
Abstract
Matrix metalloproteinase-9 (MMP-9) plays an important role in the invasion and metastasis of cancer cells. The synthesis and secretion of MMP-9 can be stimulated by a variety of stimuli, including cytokines and phorbol 12-myristate 13-acetate (PMA), during various pathological processes, such as tumor invasion, atherosclerosis, inflammation, and rheumatoid arthritis, whereas MMP-2 is usually expressed constitutively. Delphinidin, an anthocyanidin present in pigmented fruits and vegetables, possesses potent antioxidant, anti-inflammatory, and antiangiogenic properties. In this study, we investigated the antiproliferative and antiinvasive effects of delphinidin on PMA-induced MMP-9 expression in MCF-7 human breast carcinoma cells using zymography, western blotting, reverse transcription–polymerase chain reaction, and Matrigel invasion assay. Delphinidin significantly suppressed PMA-induced MMP-9 protein expression in MCF-7 human breast carcinoma cells, and it also inhibited the MMP-9 gene transcriptional activity by blocking the activation of NFkappaB (NF-κB) through MAPK signaling pathways. Moreover, the Matrigel invasion assay showed that delphinidin reduces PMA-induced cancer cell invasion. These results suggest that delphinidin is a potential antimetastatic agent that suppresses PMA-induced cancer cell invasion through the specific inhibition of NF-κB-dependent MMP-9 gene expression.
Introduction
B
Invasion and metastasis are essential properties of cancer cells. 4 These are complicated processes that involve a group of proteolytic enzymes, which participate in the degradation of tissue barriers such as the extracellular matrix (ECM) and basement membrane. ECM degradation requires the action of extracellular proteinases, of which matrix metalloproteinases (MMPs) have been shown to play an important role in breast cancer. 5 MMPs play an important role in tissue repair, angiogenesis, apoptosis, tumor invasion, and metastasis. 6 One of the members of the MMP family, MMP-9, is critical for cell migration and can mediate the invasion and metastasis of cancers.
MMP-9 specifically degrades type IV collagen, thereby facilitating tumor invasion and metastasis in breast cancer. MMP-9 also has been shown to be strongly expressed in various other cancers. The expression of MMP-9 is affected by various physical stimulators, cytokines (tumor necrosis factor-alpha, TNF-α), or the chemical, phorbol 12-myristate 13-acetate (PMA). One of these stimulators, PMA, can act as a tumor promoter that induces MMP-9 expression and other factors in certain cancer cells. 7 PMA-induced MMP-9 expression is regulated by the activation of various transcription factors. 8 For example, it modulates activator protein-1 (AP-1) 9 and nuclear factor-kappaB (NF-κB) through the activation of PKC, phosphatidylinositol 3-kinase (PI3K), and mitogen-activated protein kinase (MAPK) signaling pathways. 10 Recently, many studies have shown that active compounds from natural products 11 and food ingredients 12,13 inhibit cell migration and invasion and even metastasis of breast cancer by the suppression of MMP-9 and enzyme activity.
Delphinidin is a bioactive compound found in pomegranate and other pigmented fruits. 14 –16 Recently, delphinidin was reported to exert antitumour 17 and antioxidative activities 15 and to inhibit NF-κB. 18 However, the regulation of mechanisms underlying the expression of MMP-9, which is the main factor involved in apoptosis, invasion, and metastasis of cancer cells, by delphinidin is largely unknown. In addition, the molecular mechanisms by which delphinidin affects the expression of MMP-9 and its effect on the invasiveness of MCF-7 breast cancer cells, which are highly invasive, are still undefined. In this study, PMA-induced MCF-7 cells were used to study the effects of delphinidin on MMP-9 expression and to determine the possible mechanisms by which delphinidin reduces or prevents the expression of MMP-9 in breast cancer cells. We showed that PMA-induced MMP-9 expression was suppressed by delphinidin.
Materials and Methods
Chemical and cell culture
Delphinidin from natural plant sources was obtained from the ChromaDex. Human MCF-7 cells were obtained from the American Type Culture Collection (Rockville, MD, USA). The culture medium used in the experiments was the Dulbecco's modified Eagle's medium-low glucose (Gibco-BRL, Grand Island, NY, USA) containing 10% fetal bovine serum (Gibco-BRL) and 1% antibiotic (Gibco-BRL).
