
Abstract
Fresh ginseng was fermented with Ganoderma lucidum mycelium (GL) in solid-state culture to enhance its immunomodulatory activity. When crude polysaccharide (FG-GL-CP) was fractionated from lyophilized fermented ginseng (FG-GL), FG-GL-CP showed significantly higher mitogenic, macrophage stimulating, and intestinal immune system modulating activity (1.52-, 1.52- and 1.46-fold, respectively) than those of crude polysaccharide from nonfermented ginseng (NG-CP, 1.36-, 1.37- and 1.32-fold) and G. lucidum mycelium alone (GL-CP, 1.48-, 1.40- and 1.31-fold). After FG-GL-CP was further fractionated into two active carbohydrate-rich fractions (FG-GL-CP-II and III) on DEAE-Sepharose CL-6B (Cl− form), the most active polysaccharide (FG-GL-CP-III-1) was purified as a single peak with a molecular weight of 71 kDa. FG-GL-CP-III-1 mainly consists of Ara, Man, Gal, and Glc (molar ratio: 0.85:1.00:0.81:0.76) in addition to a small amount of uronic acid. Methylation analysis indicates that FG-GL-CP-III-1 is composed mainly of terminal/4- or 5-linked Ara f , terminal/4-linked Man, terminal/3,6-branched Gal, and terminal/4-linked Glc. NaIO4 oxidation of FG-GL-CP-III significantly decreased its activity (64.0–79.2%). These results suggest that solid-state culture of ginseng with G. lucidum may enhance the immunomodulatory activity and that neutral polysaccharides containing Ara, Man, Gal, and Glc may be important for the activity of fermented ginseng.
Introduction
G
Solid-state culture, a process where an insoluble substrate is fermented with sufficient moisture but without free water, is used for potentiating the physiological values of waste materials, enzyme production, and the value of existing foods. 6 Solid-state culture requires no complex fermentation controls and has many advantages such as less energy input over submerged liquid fermentation. Higher fungi, especially mushrooms, produce numerous unique bioactive compounds, and their fermentation studies have received much interest in recent decades. 7,8 Ganoderma lucidum, a traditional Korean medicinal mushroom, has been used for the prevention and treatment of various human diseases for several thousand years in Asia. 9 Han et al. 10 reported that the nutritional value of cornmeal was increased by solid-state culture with G. lucidum. Thus, we hypothesized that G. lucidum mycelium culture on ginseng can increase the amounts and/or potencies of pharmacological ingredients by the biotransformation process of solid-state culture than ginseng or mycelium alone.
The aim of this research was to enhance the immunomodulatory activity of ginseng by solid-state culture with G. lucidum mycelium. For this, we fermented fresh ginseng with G. lucidum, and its immunomodulating activity was compared with those of nonfermented ginseng and G. lucidum mycelium alone. Furthermore, the active polysaccharide responsible for the enhanced activity from fermented ginseng was purified and characterized.
Materials and Methods
Mushroom mycelium and fresh ginseng
The mushroom mycelium of G. lucidum was obtained from the Agricultural Research and Extension Services (Chungbuk, Korea). Fresh ginseng (P. ginseng, 5-year cultivation) for solid-state culture was purchased from the Farming Association of Jeungpyeong-gun (Chungbuk, Korea) and had been kept in a freezer (−20°C) before being used for fermentation. After ginseng extract for mycelium seed culture was prepared from fresh ginseng by decoction (95°C) with distilled water (20 volumes), the supernatant was concentrated to extracts of 65°Bx.
Preparation of fermented ginseng with G. lucidum by solid-state culture
The optimal mushroom complete medium (MCM) for G. lucidum mycelium seed contained glucose 20.0 g, peptone 2.0 g, yeast extract 2.0 g, and 1.0, 0.4, and 0.5 g of K2HPO4, KH2PO4, and MgSO4·7H2O, respectively (per liter). The seed was cultured in a flask containing MCM supplemented with ginseng extract (65°Bx, 5% v/v) at 30°C, pH 5.5, for 6 days in a shaking incubator (Jeio Tech., Daejeon, Korea). Meanwhile, the washed fresh ginseng was dried to control the moisture content and soften the tissue and then 10% (w/w, a ratio of the dried fresh ginseng) of brown rice powder was added as a nutrient source for the G. lucidum. After autoclaving, 10% (w/w) of G. lucidum was seeded and cultivated at 30°C in a multiroom incubator (Vision Scientific, Gyeonggi-do, Korea) for 30 days followed by lyophilization into fermented ginseng (FG-GL, Fig. 1). In addition, G. lucidum mycelium was cultured in MCM without ginseng extract using the same culture conditions, and the recovered mycelium was lyophilized for the preparation of G. lucidum mycelium alone (GL) as a control.
