
Abstract
The ethanol extract of fermented soybean from Glycine max (chungkookjang, CHU) has been claimed to have chemopreventive and cytoprotective effects. In the present study, we examined the inhibitory effect of CHU on inducible nitric oxide synthase (iNOS) and cytokine induction by toll-like receptor (TLR) ligands treatment and attempted to identify the responsible active components. Nitric oxide (NO) content and iNOS levels in the media or RAW264.7 cells were measured using the Griess reagent and real-time polymerase chain reaction assays. CHU treatment inhibited NO production and iNOS induction elicited by lipopolysaccharide (LPS, TLR4L) in a concentration-dependent manner. Tumor necrosis factor-α and interleukin-6 productions were also diminished. Peptidoglycans (TLR2/6L) and CpG-oligodeoxynucleotides (TLR9L) from CHU inhibited iNOS induction, but not poly I:C (TLR3L) or loxoribine (TLF7L). The anti-inflammatory effect resulted from the inhibition of nuclear factor-kappa B (NF-κB) through the inhibition of inhibitory-κB degradation. Of the representative components in CHU, specific oligopeptides (AFPG and GVAWWMY) had the ability to inhibit iNOS induction by LPS, whereas others failed to do so. Daidzein, an isoflavone used for comparative purposes, was active at a relatively higher concentration. In an animal model, oral administration of CHU to rats significantly diminished carrageenan-induced paw edema and iNOS induction. Our results demonstrate that CHU has anti-inflammatory effects against TLR ligands by inhibiting NF-κB activation, which may result from specific oligopeptide components in CHU. Since CHU is orally effective, dietary applications of CHU and/or the identified oligopeptides may be of use in the prevention of inflammatory diseases.
Introduction
A
An excess amount of nitric oxide (NO) amplifies inflammation by increasing reactive free radicals such as peroxynitrite, promoting cell injury. 7 In the inflammatory cascade, inducible nitric oxide synthase (iNOS) induction results in a large amount of NO production in macrophages and other inflammatory cells. 8 In addition, tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), cytokines produced commonly by the activation of nuclear factor-kappa B (NF-κB), also participate in the inflammatory process and other immunological effects (e.g., acute phase responses). 9 Toll-like receptors (TLRs), type I transmembrane glycoproteins, have extracellular transmembrane domains with repeated leucine-rich repeat (LRR) modules and Toll/interleukin-1 receptor (TIR) domains. 10 Activators of certain TLRs initiate the inflammatory cascade with the regulation of the innate immune system. 11 Identification of TLRs and intervention pathways affected by food components would be of assistance in understanding the immunomodulatory effects of nutrients. Of the 13 mammalian TLRs, TLR4 activation by lipopolysaccharide (LPS) leads to strong induction of iNOS in macrophages and other cells. 12
In view of the significance of iNOS and cytokines in the detrimental cascade of inflammation, this study investigated the potential of CHU to inhibit iNOS, TNF-α, and IL-6 production using a macrophage cell model. In this study, we report that CHU inhibits the induction of iNOS elicited by LPS (TLR4 ligand), peptidoglycan (PGN, TLR2/6 ligand), or CpG-oligodeoxynucleotide (CpG-DNA, TLR9 ligand). Since NF-κB plays a key role in the induction of iNOS and other proinflammatory genes downstream from TLRs, 13 –15 we were interested in the effect of CHU on NF-κB activity. Our finding indicates that CHU has an anti-inflammatory effect against TLR ligand activation by inhibiting inhibitory-κB (I-κB) degradation. Also, we attempted to identify active components responsible for the anti-inflammatory activity of CHU. Finally, the anti-inflammatory effect of CHU was verified using a rat paw edema model. Since CHU soybean extract is used as plant-derived food and/or food supplement, our findings offer key information in understanding the anti-inflammatory and/or antioxidative effect of the fermented soybean food, chungkookjang.
Materials and Methods
Materials
Anti-I-κBα antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-murine iNOS antibody was obtained from BD Biosciences (San Jose, CA, USA). Peptidoglycan, poly I:C, LPS, loxoribine, CpG-DNA, daidzein, and anti-actin antibody were purchased from Sigma (St. Louis, MO, USA).
CHU preparation
CHU was prepared from soaked soybeans (Asan, Korea), as described previously. 16 Soybeans soaked for 18 h were autoclaved at 120°C for 30 min. Bacillus licheniformis B1 (1% in culture) was added onto 1 kg of autoclaved soybeans. CHU fermentation was performed by incubating the product at 40°C for 3 days. 1 The fermented soybean powder was freeze-dried, suspended in 80% ethanol, and stored at 25°C overnight. The sample was centrifuged at 15,000 g for 30 min at 4°C. The supernatant was evaporated in a vacuum, and the preparation was freeze-dried to make a powder (300 mg from 1 kg of starting material). The peptides present in CHU extract were analyzed and quantified using HPLC. Of them, the new oligopeptides, including AFPG (OP#1), GVAWWMY (OP#2), KP (OP#3), and QK (OP#4) were custom-order synthesized from Peptron, Inc. (Daejeon, Korea) and were used for cell-based assays.