Cell viability assay
Cell viability was determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. 19 The cells were plated in 96-well plates at a density of 1×104 cells/well. Delphinidin was added at various concentrations and incubated at 37°C in a CO2 incubator. After culture for 24 h, 20 μL of the MTT reagent (2.5 mg/mL) was added to each well and incubated for 4 h, the supernatants were discarded, and the cells were dissolved in dimethylsulfoxide (DMSO). The absorption was measured at 550 nm using a microplate reader (Molecular device, Sunnyvale, CA, USA).
Matrigel invasion assay
Cell invasion was assayed as previously described 20 with some modifications, and 5×105 cells per chamber were seeded into each invasion assay. The upper insert of the transwell (Corning Costar, Cambridge, MA, USA) was coated with 30 μL of a 1:2 mixture of Matrigel:PBS. The cells were plated on the Matrigel-coated upper chamber, and the media with or without drugs were added to the upper chamber of the transwell insert. The lower chamber was filled with the culture medium. Cells in the chamber were incubated for 24 h at 37°C, and cells that had invaded the lower surface of the membrane were fixed with methanol and stained with hematoxylin and eosin. Random fields were acquired by light microscopy.
Gelatin zymography assay
The resultant supernatant was subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) containing 10% polyacrylamide and 1 mg/mL of gelatin. After electrophoresis, the gels were washed several times with 2.5% Triton X-100 for 30 min at room temperature and incubated overnight at 37°C with a buffer containing 5 mM CaCl2 and 1 M ZnCl2. The gels were stained with Coomassie Brilliant Blue R-250 (0.1% Coomassie Brilliant Blue R-250, 45.5% methanol, 9% acetic acid) (Bio-Rad Laboratories, Inc., Hercules, CA, USA) for 30 min and then destained for 1 h in a solution of 10% acetic acid and 10% methanol.
Western blot analysis
Total cell lysates were prepared using the lysis buffer (50 mM Tris, 150 mM NaCl, 5 mM EDTA, 1 mM DTT, 0.5% nonidet P-40, 100 mL phenylmethylsulfonyl fluoride, 20 mM aprotinin, and 20 mM leupeptin, pH was adjusted to 8.0) at 4°C for 30 min, followed by centrifugation at 15,000g for 5 min. The supernatant was saved as a detergent-soluble membrane fraction. The cell extract was subjected to SDS-PAGE, electrotransferred to immobilon-P membranes (Millipore, Cork, Ireland), and cytosolic and nuclear fractions were obtained using the NE-PER Nuclear and Cytoplasmic Extraction Reagents (Pierce Biotechnology, Rockford, IL, USA). Western blotting was carried out as previously reported. 21 Detection of specific proteins was carried out by enhanced chemiluminescence (Amersham Pharmacia Biotech, Piscataway, NJ, USA) following the manufacturer's instructions. The phospho-JNK, phospho-p38, and phospho-ERK antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). IκB, NF-κB, and c-Jun antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
Reverse transcription–polymerase chain reaction analysis
The total cellular RNA was extracted using TRIzol (Invitrogen Co, Grand Island, NY, USA). For reverse transcription–polymerase chain reaction (RT-PCR), a cDNA was amplified by PCR using the primers for MMP-9: forward 5′-CGGAGCACGGAGACGGGTAT-3′, reverse 5′-TGAAGGGGAAGACGCACAGC-3′; TIMP-1: forward 5′-CTGTTGTTGCTGTGGCTGATA-3′, reverse 5′ CCGTCCACAAGCAATGAGT-3′; and actin: forward 5′-GCCATCGTCACCAACTGGGAC-3′, reverse 5′-CGATTTCCCGCTCGGCCGTGG-3′. PCR products were analyzed by agarose gel electrophoresis and visualized by treatment with ethidium bromide.
Plasmid transfection and luciferase gene assays
pGL2-MMP-9-WT was used for transient transfection assays. Cells were plated onto six-well dishes at a density of 1×104 cells/mL and allowed to grow overnight. The cells were cotransfected with various plasmid constructs and the pCMA-β-galactosidase plasmid for 12 h with the DNA transfection reagent (TransIT-LT1 transfection reagent; Mirus, Madison, WI, USA) according to the manufacturer's instructions. After a 12 h incubation in fresh medium, the enzyme activities of luciferase and β-galactosidase were determined using commercial kits (Promega, Madison, WI, USA) according to the manufacturer's protocol. Luciferase activity was calculated as luciferase activity normalized to the β-galactosidase activity in each cell lysate.