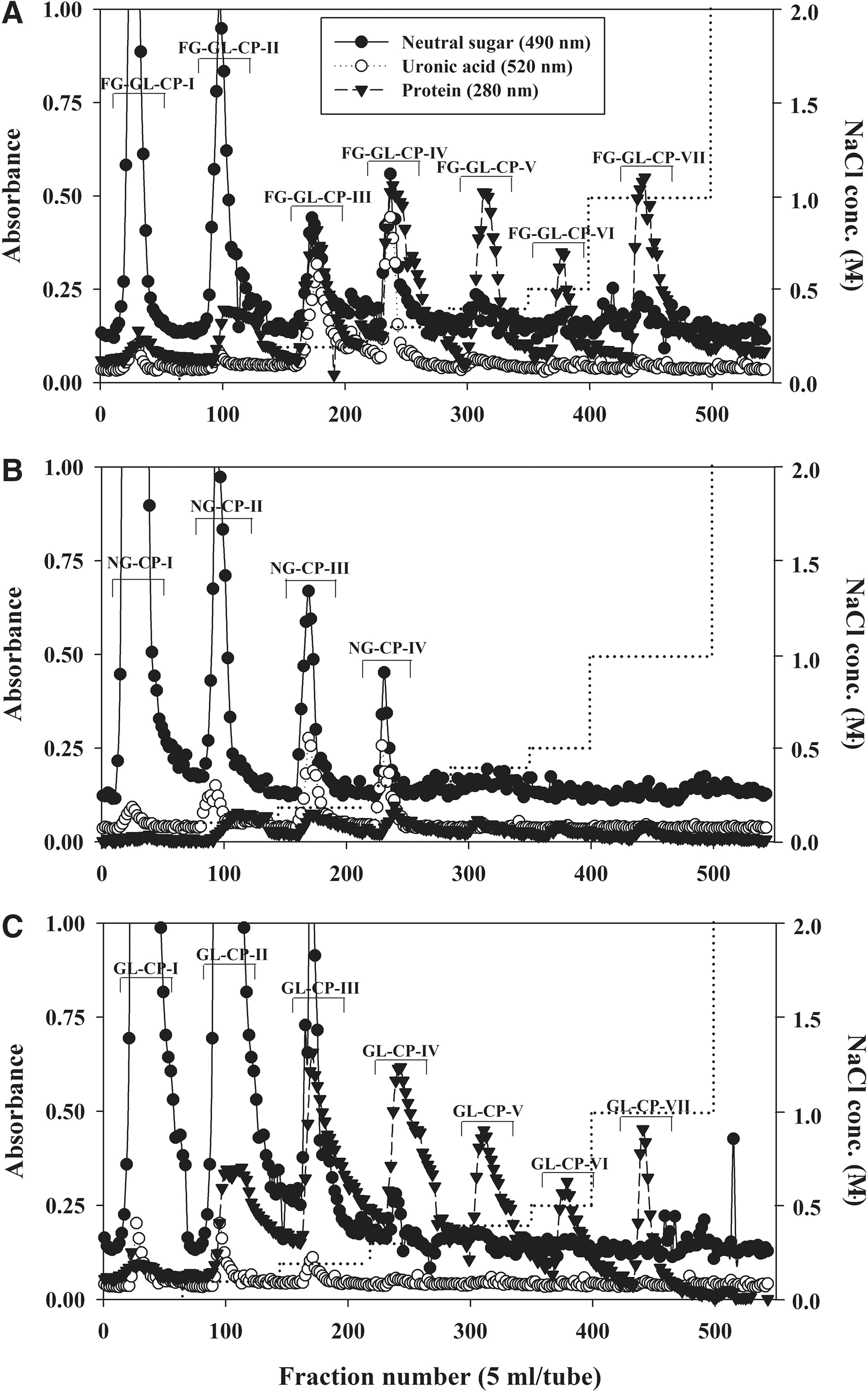
Elution profiles of crude polysaccharide fractions from
General methods
Total carbohydrate, uronic acid, and protein contents were determined by Gal, 11 GalA, 12 and BSA 13 as the respective standards (Sigma-Aldrich Co., St. Louis, MO, USA). The aldose component of the polysaccharides was analyzed as alditol acetate by NaBH4 reduction and acetic anhydride acetylation after hydrolysis with 2 M TFA for 1.5 h at 121°C. 14 For uronic acid determination, the reduced hydrolysates were loaded onto Dowex-1 resin and the bounded aldonic acids were eluted by 1.0 N HCl. The eluted fraction was evaporated to dryness, and the resulting lactones were reduced under neutral conditions. 14 After acetylation, the resulting carboxyl-reduced alditol acetates were analyzed by GC according to the procedure of Zhao et al. 15 The GC (HP 5890 A; Hewlett-Packard, Palo Alto, CA, USA) was equipped with FID using a Supelco SP-2380 capillary column (30 m×0.25 mm inner diameter, 0.2 μm film; Bellefonte, PA, USA). Molecular weight of the purified polysaccharide fraction was determined using a Waters 2690 system (Waters Co., Milford, MA, USA) equipped with column (8 mm×300 mm, Shodex® OHpak KB-805; Showa Denko, Tokyo, Japan) and RI detector. The molecular weight of the polysaccharide was estimated from a calibration curve constructed for standard dextrans (T-2000, T-500, T-70, and T-20; Showa Denko) and glucose. 0.2 M NaCl was used as the mobile phase at the flow rate of 0.5 mL/min.