MTT assay
The cells were plated at a density of 1×105 cells per well in a 48-well plate to measure the degree of cell survival. After treatment, viable cells were stained with MTT as previously described. 17
NO production
NO production was monitored by measuring the nitrite content in the culture medium. The assays were done by mixing samples with Griess reagent (1% sulfanilamide, 0.1% N-1-naphthylenediamine dihydrochloride, and 2.5% phosphoric acid). Absorbance was determined at 540 nm after incubation for 10 min.
Immunoblot analysis
SDS-polyacrylamide gel electrophoresis and immunoblot analyses were performed according to previously published procedures. 17 Proteins of interest were visualized using an ECL chemiluminescence detection kit (Amersham Biosciences, Buckinghamshire, United Kingdom). Equal loading of proteins was verified by actin immunoblotting.
Real-time polymerase chain reaction assays
Total RNA isolation and real-time polymerase chain reaction were done as previously described. 17 The relative levels of mouse iNOS mRNA (sense: 5′-CCTCCTCCACCCTACCAAGT-3′, antisense: 5′-CACCCAAAGTGCTTCAGTCA-3′, 199 bp), mouse TNF-α mRNA (sense: 5′-TACTGAACTTCGGGGTGATCGGTCC-3′, antisense: 5′-CAGCCTTGTCCCTTGAAGAGAACC-3′, 295 bp), or mouse IL-6 mRNA (sense: 5′-TTCCATCCAGTTGCCTTCTT-3′, antisense: 5′-ATTTCCACGATTTCCCAGAG-3′, 170 bp) were normalized based on that of glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
Stable transfection and iNOS-promoter luciferase assay
Firefly luciferase construct, pGL-miNOS-1588, containing murine iNOS promoter from −1588 to +165 bp was constructed as previously described. 18 For luciferase assay, cells were plated at a density of 7×105 cells/well in six-well plates overnight, serum-starved for 12 h, and then exposed to 0.1 mg/L LPS in the presence or absence of the agent of interest for 18 h. After removing the medium, passive lysis buffer® (Promega, Madison, WI, USA) was added to the cells. Firefly luciferase activities in lysates were measured using a luminometer (Berthold Technologies, Bad Wildbad, Germany). The relative luciferase activity was determined by normalizing iNOS reporter activity to that of total protein content.
NF-κB transient transfection and reporter gene assay
The cells were transiently transfected with NF-κB×5-LUC luciferase reporter plasmid (0.5 μg) for 3 h in the presence of Lipofectamine® reagent. The activity of luciferase was measured as described above. Transfection efficiency was normalized using lacZ.
Enzyme-linked immunosorbent assay
RAW264.7 cells were preincubated with CHU for 1 h and were continuously incubated with LPS (or other TRL ligand) for 6 h (TNF-α) or 18 h (IL-6). TNF-α and IL-6 levels in the culture medium were measured using the enzyme-linked immunosorbent assay (ELISA) kits purchased from Thermo Fisher Scientific (Waltham, MA, USA).
Animal treatment
Animal studies were conducted in accordance with the institutional guidelines for the care and use of laboratory animals. Rats were obtained from Charles River Orient (Seoul, Korea), and were acclimatized for 1 week in a clean room at the Animal Center for Pharmaceutical Research, Seoul National University. CHU was orally administered to rats (n=4) at a daily dose of 100 mg/kg body weight for 3 consecutive days. At 6 h after the last dose of CHU treatment, 50 μL of carrageenan dissolved in 0.9% saline (1% m/v) was injected into the right hind paw of rats. 19 Paw swelling volumes were measured at 30, 60, and 120 min after carrageenan administration. After the measurements of swelling volumes, the homogenates of right hind paw were used for immunoblotting.
Statistical analysis
For animal experiment and cell-based assays, data represent the mean±standard error means. Data were compared among groups using analysis of variance and/or Student's t-test. P<.05 was considered to be statistically significant.
Results
CHU inhibition of iNOS, TNF-α, and IL-6 induction by LPS
First, we determined whether CHU has the ability to inhibit iNOS induction by LPS (TLR4 ligand). Since CHU had no effect on the viability of RAW264.7 cells, at least up to 2 g/L in MTT assays (data not shown), we used the concentration range from 0.03 g/L through 1 g/L. In the dose-response study, CHU treatment significantly diminished LPS induction of iNOS, which agrees with decreased accumulation of NO in the culture media (Fig. 1A). Quantitative reverse transcription-polymerase chain reaction assays indicated that the level of iNOS mRNA was significantly increased 12 h after LPS treatment, which was also prevented by CHU (Fig. 1B). A promoter reporter analysis confirmed that CHU inhibited LPS induction of pGL-iNOS-1588, which contained the luciferase structural gene downstream of the murine iNOS promoter from −1588 to +165 bp region (Fig. 1C). All of these results show that CHU effectively inhibits the transcriptional induction of iNOS gene by LPS.