Invasion assay
Cell invasion assay was performed as previously reported. MCF-7 cells to be tested for invasion were detached from the tissue culture plates, washed, and resuspended in a conditioned medium (1×105 cells/well) by using Matrigel-coated filter inserts (8 μm pore size) that fit into the 24-well invasion chambers (Becton–Dickinson, New Jersey, NJ, USA). The cells were then added to the upper compartment of the invasion chamber in the presence or absence of drugs. The conditioned medium (500 μL) was added to the lower compartment of the invasion chamber. The chambers were incubated at 37°C for 24 h in 5% CO2. After incubation, the filter inserts were removed from the wells and the cells on the upper side of the filter were removed using cotton swabs. The filters were fixed, stained, and mounted according to the manufacturer's instructions (Becton–Dickinson). The cells that invaded through the Matrigel and were located on the underside of the filter were counted. Three to five chambers were used per condition. The values obtained were calculated by averaging the total number of cells from three filters.
Statistical analysis
The results are expressed as means±SE, and the differences between means for two groups were determined by the unpaired Student's t-test. The minimum significance level was set at P<0.05 for all analysis. All experiments were performed at least three times.
Results
Delphinidin inhibits PMA-induced MMP-9 secretion of MCF-7 cells
Figure 1A shows the structure of delphinidin. We first evaluated the effect of delphinidin on the viability of MCF-7 cells by MTT assay. Delphinidin decreased the cell viability from 0% to 95% in a dose-dependent manner (Fig. 1B), and it showed 5% cytotoxicity in high concentrations toward MCF-7 cells. Based on these results, noncytotoxic