Fractionation of crude polysaccharide from fermented ginseng
Fermented ginseng (FG-GL, 100 g) was decocted with distilled water (2 L) to half volume (three times). All extracts were centrifuged (7600 g, 4°C, 20 min; Herolab GmbH Laborgerate, Wiesloch, Germany), and the supernatant was lyophilized to give hot water extract (FG-GL-HW). After five volumes ethanol were added to the dissolved FG-GL-HW and stirred, the resulting precipitate was dialyzed (Spectra/Por 4, MWCO 12–14,000; Spectrum Laboratories, Inc., Rancho Dominguez, CA, USA) against distilled water. Finally, the insoluble materials were removed and the nondialyzable portion was lyophilized to obtain crude polysaccharide fraction (FG-GL-CP).
Purification of immunomodulating polysaccharide from FG-GL-CP
FG-GL-CP (1 g) was applied to a column (4.0 cm×40 cm) of DEAE-Sepharose CL-6B (Cl− form; Amersham Biosciences, Uppsala, Sweden) equilibrated with distilled water. The column was eluted with distilled water to obtain a nonadsorbed fraction (FG-GL-CP-I; yield 5.4% of FG-GL-CP). The absorbed fractions were eluted by stepwise gradient elution with 0.1, 0.2, 0.3, 0.4, 0.5, and 1.0 M NaCl. Six absorbed fractions (FG-GL-CP-II, III, IV, V, VI and VII) were obtained as lyophilizates after dialysis (0.1 M NaCl; FG-GL-CP-II, yield 13.0%: 0.2 M; FG-GL-CP-III, 23.6%: 0.3 M; FG-GL-CP-IV, 8.3%: 0.4 M; FG-GL-CP-V, 6.2%: 0.5 M; FG-GL-CP-VI, 3.9%: 1.0 M; FG-GL-CP-VII, 2.4%). After the immunomodulating polysaccharide fraction (FG-GL-CP-III) was further fractionated by Sepharose CL-6B (Amersham Biosciences; 2.6 cm×90 cm, eluant of 0.2 M NaCl), the most active polysaccharide (FG-GL-CP-III-1; yield 10.3% of FG-GL-CP-III) was purified in the void volume.
Chemical and enzymatic treatments of the active polysaccharide fraction
Periodate (NaIO4) oxidation and pronase digestion of the active polysaccharide fraction (FG-GL-CP-III) were performed as described by Yamada et al. 16 Briefly, FG-GL-CP-III (10 mg) was oxidized in 50 mM acetate buffer (pH 4.5) containing 25 mM NaIO4 (10 mL) at 4°C for 96 h in the dark. The NaIO4-treated products were reduced with NaBH4 and then dialyzed to obtain the oxidized-FG-GL-CP-III (yield; 64.8%). In addition, FG-GL-CP-III (10 mg) was digested with Actinase E (10 mg; Kaken Pharmaceutical Co. Ltd., Tokyo, Japan) in 50 mM Tris-HCl buffer (pH 7.9, 10 mL) containing 10 mM CaCl2 at 37°C for 48 h. After the reaction was terminated by neutralization with 0.1 N HCl, pronase-treated products were dialyzed and lyophilized to obtain the digested-FG-GL-CP-III (yield; 78.7%).
Methylation analysis
Methylation analysis was performed according to the Hakomori method, 17 and the methylated products were recovered by the modified procedure of Waeghe et al. 18 After samples (500 μg) were completely solubilized in dry DMSO and methylated with methylsulfinyl carbanion and methyl iodide, the methylated samples were recovered by ethanol elution using a Sep-pak C18 cartridge (Waters Co.). The products were hydrolyzed with 1 M TFA for 2 h at 121°C and reduced with NaBH4 followed by acetylation with (CH3CO)2O. The resulting partially methylated alditol acetates were analyzed by GC and GC-EIMS using a SP-2380 capillary column. 15,19 GC-EIMS was performed with an HP 5890 A GC equipped with a 5970B mass-selective detector (Hewlett-Packard), and He was the carrier gas (0.5 mL/min). The compounds were identified by their fragment ions and relative retention time, and their molar ratios were estimated from the peak areas and response factors. 20
Mice and cell culture
The experimental protocol was reviewed and approved by the Animal Research Committee of Korea University (Seoul, Korea). Specific pathogen-free C3H/He mice (female, 6 weeks old) were purchased from Orient Bio (Gyeonggi-do, Korea), and BALB/c and ICR mice (female, 6 weeks old) from Nara Biotech Co., Ltd. (Gyeonggi-do, Korea). Mice were housed and maintained at a constant temperature, 24°C±1°C and humidity (55%) with 12 h cycle of light and dark. Mice were fed with a commercial chow pellet diet (Samyang Co. Ltd., Incheon, Korea), and water was freely available. Primary cells, such as peritoneal macrophage, splenocyte, and Peyer's patch cells, from each mouse were cultured in RPMI-1640 (Gibco-BRL Co., Grand Island, NY, USA) supplemented with fetal bovine serum (FBS; Cell Culture Laboratories, Cleveland, OH, USA) and penicillin (100 U/mL), streptomycin (100 μg/mL), and amphotericin B (Flow Laboratories, Irvine, Scotland) at 37°C in a 5% CO2 incubator.