The effect of CHU on iNOS induction by LPS.
We then determined the effect of CHU on the induction of representative proinflammatory cytokines. In the ELISA assays, treatment of RAW264.7 cells with LPS increased the production of TNF-α and IL-6 in the media, but it was substantially diminished by CHU pretreatment (Fig. 2). Our finding that CHU inhibited toxic cytokine production further strengthens the anti-inflammatory effect of CHU against the LPS challenge.
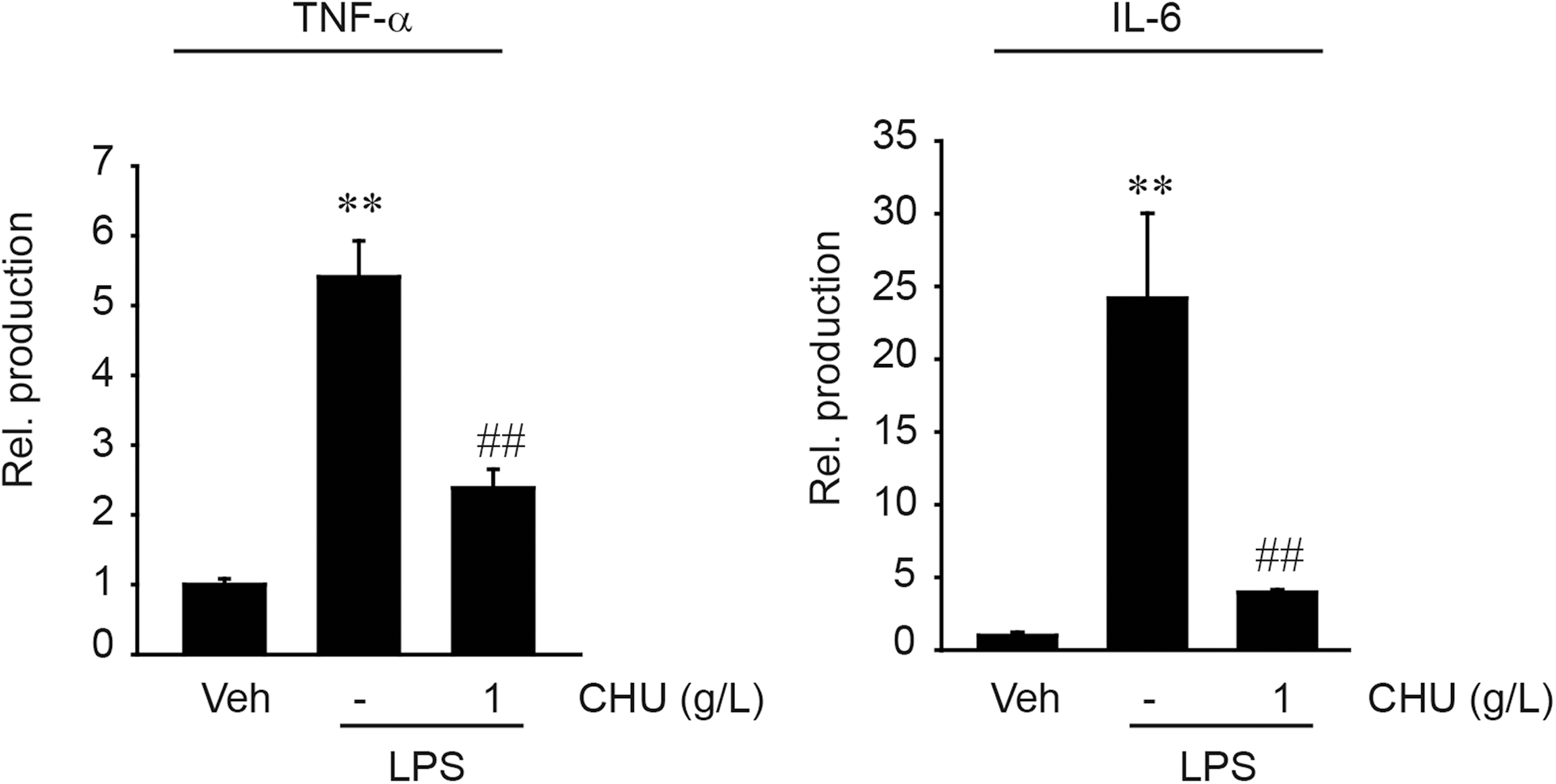
The effect of CHU on TNF-α and IL-6 production by LPS. TNF-α and IL-6 contents were measured in culture media of RAW264.7 cells treated with 1 g/L CHU for 1 h and continuously exposed to 0.1 mg/L LPS for 6 h (TNF-α) or 18 h (IL-6). Data represents the mean±SEM of three replicates (significant as compared with vehicle-treated control, **P<.01; or LPS treatment alone, ## P<.01). IL-6, interleukin-6; TNF-α, tumor necrosis factor-α.