concentrations of delphinidin, ranging from 0 to 60 μg/mL, were used in further experiments. Next, the effect of delphinidin on PMA-induced MMP-9 secretion was determined using the gelatin zymography method. As shown in Figure 1C, treatment with PMA for 24 h induced MMP-9 secretion in MCF-7 cells. PMA-induced MMP-9 secretion was completely inhibited in the presence of 60 μM delphinidin.

Effect of delphinidin on phorbol 12-myristate 13-acetate (PMA)-induced MMP-9 activity in MCF-7 cells. The structure of delphinidin
Next, we performed RT-PCR and promoter assay. As shown in Figure 2, the treatment of cells with delphinidin decreased PMA-stimulated mRNA level and luciferase activity in a dose-dependent manner. These results show that delphinidin suppresses PMA-induced MMP-9 secretion through inhibition of its transcriptional activity.
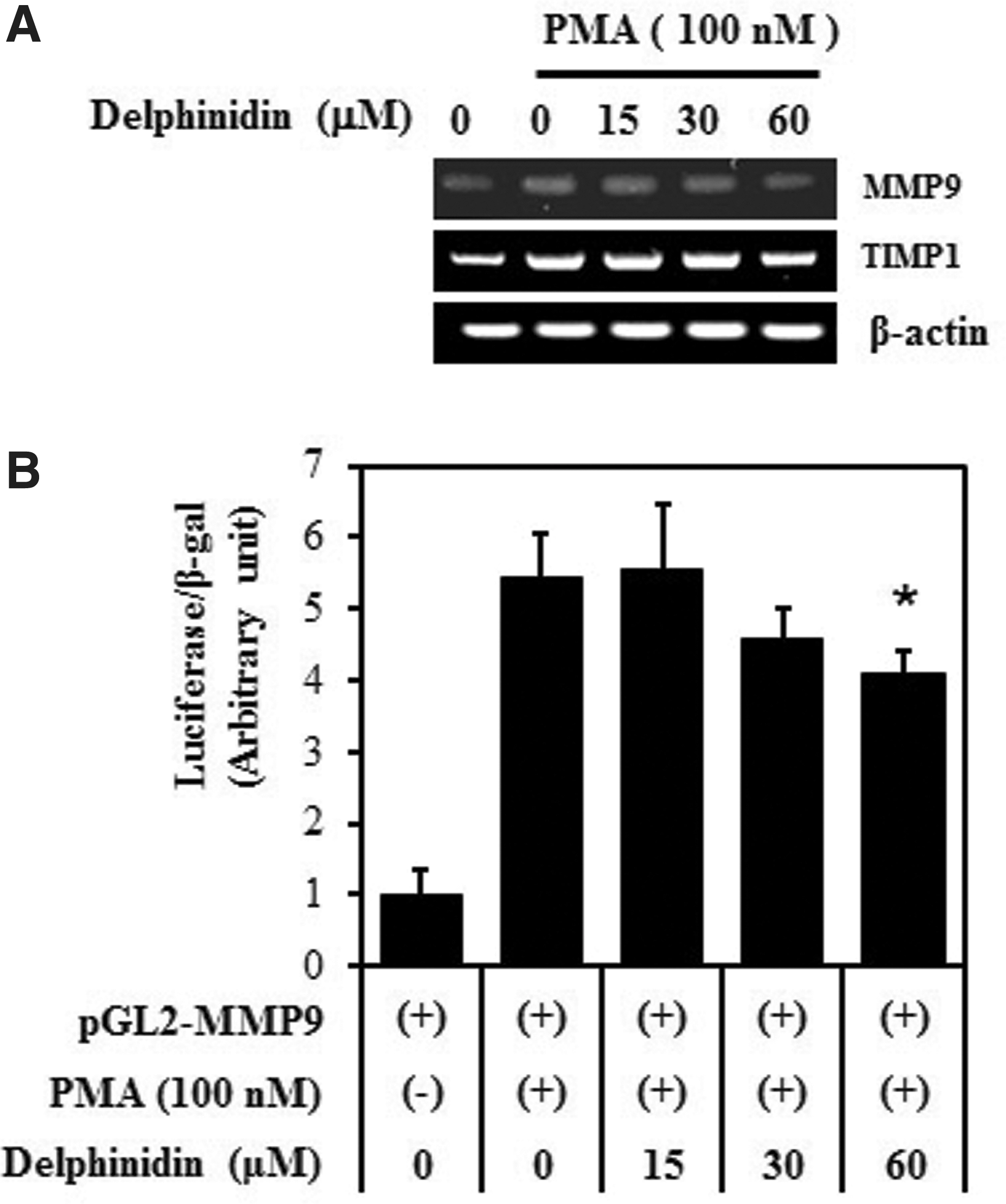
Effect of delphinidin on PMA-induced transcriptional activation of MMP-9 gene in MCF-7 cells. Cells were treated with the indicated concentrations of delphinidin for 24 h. The MMP-9, TIMP-1, and mRNA levels were measured by reverse transcription–polymerase chain reaction. β-actin was used as a control
Delphinidin inhibits PMA-induced activation of the MAP kinase pathway
MAPKs (ERK, p38, and JNK) are upstream modulators of NF-κB in the activation of MMP-9 expression. 22 We investigated the inhibitory effect of delphinidin against MAPK-induced MMP-9 expression. The effect of delphinidin on PMA-induced phosphorylation of the MAPKs was investigated. Delphinidin showed inhibitory effects on the phosphorylation of p38 and JNK, but not on the phosphorylation of ERK (Fig. 3). These results suggest that the specific inhibition of p38 and JNK pathways is directly involved in the regulation of PMA-induced MMP-9 expression by delphinidin.