Mitogenic activity of splenocytes
Spleen lymphocytes (5×106 cells/well) from BALB/c mice were co-cultured with samples in 96-well culture plates (MaxiSorp™ F96; Nunc™, Roskide, Denmark) for 48 h. Lymphocyte proliferation was assayed by cell counting kit (CCK-8; Dojindo Laboratories, Kumamoto, Japan). 21 Four hours before culture termination, 20 μL of CCK-8 solution was added to each well and the cells were continuously incubated (4–24 h). The culture was measured at 450 nm by a microtiter plate reader (Sunrise; Tecan, Grödinger, Austria). The mitogenic activity was expressed as relative percentage of splenocyte growth compared to the saline control incubated without the test sample.
Macrophage stimulating assay
ICR mice were injected interperitoneally with 1 mL of 3% thioglycollate medium. After 3 days, macrophage cells were prepared from the peritoneal cavity of mice by washing with the cold RPMI 1640 medium containing 5 mM HEPES, penicillin, and streptomycin. An aliquot (200 μL) of the cell suspension (1×106 cells/mL) was seeded and cultured in 96-well culture plates under a humidified atmosphere of 5% CO2 and 95% air (2 h, 37°C). Nonadherent cells were removed by washing with RPMI-1640 medium, and then, the adherent cells were prepared into a macrophage monolayer. 22 Macrophage stimulating activity was assayed by the procedures of Suzuki et al. 23 After macrophage monolayer cells (1×105 cells/mL) were cultured with samples for 24 h, cells were solubilized by the addition of 25 μL of 0.1% Triton X-100. One hundred fifty microliters of 10 mM p-nitrophenyl phosphate was added to the reaction mixture, and the absorbance at 405 nm was measured using a microplate reader as described previously.
Intestinal immune system modulating activity using Peyer's patch cells
Suspensions of Peyer's patch cells in RPMI 1640 medium supplemented with 5% FBS (RPMI 1640-FBS) were prepared from small intestines of C3H/He mice. 24 One hundred eighty microliters of aliquots of the cell suspension (2×106 cells/mL in RPMI 1640) was cultured with the samples (20 μL) in a 96-well flat bottom microtiter plate for 5 days at 37°C in a humidified atmosphere of 5% CO2 and 95% air. Then, the resulting culture supernatants (50 μL) were then incubated with bone marrow cells suspension (2.5×105 cells/mL in RPMI 1640-FBS) from C3H/He mice for 6 days in the same incubator. After 20 μL of CCK-8 solution was added to each well, the cells were continuously cultured for 4–6 h and the intensity was measured by microplate reader at 450 nm. The intestinal immune system modulating activity was expressed as relative percentage of the bone marrow cell growth compared to the saline control incubated without the test sample.
Statistical analysis
All measurements were carried out in quadruplicate. Data were analyzed using the Statistical Package for Social Sciences (SPSS, Inc., Chicago, IL, USA). A one-way analysis of variance was performed for each response variable. The results were presented as the mean±standard deviation. The statistical analysis was assessed by Tukey's multiple range tests, and the significance was determined at P<.05.
Results
Immunomodulating enhancement of crude polysaccharide from fermented ginseng
To enhance the immunomodulatory activity of ginseng, fresh ginseng was fermented with G. lucidum mycelia in solid-state culture and lyophilized (FG-GL, Fig. 2). After hot water extraction (FG-GL-HW, yield from FG-GL; 31.3%, w/w) was used to prepare the FG-GL decoction, FG-GL-HW was fractionated by ethanol precipitation to obtain a crude polysaccharide fraction (FG-GL-CP, yield from FG-GL; 3.6%). Hot water extracts and crude polysaccharides were also fractionated from nonfermented ginseng (NG-HW, NG-CP) and G. lucidum mycelium alone (GL-HW, GL-CP) as controls, respectively. As shown in Table 1, FG-GL-HW (1.39-fold of saline control at 100 μg/mL) was higher than NG-HW (1.24-fold; P<.05) and GL-HW (1.32-fold) in mitogenic activity in splenocytes. Crude polysaccharide (FG-GL-CP, 1.52-fold) fractionated from FG-GL-HW also showed the potent mitogenic activity compared to NG-CP (1.36-fold) and GL-CP (1.48-fold; Table 1). Hot water extractions of mycelium, nonfermented ginseng, and fermented ginseng (GL-HW, NG-HW, and FG-GL-HW, respectively) resulted in 1.22-, 1.26-, and 1.30-fold greater stimulation of macrophages compared to saline control but were not statistically significant (Table 1). However, FG-GL-CP enhanced the activity by 1.52-fold compared to NG-CP (1.37-fold; P<.05) and GL-CP (1.40-fold). In addition, although FG-GL-HW did not exert potent intestinal immune system modulating activity through Peyer's patch in hot water extract, FG-GL-CP (1.46-fold) showed significantly higher activity than controls (NG-CP, 1.32-fold; GL-CP, 1.31-fold; Table 1). These results suggest that crude polysaccharide fractions from fermented ginseng presented more potent immunomodulating activities than those of nonfermented ginseng and G. lucidum mycelium alone.