CHU inhibition of NF-κB activation
LPS and other inflammatory stimuli activated NF-κB, thereby mediating transcriptional induction of the proinflammatory gene expression. 20 Treatment of RAW264.7 cells with LPS resulted in a significant increase in luciferase expression from NF-κB reporter construct, which was completely abrogated by CHU treatment (Fig. 3A). A previous study from this laboratory showed that LPS activated NF-κB with I-κBα degradation in RAW264.7 cells (i.e., proteolytic degradation of I-κBα subunit precedes nuclear translocation of NF-κB), which reached its maximum at 30 min–1 h. 20 CHU treatment prevented I-κBα degradation induced by LPS, consistent with the inhibition of NF-κB activation (Fig. 3B).
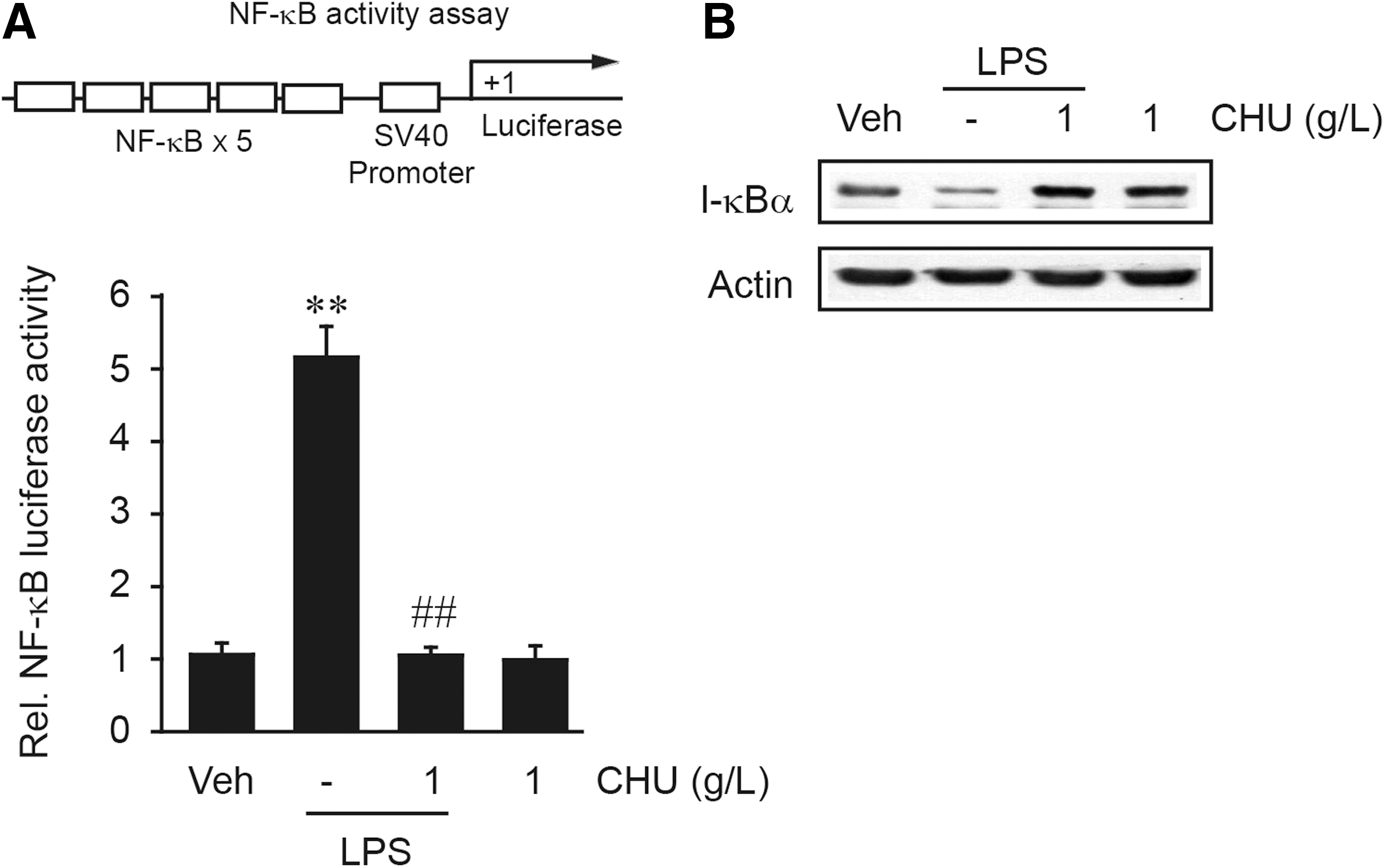
CHU inhibition of NF-κB activation by LPS.