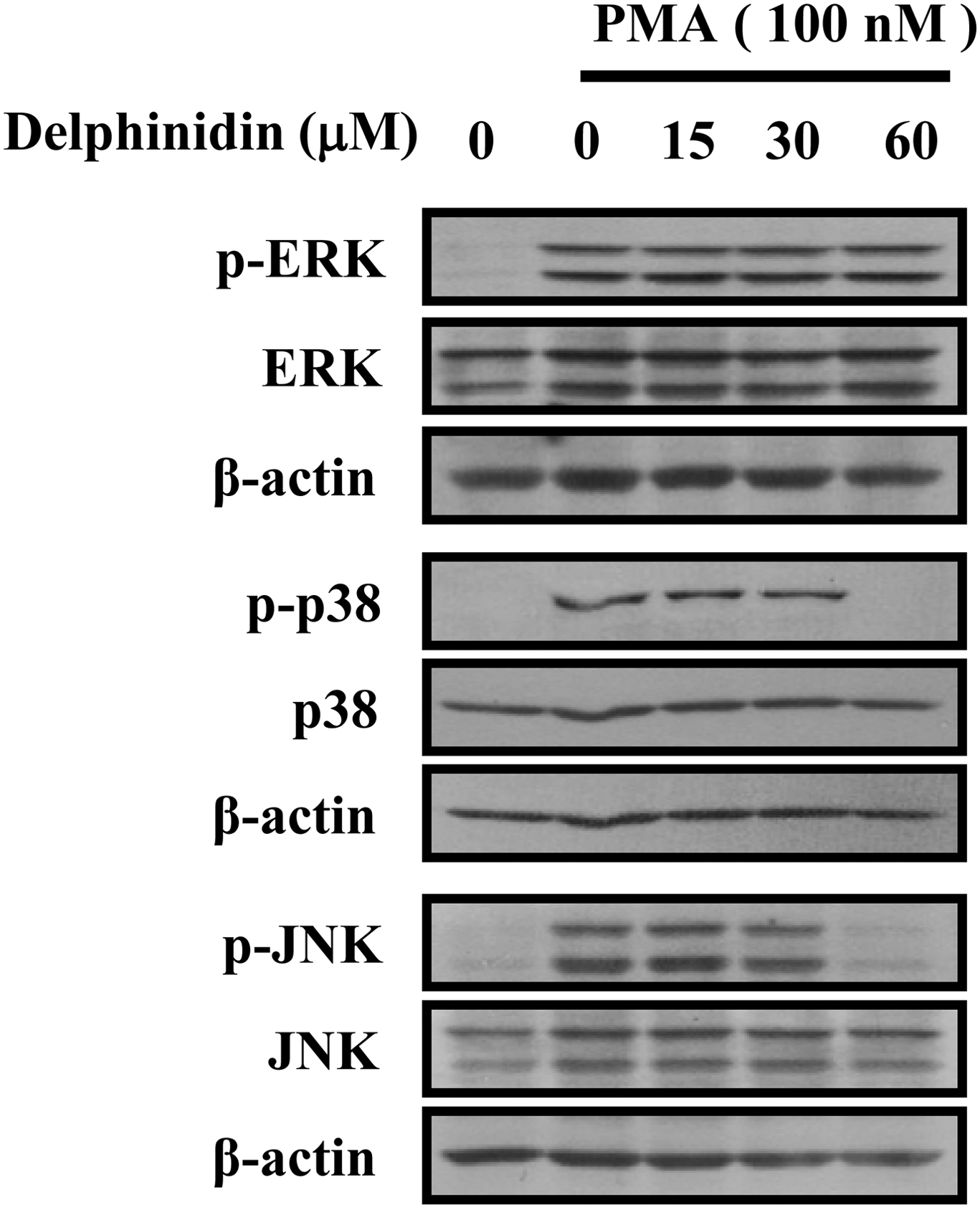
Effects of delphinidin on PMA-induced activation of MAPK signaling pathways in MCF-7 cells. Cells were treated with PMA (100 nM) for 15 min in the presence of delphinidin, and the phosphorylation levels of ERK1/2, p38 MAPK, and JNK were measured by western blotting. The results are representative of three independent experiments.
Delphinidin suppresses NF-κB and AP-1 activity
We next investigated the transcription factors that are involved in the inhibition of transcription by delphinidin in MCF-7 cells. Recently, it has been reported that MMP-9 promoter contains cis-acting regulatory elements for transcription factors that include two AP-1 sites (located at −79 and −533 bp) and an NF-κB site (located at −600 bp), and the AP-1 and the NF-κB elements are centrally involved in the PMA-induced MMP-9 expression. 13 Therefore, to determine the inhibitory effect of delphinidin on MMP-9 gene transcription through suppression of AP-1 and NF-κB, we examined the effects of delphinidin on PMA-stimulated nuclear translocation of c-fos, c-Jun (AP-1 subunits), and p65 (NF-κB subunits) through western blot analysis. As shown in Figure 4, PMA stimulated the nuclear translocation of c-Jun and p65, delphinidin inhibited the translocation of p65 in a dose-dependent manner. These results were consistent with the view that delphinidin specifically blocks NF-κB and AP-1 activation in MCF-7 cells. It is well known that NF-κB activity is regulated by IκB protein and that IκBα degradation activates NF-κB. As shown in Figure 4, PMA treatment of the cells stimulated IκBα degradation and delphinidin suppressed PMA-stimulated IκBα degradation. These results suggest that the inhibitory effect of delphinidin on PMA-induced MMP-9 expression is related to the inactivation of NF-κB through the prevention of IκBα degradation.