Fermented ginseng with G. lucidum in solid-state culture.
Results are expressed as mean±SD (n=4)
Results in the same column with different superscript letters are significantly different at P<.05.
NG, nonfermented ginseng; GL, G. lucidum mycelium alone; FG-GL, fermented ginseng with G. lucidum mycelium; NG-, GL-, and FG-GL-HW, hot water extracts from NG, GL, and FG-GL, respectively; NG-, GL-, and FG-GL-CP, crude polysaccharide fractions from NG, GL, and FG-GL, respectively; SD, standard deviation.
To further elucidate the difference between the more active crude polysaccharide from fermented ginseng and those from nonfermented ginseng and G. lucidum mycelium alone, neutral sugar, uronic acid, and protein contents of crude polysaccharide 25 were analyzed (Table 2). Neutral sugar content in FG-GL-CP from fermented ginseng was lower compared with NG-CP and GL-CP (63.1% vs. 85.1% and 87.6%), and FG-GL-CP contained more uronic acid and higher protein content (24.5% vs. 12.4% and 7.0%, and 12.4% vs. 2.4% and 5.3%, respectively). These changes in the main component contents (neutral sugar and uronic acid) of polysaccharide suggested that fresh ginseng has been changed in its chemical composition by G. lucidum mycelium in solid-state culture.
Neutral sugar, uronic acid, and protein were determined by phenol-sulfuric acid (490 nm), the m-hydroxybiphenyl method (520 nm), and the Bradford method (595 nm), respectively. Results are expressed as mean±SD (n=4).
Results in the same column with different superscript letters are significantly different at P<.05.
NG-, GL-, and FG-GL-CP-II and -III, crude polysaccharide subfractions from NG, FG, and FG-GL using a DEAE-Sepharose CL-6B; FG-GL-CP-III-1 and FG-GL-CP-III-2, fractions from FG-GL-CP-III using a Sepharose CL-6B.
Immunomodulating polysaccharide purification from fermented ginseng
FG-GL-CP, which had the enhanced immunomodulating activity compared to nonfermented ginseng, was further fractionated by anion-exchange chromatography on DEAE-Sepharose CL-6B (Cl− form), and a nonadsorbed (FG-GL-CP-I) eluted with distilled water and 6 adsorbed fractions (FG-GL-CP-II to VII) by 0.1–1.0 M NaCl were obtained (Fig. 1A). NG-CP and GL-CP were also fractionated into a nonadsorbed fraction (NG-CP-I and GL-CP-I) and three (NG-CP-II–IV) or six adsorbed fractions (GL-CP-II to VII) on the same column (Fig. 1B, C). When the elution profiles of crude polysaccharides were compared (Fig. 1), fractions from FG-GL-CP showed more uronic acid and less neutral sugar contents than those from NG-CP and GL-CP. In addition, fractions (FG-GL-CP-V–VII and GL-CP-V–VII) eluted by NaCl concentrations over 0.4 M were detected in fractionation of FG-GL-CP and GL-CP (Fig. 1). Furthermore, FG-GL-CP-III showed the most potent mitogenic activity (1.76-fold of the saline control at 100 μg/mL) and significantly higher mitogenic effect than fractions from nonfermented ginseng (NG-CP-III, 1.44-fold; NG-CP-IV, 1.54-fold) and G. lucidum mycelium (GL-CP-II, 1.40-fold; GL-CP-III, 1.53-fold), respectively (Fig. 3A). When fractions from crude polysaccharide on DEAE-Sepharose CL-6B were tested on the macrophage stimulating activity in vitro, FG-GL-CP-III also showed the higher activity (1.82-fold; P<.05) than any other fractions, including the same NaCl elution fraction from nonfermented ginseng (NG-CP-III, 1.59-fold) and mycelium (GL-CP-III, 1.64-fold; Fig. 3B). Fractions from crude polysaccharide of fermented ginseng were also examined to determine if they can modulate the activity of intestinal immune system modulating activity through Peyer's patches. FG-GL-CP-II and III had the more potent activity (1.90 and 1.81-fold, respectively) than those of NG-CP (NG-CP-II, 1.51-fold; NG-CP-III, 1.65-fold; P<.05) and GL-CP (GL-CP-II, 1.36-fold; GL-CP-III, 1.42-fold; P<.05), respectively (Fig. 3C). Taken together, these results suggest that FG-GL-CP-III has the potent activity of all immunomodulating assays (Fig. 3) and the highest yield (23.6% of FG-GL-CP) among carbohydrate-rich fractions. Thus, we purified the active polysaccharides from FG-GL-CP-III of fermented ginseng and identified its structural properties. When FG-GL-CP-III was fractionated by gel filtration on Sepharose CL-6B, two fractions (FG-GL-CP-III-1 and 2) were obtained (Fig. 4A). Of these fractions, FG-GL-CP-III-1 showed the greatest immunomodulatory activity (mitogenic activity, 1.88-fold of saline control at 100 μg/mL; macrophage stimulation, 2.13-fold; intestinal immune system modulation, 1.76-fold), whereas FG-GL-CP-III-2 had only weak activity (1.36, 1.62, and 1.28-fold, respectively; Table 3).