Effect of CHU on iNOS induction by other TLR ligands
TLR2/6, TLR4, or TLR9 promote inflammatory responses through NF-κB, 21 –23 whereas TLR3, TLR7, or TLR8 do so through interferon regulatory factor family members. 24 –27 Next, we were interested in the effect of CHU on the induction of iNOS by other TLR ligands. CHU treatment prevented iNOS induction by PGN (TLR2/6 ligand) or CpG-ODN (TLR9 ligand), but not by poly I:C (TLR3 ligand) or loxoribine (TLR7 ligand) (Fig. 4). These results in conjunction with CHU effect on LPS's induction of iNOS support the contention that CHU may be active against inflammation induced by TLR2/6, TLR4, or TLR9 ligand presumably through the inhibition of NF-κB pathway.

CHU inhibition of iNOS induction by other TLR ligands. Immunoblotting for iNOS was done on the lysates of RAW264.7 cells treated with vehicle or CHU (1 g/L) for 1 h and continuously exposed to TLR2/6L (PGN, 10 mg/L), TLR3L (poly I:C, 3 mg/L), TLR7/8L (loxoribine, 100 μM), or TLR9L (CpG-DNA, 0.3 μM) for 24 h. TLR, toll-like receptor.
iNOS inhibition by oligopeptides in CHU
In an effort to find the active components comprised in CHU, we subsequently examined the effects of oligopeptides or dipeptides in CHU on the induction of iNOS and cell viability in comparison with the effect of daidzein, a representative isoflavone. Of the numerous oligopeptide fragments in CHU, four major candidates were chosen by HPLC analysis to see if they possess anti-inflammatory effects (Fig. 5A). OP#1 (Ala-Phe-Pro-Gly) treatment at the concentrations between 10 and 100 mg/L (26 and 260 μM, respectively) prevented the ability of LPS to induce iNOS in RAW264.7 cells (Fig. 5B). A similar result was obtained with OP#2 (Gly-Val-Ala-Trp-Trp-Met-Tyr) treatment. Consistently, iNOS