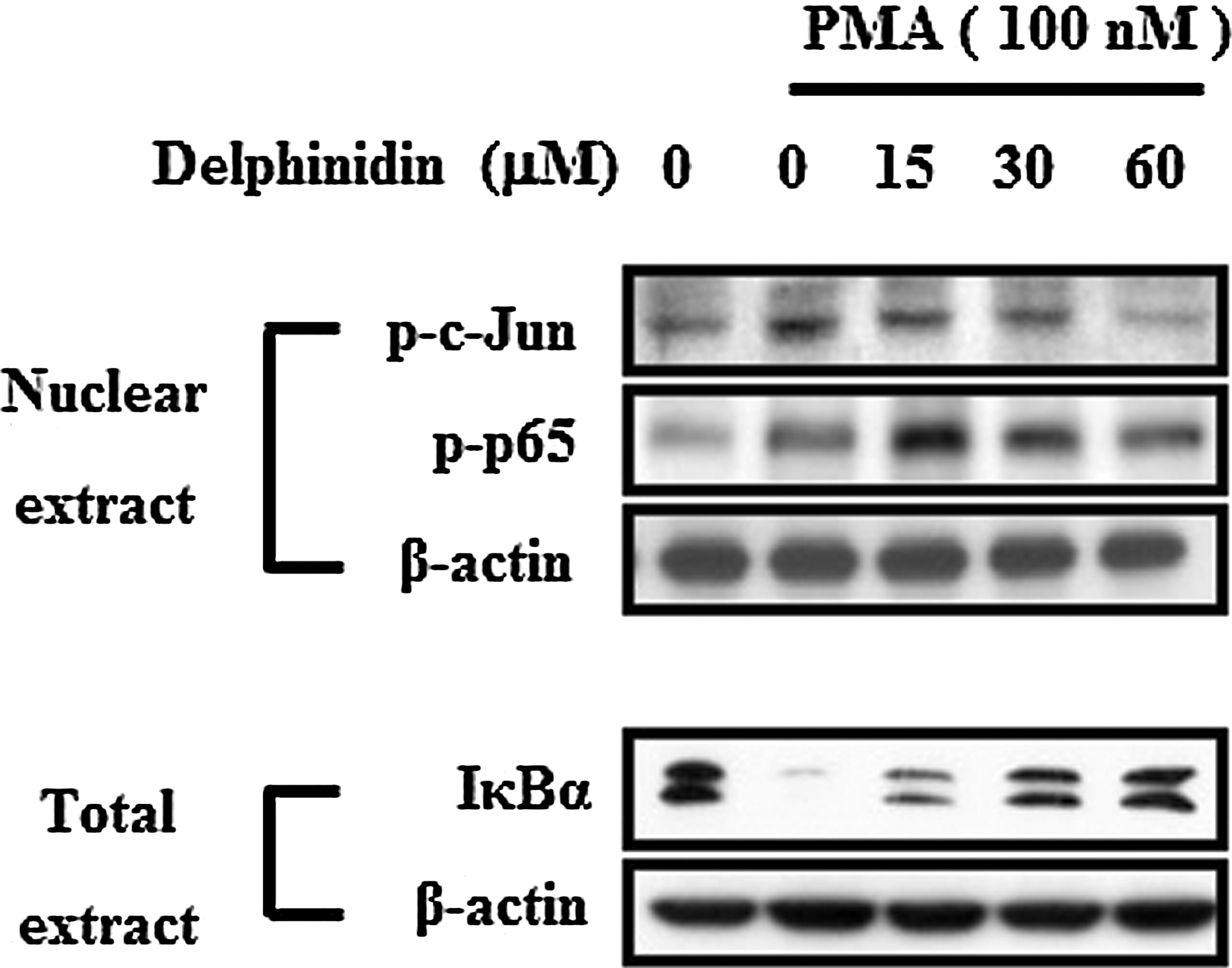
Effects of delphinidin on PMA-induced NF-κB and AP-1 activation in MCF-7 cells. Cells were treated with PMA (100 nM) for 30 min in the presence of delphinidin. Nuclear extracts were prepared and examined for phosphorylation levels of p65 and c-Jun proteins by western blotting. Whole cell lysates were subjected to western blotting using IκBα. β-actin was used as a control.