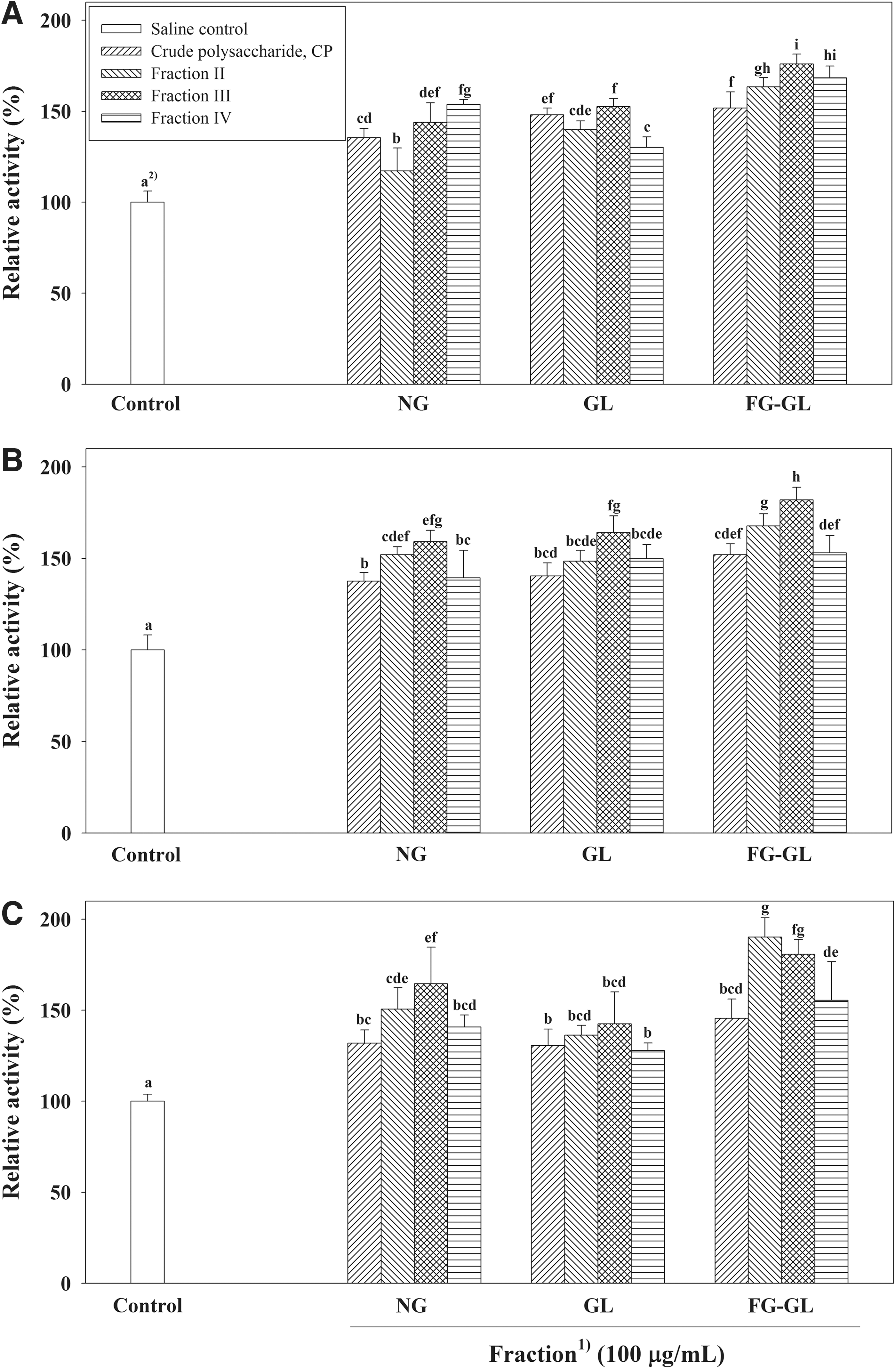
Immunomodulating activities of the active fractions from crude polysaccharide of fermented ginseng on DEAE-Sepharose CL-6B column chromatography.

Results are expressed as mean±SD (n=4).
Results in the same column with different superscript letters are significantly different at P<.05.