mRNA levels and NO production were both significantly inhibited by each of the oligopeptides (Fig. 5C). In contrast, treatment with dipeptide, OP#3 (Lys-Pro) or OP#4 (Gln-Lys), or with their combined mixture (1:1) failed to do so (Fig. 5D, upper panel). However, a mixture of OP#1–OP#4 at subeffective or ineffective concentrations exerted an inhibitory effect, suggesting their synergism (Fig. 5D, middle panel). Treatment with daidzein, a known anti-inflammatory isoflavone, 28 also decreased the induction of iNOS, which occurred at a relatively higher concentration (Fig. 5D, lower panel). We also found that either OP#1 or OP#2 at the concentration of 100 mg/L slightly but significantly decreased MTT-positive cell numbers at 24 h (Fig. 5E). Because Trypan blue assays showed no cytotoxicity at the concentration used (i.e., 96–98% of vehicle control), they might slightly inhibit the growth of RAW264.7 cells. We then confirmed that both OP#1 and OP#2 at 30 mg/L were capable of inhibiting the transcriptional induction of TNF-α and IL-6 (Fig. 6). All of these results support the idea that specific oligopeptides present in CHU may contribute to the inhibition of macrophage activation.
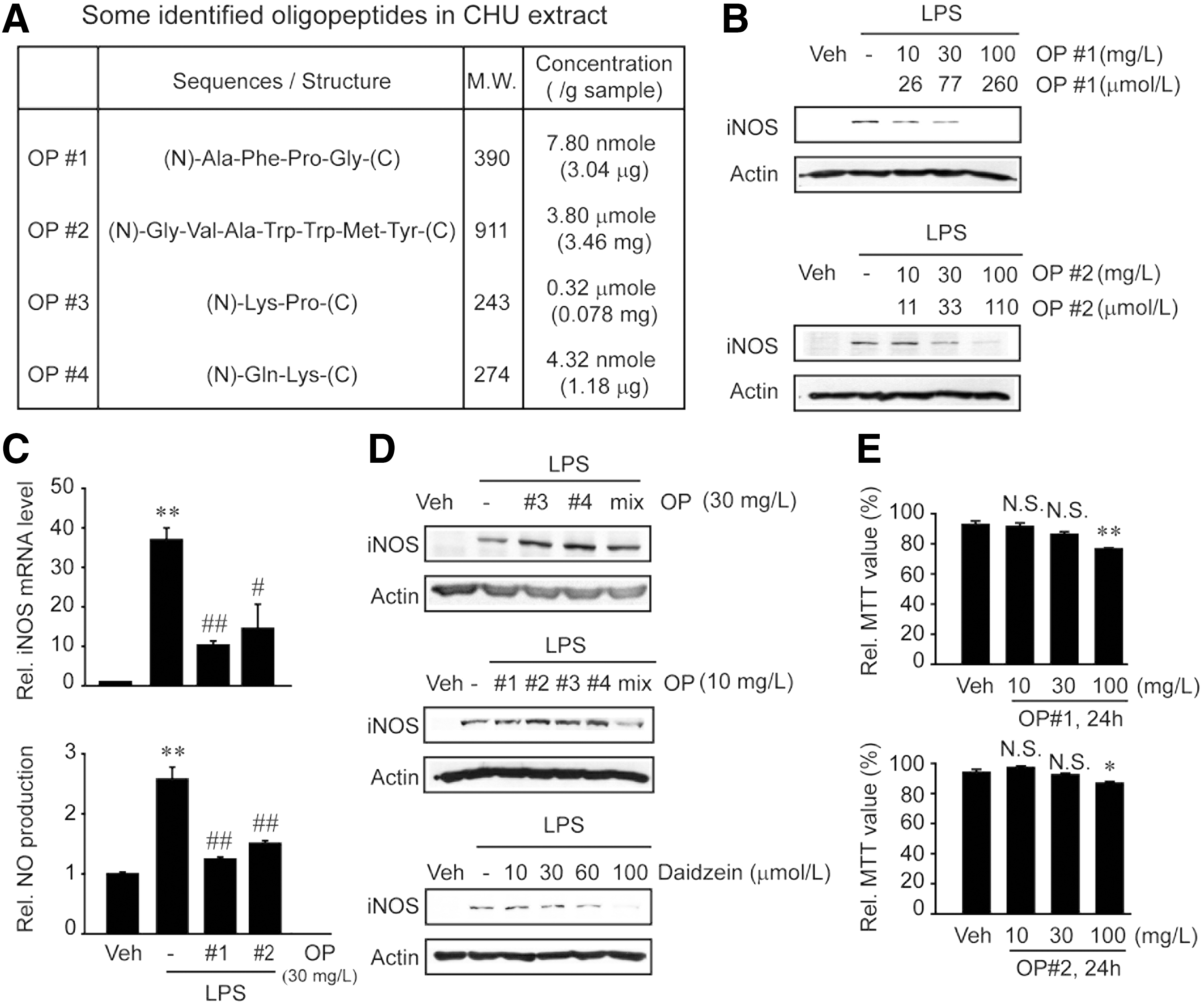
The effects of CHU components on iNOS induction and cell number.
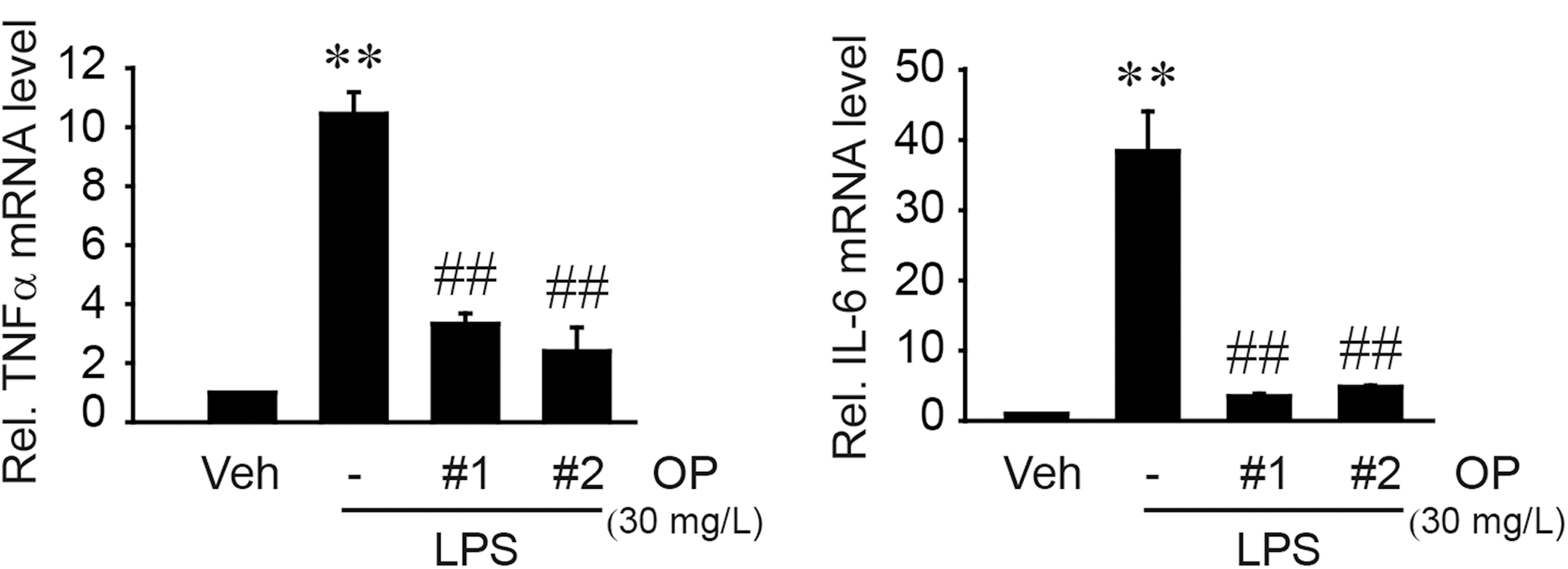
Inhibition of TNF-α and IL-6 mRNA production by OP#1 and OP#2. TNF-α and IL-6 mRNA levels were measured using qRT-PCR assays on the cells treated with either 30 mg/L OP#1 or OP#2 for 1 h, and continuously exposed to LPS for 12 h. Data represent the mean±SEM from three separate experiments (significant as compared with vehicle-treated control, **P<.01; or LPS treatment alone, ## P<.01).