Delphinidin inhibits PMA-induced invasion and migration of MCF-7 cells
The invasion (Fig. 5A) assay showed that the invasion and migration of MCF-7 cells were increased by PMA treatment compared with that in PMA-untreated control cells. Delphinidin (60 μg/mL) almost completely inhibited PMA-induced invasion and migration of MCF-7 cells. These results suggest that delphinidin exerts antitumor effects through the suppression of PMA-induced cell invasion and migration.
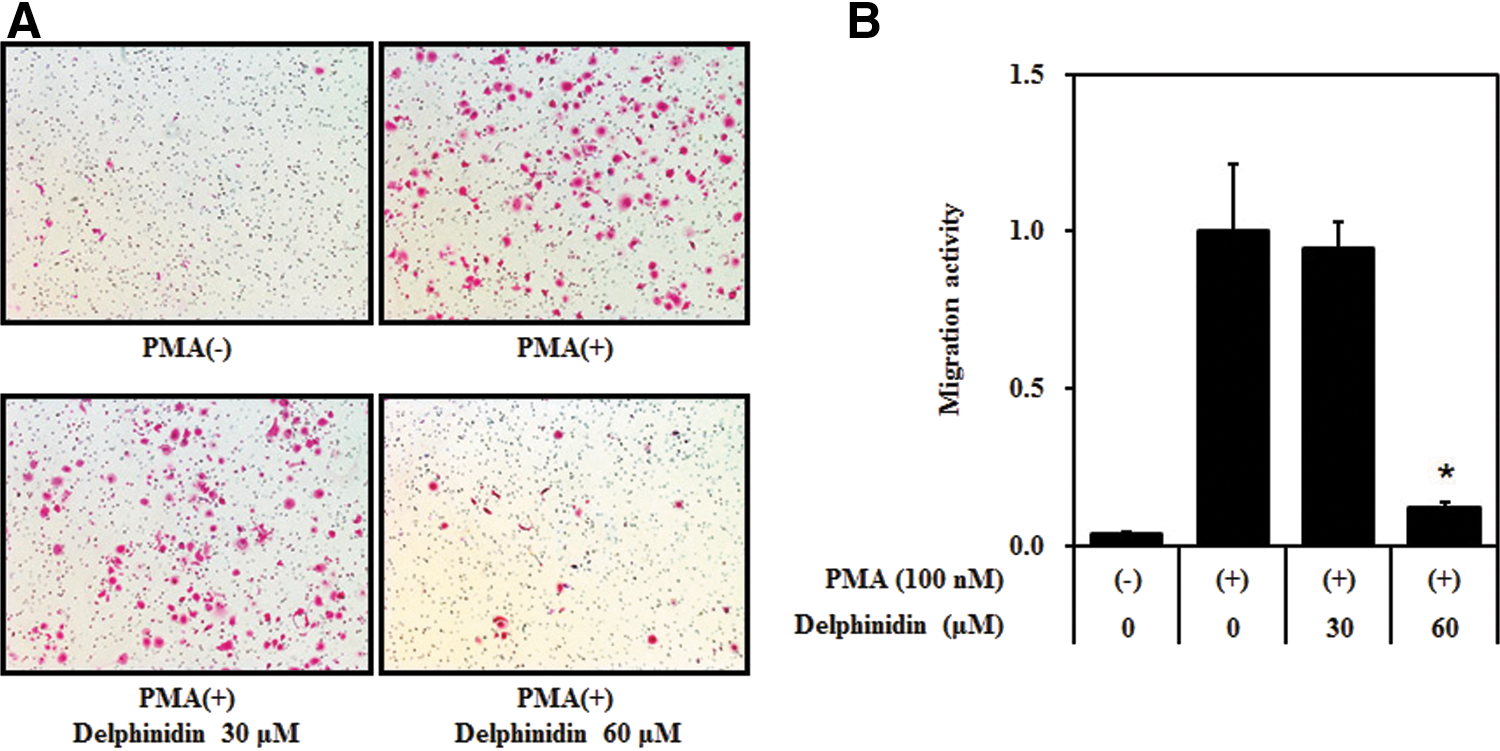
Effect of delphinidin on PMA-induced Matrigel invasion of MCF-7 cells. A Matrigel migration assay was performed with delphinidin in the presence of PMA (100 nM). After 24 h incubation, cells on the underside of the filter were fixed and stained
Discussion
Breast cancer is the main cause of cancer-related death in women worldwide and the second leading cause of death in women in the United States. 23 Tumor metastasis is the primary cause of breast cancer mortality. Metastasis is a multistep process by which a subset of individual cancer cells disseminates from a primary tumor to distant secondary organs or tissues. This process involves cell proliferation, ECM degradation, cell migration, and tumor growth in metastatic sites. 24,25 Tumor cell invasion is an early step in the metastatic cascade and represents the beginning of the transition from the benign stage to malignancy. Morphologically, tumor invasion is associated with a distorted edge of the primary tumor where individual or cohorts of tumor cells actively invade the tissue ECM surrounding the primary tumor. 26 This study demonstrated that delphinidin has the potential to decrease the invasion of MCF-7 cells and inhibits PMA-induced MMP-9 expression. It also explored the molecular mechanisms underlying this activity. We first evaluated the inhibitory effect of delphinidin on PMA-induced MMP-9 expression of MCF-7 cells. MMP-9 is a critical molecule in tumor invasion and metastasis. MMP-9 activation is associated with tumor progression and invasion, even in mammary tumors. 27 In previous reports, inflammatory cytokines, growth factors, and PMA were shown to stimulate MMP-9 by activating different intracellular signaling pathways in breast cancer cells. 28,29 The inhibition of MMP-9 expression is important for the development of therapeutic experimental models of tumor metastasis. Our data demonstrate that delphinidin inhibits PMA-induced MMP-9 expression and invasion of MCF-7 cells. PMA increases the invasion of MCF-7 cells by activating MMP-9 through transcription factors through MAPK pathways. NF-κB and AP-1 subunits are transcription factors of MMP-9 expression. Other studies have identified signal transduction pathways that are involved in regulating MMP-9 expression in tumor cells, 30 endothelial cells, 31 keratinocytes, 32 and vascular smooth muscle cells. 