The immunomodulating polysaccharide fraction (FG-GL-CP-III-1) was further analyzed by HPLC on Asahi-pak GS-510+320+220 using an isocratic mobile phase (0.2 M NaCl) at 0.5 mL/min of flow rate. The active fraction was eluted as a single and symmetrical peak, and its molecular weight was estimated to be 71 kDa (Fig. 4B).
Identification of the purified immunomodulating polysaccharide
As shown in Table 2, the active carbohydrate-rich fractions (FG-GL-CP-II and III) were composed of neutral sugar (88.8% and 69.2%, respectively) and uronic acid (7.5% and 27.0%). The less active polysaccharides from nonfermented ginseng (NG-CP-II/III) and G. lucidum mycelium (GL-CP-II/III) also mainly contained 82.4/65.5% and 90.5/75.1% of neutral sugar with 9.6/26.0% and 6.8/19.3% of uronic acid, respectively (Table 2). However, the purified active fraction (FG-GL-CP-III-1) mainly composed of neutral sugar (91.1%) with small amounts of uronic acid and protein (3.6% and 1.1%), whereas the other fraction (FG-GL-CP-III-2) having a weak activity consisted of neutral sugar and uronic acid (62.1% and 33.7%, respectively; Table 2). Therefore, component sugars of these main fractions were analyzed to identify the compositional difference of polysaccharides from fermented and nonfermented ginseng or G. lucidum. The monosaccharide compositions of the collected fractions separated from crude polysaccharide of fermented ginseng by DEAE-Sepharose CL-6B chromatography are listed in Table 4. As shown, NG-CP-III and FG-GL-CP-III contained considerable amounts of acidic sugars (29.31 and 32.60 mol%), such as GalA and GlcA. NG-CP-III also contained the main neutral sugars, such as Ara, Gal, Rha, and Glc (27.79%, 23.26%, 7.60%, and 5.28%). Although FG-GL-CP-III also contained Ara (16.31 mol%) and uronic acid (32.60%), Gal was detected in less (9.52%) than half of NG-CP-III and Man was significantly increased (12.19%) in FG-GL-CP-III (Table 4), suggesting that the active polysaccharide in fermented ginseng resulted from G. lucidum mycelium growth or enzymes released from mycelium. The polysaccharide fraction (GL-CP-III) from mycelium contained less Ara and uronic acid (1.66 and 17.90 mol%) and much more Glc (42.80%) compared to FG-GL-CP-III. In addition, less active polysaccharide fractions, NG-CP-II, GL-CP-II, and FG-GL-CP-II, contained Gal (40.78, 13.08, and 32.31 mol%), Glc (14.27%, 44.63%, and 17.31%), Ara (22.62%, 1.08%, and 2.59%), and Man (6.44%, 21.24%, and 19.34%, respectively) in addition to a small amount of uronic acid (Table 4). Component sugar analysis showed that the purified active FG-GL-CP-III-1 consisted mainly of Man, Ara, Gal, and Glc (molar ratio of 1.00:0.85: 0.81:0.76) similar to FG-GL-CP-III. However, the level of uronic acid in FG-GL-CP-III-1 was significantly lower (5.23%) than FG-GL-CP-III (Table 4). In contrast, the weak active fraction (FG-GL-CP-III-2) contained the main neutral sugars (58.4%) as well as large amounts of GalA and GlcA (35.2%; data not shown). Methylation analysis also indicated that FG-GL-CP-III-1 was composed mainly of 4-linked Glc or Man, terminal Man, Gal, Glc, or Ara, and 4- or 5-linked Ara, 2,4-branched Rha or 3,6-branched Gal for neutral sugar (Table 5). Glycosidic linkages of uronic acid were not analyzed because of low concentration.
The active fraction (FG-GL-CP-III) was treated with NaIO4 and pronase to elucidate which moiety contributes to the activity. NaIO4 oxidation significantly decreased the activity of FG-GL-CP-III (64.0–79.2%), whereas pronase digestion did not affect the activity (Table 6). These results suggest that polysaccharide moieties in FG-GL-CP-III contribute to the enhanced immunomodulatory activity of fermented ginseng.
Results are expressed as mean±SD (n=4).
Results in the same column with different superscript letters are significantly different at P<.05.
The present study demonstrated the enhanced immunomodulating activity of fermented ginseng with G. lucidum in solid-state culture compared with nonfermented ginseng and G. lucidum mycelium alone. After fractionation of crude polysaccharide from fermented ginseng, we identified a polysaccharide fraction that contributed to the enhanced activity of fermented ginseng. Moreover, we showed that the immunomodulatory activity of the active polysaccharide (FG-GL-CP-III) was significantly decreased by NaIO4 oxidation. The purified active polysaccharides from fermented ginseng contains neutral sugars, such as Ara, Man, Gal and Glc, and their related various neutral glycosidic linkages. Finally, component sugar and linkage changes of polysaccharides in fermented ginseng by mycelium culture may also contribute to the enhancement of immunomodulating activity of ginseng.