Anti-inflammatory effect of CHU in an animal model
To verify the in vivo efficacy of CHU, we finally determined the effect of CHU on carrageenan-induced paw edema in rats. A daily dose of 100 mg CHU/kg body weight was chosen for this experiment because a daily dose of 30 mg CHU/kg was insufficient to achieve anti-inflammatory efficacy in rats (data not shown). As expected, an injection of carrageenan notably induced paw edema, which was significantly attenuated at 30 or 120 min by CHU pretreatment (Fig. 7A). Our data showed that CHU treatment not only delayed the onset time of paw edema, but decreased the total swelling volume as a function of time (i.e., area under the curve). Consistently, iNOS induction was diminished in the paws (Fig. 7B).
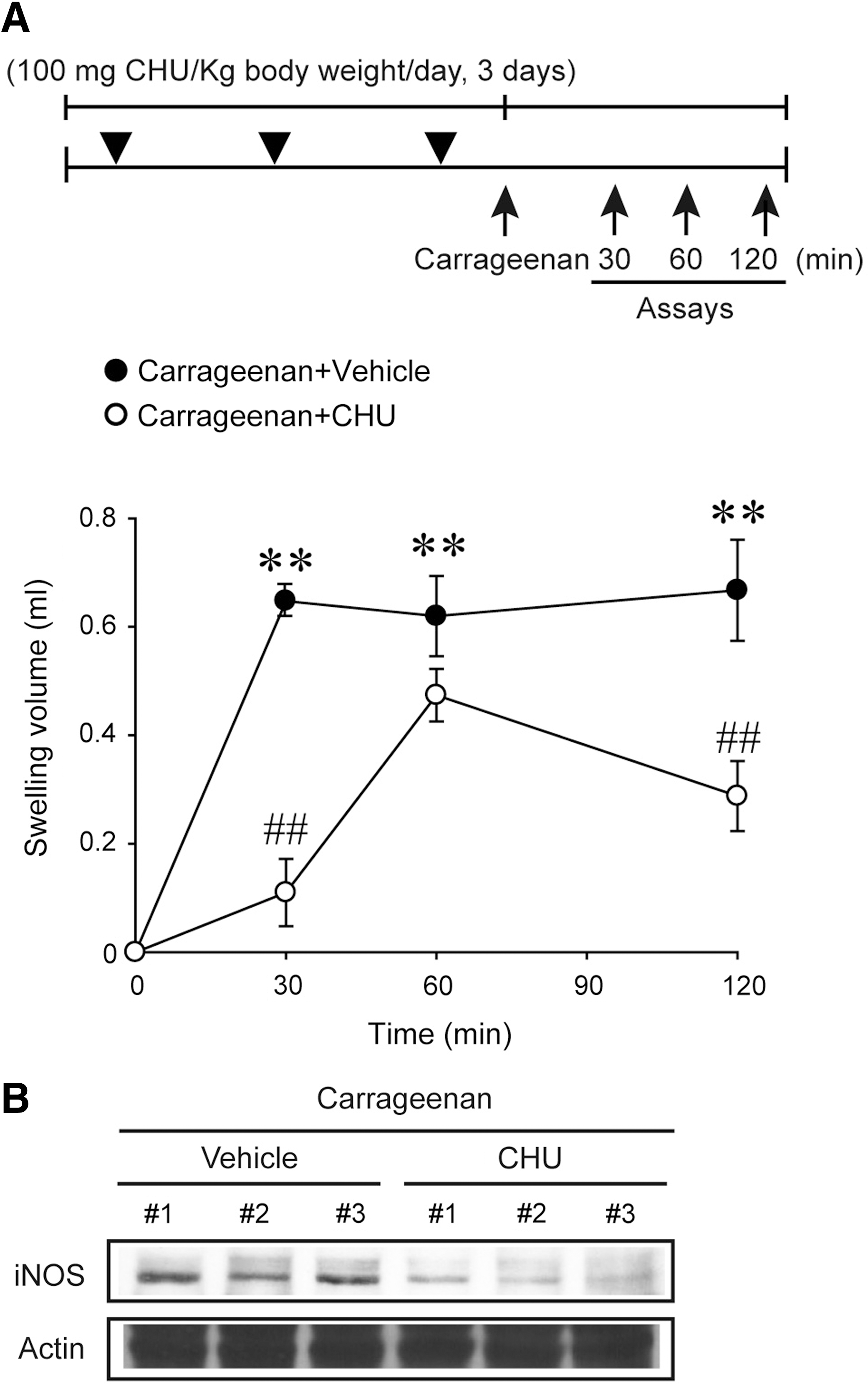
Inhibition of rat paw edema by CHU treatment.
