis activators. Expression of these genes occurs early during adipocyte differentiation. 26 –28 PPARγ and C/EBPα, two central transcriptional regulators, are induced before the transcriptional activation of most adipocyte-specific genes. They play central roles in adipocyte differentiation and coordinate the expression of genes involved in creating or maintaining the adipocyte phenotype. 29 Overexpression of these transcription factors can accelerate adipocyte differentiation. SREBP1 critically cross activates the ligand-binding domain of PPARγ 30 and regulates the expression of the enzymes involved in lipogenesis and fatty acid desaturation. 7
The effect of the GAM extract on the expression of these transcriptional factors, at a protein level, was determined. As a result, the GAM extract induced significant downregulation in a dose-dependent manner compared with that of the untreated differentiated adipocytes. The downregulation of SREBP1 and C/EBPα following treatment with the GAM extract may have reduced fatty acid synthesis as well as the synthesis and activity of PPARγ, which may have resulted in the inhibition of lipid accumulation by blocking adipogenesis. The PPARγ and C/EBPα synergistically activate the downstream adipocyte-specific gene promoters, such as FAS, ACS1, FABP4, FATP1, and perilipin at the late stage, leading to fat droplet formation. 31 Through differentiation induction, the mRNA expressions of FAS, ACS1, FABP4, FATP1, and perilipin, which are subfactors of transcription factors such as PPARγ, SREBP1, and C/EBPα, were increased; however, the expressions were decreased due to the treatment of GAM extract.
FAS is expressed early in the differentiation of adipocytes and it is an enzyme that produces palmitate from acetyl-CoA and malonyl-CoA. While FAS causes obesity by increasing the storage of excess energy in the form of TGs, it is reported that the TG palmitate produced by FAS causes insulin resistance through the destruction of pancreatic β cells. 32 FABP4 is expressed in adipocytes and it is known to regulate intracellular fatty acid and lipid metabolisms. 33
Ganoderma spp. is a well-known Chinese traditional medicine that has long been used clinically in China, Japan, and Korea. More than 130 highly oxygenated lanostane-type triterpenoids have been isolated from the fruiting bodies, mycelia, and spores of Ganoderma lucidum, including common fungal steroids derived from ergosterol, some of which exhibit a bitter taste or have useful biological activities. 31,34
Several lanostane triterpenes isolated from G. lucidum have an in vitro inhibitory effect on adipocyte differentiation in 3T3-L1 cells. 35 PCA is a naturally occurring polyphenol (3,4-dihydroxybenzaldehyde) found in barley, green Cavendish bananas and grapevine leaves. Lee et al. 36 reported that PCA isolated from GA fruiting bodies acts as an aldose reductase inhibitor and may be useful for preventing and/or treating diabetic complications. Therefore, the GAM extract was analyzed using HPLC to identify the active components and the PCA content was quantified. Lipid accumulation by Oil Red O staining, TG content, and GPDH activity was determined to confirm the inhibitory effect on adipocyte differentiation in 3T3-L1 cells in the presence of PCA. Lipid droplet accumulation dose dependently decreased in the Oil Red O stained cells (Fig. 6B). Additionally, the TG content and GPDH activity decreased in the differentiation process of cells treated with PCA during 8 days. This result could be related to the observation that fatty acid synthesis from glucose was suppressed when PCA was added to the 3T3-L1 cells.
In conclusion, GAM extract inhibited adipocyte differentiation, as indicated by decreased TG content and GPDH activity, and appeared to be mediated through downregulated expression of the adipogenic transcription factors PPARγ, C/EBPα, and SREBP1 as well as the adipogenic-specific genes perilipin, FAS, FATP1, and FABP4. In addition, we confirmed that the GAM EtOH extract contained 1.44 mg/g PCA as an active component. These findings suggest that GAM extracts containing PCA might have a beneficial effect by preventing adipogenesis-related obesity and may be a potential natural drug for the treatment of obesity.
Footnotes
Acknowledgments
This research was supported by the High Value-added Food Technology Development Program, Ministry for Food, Agriculture, Forestry and Fisheries, Republic of Korea, supported (in part) by the Ministry of Education, Science Technology (MEST), and the National Research Foundation of Korea (NRF) through the Human Resource Training Project for Regional Innovation.
Author Disclosure Statement
No competing financial interests exist.