Discussion
The root of ginseng (P. ginseng C.A. Meyer) has been widely prescribed and intensively studied as a medicinal herb in East Asia for over 2000 years. There are two traditional preparations of ginseng: white ginseng (WG) and red ginseng (RG). WG is the dried root of ginseng, and RG is the steamed and dried root of ginseng. Nam 26 reported that the heat processing improved gastrointestinal absorption by gelatinization of starch. Beak et al. 27 reported that the heat processing also induced structural changes that might enhance its biological activities and effectiveness. However, heating also degrades heat-labile malonyl-ginsenosides into corresponding neutral ginsenosides 28 and decreases the essential amino acid such as Lys by Maillard reaction.
In recent years, solid-state culture has received much interest since several studies have demonstrated that this process may lead to higher yields and productivities or better product characteristics than submerged fermentation. 29 In addition, due to the utilization of low cost agricultural and agro-industrial residues as substrates, capital and operating costs are lower compared to submerged fermentation. The low water volume in solid-state culture has also made a large impact on the economy of the process mainly due to smaller fermenter-size, reduced downstream processing, reduced stirring, and lower sterilization costs. 30,31 Although many bioactive compounds are still produced by submerged fermentation, in the past decades, there has been an increasing trend toward the utilization of the solid-state culture technique to produce these compounds since this process has been shown to be more efficient, producing higher yield than submerged fermentation. 31 Our present data suggest that solid-state culture of ginseng with G. lucidum mycelium enhances the immunological activity of fresh ginseng (Table 1), probably due to modified component contents in fermented ginseng (Table 2). These results also suggest the possibility that solid-state culture with G. lucidum mycelium can enhance the immunomodulatory activity of ginseng.
The ginseng roots have been valued for their important biological and chemical compounds and because of their use in the treatment of a variety of pathological conditions and illnesses. Historically, most interest has been focused on the ginsenosides to explore the pharmacological mechanisms of ginseng. 32 –34 However, it has been difficult to make significant progress in elucidating the underlying mechanism by which ginseng take effect only by ginsenoside. Because ginseng contains multiple constituents, such as ginsenosides, essential oil, polysaccharides, and peptides, 35,36 more recent attentions have been focused on ginseng polysaccharides due to their various important biological activities. 37,38 Therefore, the aim of present research was to elucidate the chemical constituents and biological effect of polysaccharides from fermented ginseng to help us to take for the development of functional food material.
As a white-rot medicinal fungus, G. lucidum has been used as a remedy to promote health and longevity 39 and has many interesting bioactivities, including antitumor and immunomodulating effects. 40,41 Bioactive compounds from fruiting body and mycelia of G. lucidum include polysaccharides, triterpenoids, steroids, alkaloids, nucleotides, lactones, and fatty acids. 39 Because of its perceived health benefits, G. lucidum has gained wide popularity as a health food in Korea, China, and Japan. In addition, G. lucidum could use various substrates for mycelial growth and metabolite production. 10 However, few reports have paid attention to the changes in the pharmacological activity and components of ginseng by the fermentation with G. lucidum. In this work, ginseng was fermented with the basidiomycete G. lucidum, and the changes in the immunomodulating activity and polysaccharides during the fermentation were examined. Since the middle of the last century, many studies had been conducted on the purification, structural analysis, and bioactivities of polysaccharides from ginseng.
Many biological active polysaccharides have been isolated from the roots of ginseng. Modern pharmacological studies have shown that ginseng polysaccharides had immunomodulation, antitumor, antiadhesive, antioxidant, hypoglycemic activities, and so on. 36,42,43 A total of 18 pharmacological active polysaccharides from ginseng roots were identified, and these polysaccharides consisted mainly of neutral sugar containing Ara, Gal, Rha, and Glc and uronic acid with various molar ratios depending on the origin of ginsengs. 44 –46 Previous structural analysis indicated that ginseng polysaccharides include arabinogalatan, starch-like glucan, and acidic pectic polysaccharide with molecular weight of 3500–160,000. Our component sugar analysis showed that polysaccharides (NG-CP-II or III) from nonfermented ginseng were similar to those reported (Table 4). Previously, Zhang et al. 47 reported that ginseng polysaccharides contained Man with Ara, Rha, Gal, and Glc. Although the purified active FG-GL-CP-III-1 from fermented ginseng also contained all of these neutral sugars, uronic acid abruptly decreased and Man increased compared to nonfermented ginseng (Table 4), showing that solid-state culture with mycelium affected ginseng composition. Finally, methylation analysis revealed that the purified active polysaccharide (FG-GL-CP-III-1) from fermented ginseng contained unusual linkages, such as terminal or 4-linked Man, 4-linked Gal, compared to general ginseng polysaccharides (Table 5). 44 –47 We are currently investigating whether changes in component sugars and linkages of polysaccharide in fermented ginseng result from simple mycelium attachment by mycelium growth, bioconversion by mycelium, or its secreted enzymes.
Footnotes
Acknowledgment
This work was supported by a grant of overseas-dispatched research professorship from the Korea National University of Transportation, Republic of Korea (2013). The authors are thankful for its financial support.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.