33 The present results showed that delphinidin markedly inhibits PMA-activated p38 and JNK MAPK pathways. Delphinidin, a purple anthocyanidin, is found in vegetables such as purple cabbage and egg plants and in fruits such as red grapes, blue berry, blackcurrants, and pomegranate. It has exhibited various biological activities in both in vitro and in vivo studies. Recently, other studies showed that delphinidin induced apoptosis of tumor cells and cell cycle progression and directly inhibited tumor growth. Especially, many researchers have reported anticancer effects in vitro and in vivo. Hafeez and their coworkers investigated delphinidin-induced apoptosis in human prostate cancer PC3 cells involving NF-κB signaling in vitro and in vivo. 17 Yun et al. reported apoptosis of human colon cancer HCT116 cells and cell cycle arrest in human colon cancer HCT116 cells. 18 A study by Bin et al. observed that decreased phosphorylation of NF-κB inhibitory protein IκBα by delphinidin induces apoptosis of both androgen-independent and androgen-refractory human PC3 cells through the activation of caspases. 14 However, antimetastatic effects in MCF-7 cells have not been reported. Therefore, we investigated the ability of delphinidin to inhibit invasion of MCF-7 cells against PMA, while focusing on its ability to counteract the activation such as MAPKs, NF-κB, and AP-1 after PMA treatment. In conclusion, the present study demonstrated that delphinidin has antimetastatic potential in PMA-treated breast cancer MCF-7 cells. Although further studies are warranted to fully elucidate the molecular mechanism underlying the antimetastatic effect of delphinidin, this study demonstrated for the first time that delphinidin suppressed PMA-induced cancer cell migration and invasion by inhibiting MMP-9 activity and expression through the NF-κB and AP-1 signaling pathways. Our results suggest that delphinidin may be a useful therapeutic agent for treating metastatic cancers and for use in pharmacological applications.
Footnotes
Acknowledgment
This research was supported by the College of Pharmacy-specialized Research Fund (from the Institute for New Drug Development) of the Keimyung University in 2013.
Author Disclosure Statement
No competing financial interests exist.