Discussion
One of the important healthy traditional diets in Asian regions are fermented soybeans, among which CHU is distinctive as compared with other fermented soybean products as it is fermented in a short period of time without using sodium chloride. 29 The salt free status of fermented CHU makes it a health-promoting food because high-salt diets may increase the risk of cardiovascular and/or other inflammatory diseases (e.g., diabetes). 30 –32 CHU has been shown to improve blood circulation and intestinal function, and has been used for the treatment of inflammatory skin disorders, hyperlipidemia, and cardiovascular disease. 32,33 Other fermented soybean products have beneficial effects on microbiota, adding value to their anti-inflammatory activity. 34 CHU may also have a similar effect. In the present study, CHU treatment apparently inhibited NO production at relatively low concentrations: treatment with CHU at as low as 100 mg/L significantly inhibited NO production. Our data showing CHU's inhibitory effect on iNOS supports the idea that it inhibits inflammatory responses in macrophages and other related cells.
Activated immune cells remove microbes and apoptotic cells by phagocytosis and presents antigens to lymphocytes. Inflammatory response to harmful stimuli enables a host organism to cope with infection or deleterious tissue damage. Inflammation is thus closely linked to the activation of macrophages or dendritic cells, which play a role in the cascade of inflammation by producing TNF-α, IL-1, and IL-6. As a major mediator, TNF-α induces innate immune responses by activating T cells and macrophages, stimulating secretion of other cytokines. 35 Therefore, increased levels of TNF-α amplify endotoxin-induced tissue injury or other systemic effects (e.g., septic shock). 36 In the present study, CHU treatment diminished TNF-α and IL-6 production in cell-based assays, supporting the notion that the production of inflammatory mediators was suppressed.
CHU treatment decreased the level of iNOS mRNA, indicating its ability to inhibit the gene transcription. Our results showing a decrease of iNOS promoter reporter activity by CHU strengthen the ability of CHU to inhibit gene transactivation. iNOS transactivation by LPS depends on AP-1, NF-κB, and interferon regulatory transcription factor (IRF) transcription factors. 37,38 Our finding that CHU inhibited NF-κB activity with a decrease in I-κB degradation parallels transcriptional inhibition of the genes responsible for inflammatory processes and immunity.
Inflammation is provoked by the products derived from microbial pathogens (e.g., LPS and viral DNA/RNA) or apoptotic cells, and plays a critical role in acute and chronic illnesses such as hepatitis and atherosclerosis. 39,40 In fact, the substances occupy a pivotal position at the innate immune system. In particular, LPS, a prototypical endotoxin derived from gram-negative bacteria, stimulates macrophage activation and inflammatory cytokine production, facilitating progression of septic shock syndrome. 41 After stimulation of TLR4 by LPS, intracellular domain of TLR4 recruits adaptor proteins such as MYD88 and TRIF, and their interactions facilitate the activation of NF-κB and/or mitogen-activated protein kinases (MAPKs). 42 Besides TLR4, some TLRs, including TLR1/2, TLR2/6, TLR5, and TLR9 also rely on the NF-κB pathway to induce cytokines, 21 –23 whereas others (TLR3 and TLR7) lead to the production of type I IFNs through the IRF pathway. 24 –27 Our result that CHU inhibited the effect of TLR2/6, TLR4, or TLR9 ligand on iNOS induction, but not that of TLR3 or TLR7 ligand, further supports the contention that CHU has an anti-inflammatory effect by inhibiting the NF-κB pathway.
An important finding of this study is the identification of anti-inflammatory effects of OP#1 and OP#2 present in CHU: they inhibited not only iNOS induction, but also TNF-α and IL-6 production. The amount of OP#1–OP#4 in CHU was 3.04 μg/g, 3.46 mg/g, 0.078 mg/g, and 1.18 μg/g, respectively. Daidzein in CHU was 8.76 mg/g, 29 which is a weakly active concentration. Bioactive isoflavonoids such as genistein and daidzein in CHU may also promote a healthy lifestyle and lower cancer risks such as breast cancer and prostate cancer. 43,44 Our findings shown in this study indicated that daidzein treatment inhibited iNOS induction at relatively higher concentrations. Since various oligopeptides are possibly abundant in CHU due to the fermentation process, 45 the identified or other similar oligopeptides may contribute to the anti-inflammatory effect to a significant extent. We found that OP#1 and OP#2 at the concentration used slightly (∼15%) decreased RAW264.7 cell numbers with no cytotoxicity, which may have resulted from their ability to inhibit cell growth. This property remains to be studied in the future.
Overall, our results demonstrate the protective effects of CHU and the oligopeptides comprised in the extract against the TLR-mediated inflammation. Potential dietary applications of CHU and the related products may also be of value for the prevention of inflammatory diseases.
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. 2007-0056817), and, in part, by the World Class University project (R32-2012-000-10098-0) and a grant from National Research Foundation of Korea (2010-0021179).
Author Disclosure Statement
No competing financial interests exist.