
Abstract
In this study, we found that the total polyphenol and ascorbic acid levels in the fruit of Opuntia humifusa are higher than those in other parts of the plant. We further hypothesized that antioxidants in O. humifusa might affect the growth or survival of cancer cells. Hexane extracts of seeds and ethyl acetate extracts of fruits and stems significantly suppressed the proliferation of HeLa cervical carcinoma cells, but did not affect the proliferation of normal human BJ fibroblasts. Additionally, the extracts of O. humifusa induced G1 phase arrest in HeLa cells. The O. humifusa extracts reduced the levels of G1 phase-associated cyclin D1, cyclin-dependent kinase 4 (Cdk4), and phosphorylated retinoblastoma proteins. Moreover, p21WAF1/Cip1 and p53 expression significantly increased after treatment. We examined the effects of ethyl acetate extracts of O. humifusa fruit (OHF) on HeLa cells xenograft tumor growth. OHF treatment significantly reduced tumor volume and this decrease was correlated with decreased Cdk4 and cyclin D1 expression. Furthermore, flavonoids, trans Taxifolin, and dihydrokaempferol, were isolated from OHF. Thus, this extract may be a promising candidate for treating human cervical carcinoma.
Introduction
C
Plants in the genus Opuntia are members of the Cactaceae family and are widely distributed in semi-arid countries throughout the world. 4,5 O. ficus-indica var. saboten and other Opuntia spp. have been characterized extensively at the biochemical level, 6 and their biological effects, including therapeutic properties against cancer, 7 oxidative stress, and ulcers, 8 have been well documented. However, little is known about the pharmacological properties of Opuntia humifusa. In Korea, O. humifusa has a long history of cultivation and can be grown during winter in areas where the temperatures reach −20°C or below. 9 A recent study revealed that O. humifusa displays both antioxidant and scavenging activities against the free radical 2,2-diphenyl-1-picrylhydrazyl (DPPH). Additionally, O. humifusa extracts potently modulate the expression of inflammatory cytokines. 10 Recently, water extracts from O. humifusa have been reported to exert anticancer effects on human glioblastoma or astrocytoma, epithelial-like (U87MG) cells. 11
One strategy for cancer control is chemoprevention, which is the use of dietary or synthetic agents to prevent or to slow carcinogenesis. The cell cycle in eukaryotes is regulated by a precise balance between positive and negative regulatory components that exert their effects during the G1 phase. The most critical positively acting components are G1 phase cyclins (cyclin D and cyclin E) and cyclin-dependent kinases (Cdks). 12,13 The transition from the G1 phase to the DNA synthesis (S) phase depends upon the activation of G1 phase Cdk, and phosphorylation of retinoblastoma protein (Rb). 14,15 Cdk activity and cell cycle progression are inhibited by Cdk inhibitors (CKIs), which can bind and inactivate Cdk-cyclin complexes. 16 Cdk is inhibited by p21WAF1/Cip1 and p18INK4, which ultimately blocks cell proliferation. 17,18 The inhibitory effect of p21WAF1/Cip1, which is transcriptionally regulated by p53-dependent and p53-independent pathways, is largely exerted during the G1 phase, with preferential binding to Cdk4- and Cdk2-containing complexes. 19 –21
Therefore, the first objective of this study was to investigate the antioxidant contents of O. humifusa. Based on the components of O. humifusa and its antioxidant properties, we hypothesized that extracts from the plant may have anticancer activities both in vitro and in vivo. To test this hypothesis, we investigated the inhibitory effects of O. humifusa extracts on the proliferation of HeLa cells in vitro and the growth of human cervical carcinoma in vivo. We also examined the mechanism by which the extract exerts its activity by analyzing the expression of cell cycle-regulated genes.
Materials and Methods
Antioxidant content analysis
The total polyphenol content was measured using the Folin–Ciocalteu reagent as described by Singleton and Rossi. 22 To each tube, 0.1 mL of methanol extracts was added followed by 0.4 mL of distilled water (DW) and 0.25 mL of Folin–Ciocalteu reagent. After 3 min, 1.25 mL of 20% Na2CO3 was added. The tubes were mixed thoroughly, kept for 1 h at room temperature (RT), and read at 725 nm using gallic acid as a standard.
An aluminium chloride colorimetric assay with some modifications was used for total flavonoid content. 23 Briefly, 100 μL of the diluted sample was mixed with 100 μL of 2% AlCl3·6H2O. The mixture was incubated at RT for 10 min. The mixture was vigorously shaken, and absorbance was read at 430 nm against a blank prepared with ethanol using a microplate reader (Bio-Tek, Winooski, VT, USA). Quercetin was used as a standard. Using a standard curve (0–100 μg/mL), the level of total flavonoid contents in O. humifusa were determined.
Ascorbic acids were extracted from the samples (15–20 g) with 80 mL of 5% metaphosphoric acid. Samples were filtered using a 0.45 μm membrane filter (MSI, Inc., Littleton, MA, USA); 10 μL of each sample was injected into an HPLC apparatus (PU-980; Jasco [Tokyo, Japan] and UV-detector, UV-975; Jasco) with a 250×4.6 mm Waters Spherisorb NH2 column (Waters, Milford, MA, USA) with 60 mM phosphoric acid (pH 2.0)/acetonitrile (20/80) used as eluent. Quantitative determination was based on the integrator data, which were calibrated with a pure standard of ascorbic acid (Tokyo Kasei Kogyo Co., Ltd., Tokyo, Japan). The flow rate was set at 1.0 mL/min and the temperature of the column oven at 40°C. The eluate was monitored at 254 nm. The HPLC system used for β-carotene assay was the same as that for ascorbic acid assay. The mobile phase was methanol:acetonitrile:DW (47:47:6, v/v/v) at a flow rate of 1.0 mL/min, and UV detection was performed at 325 nm. All samples were analyzed in triplicate.
Extraction of the O. humifusa
O. humifusa was collected during October in a field located in the city of Goyang, Korea. Each specimen was freeze-dried using a freeze drier (EYELA, Tokyo, Japan) at −70°C and grounded using a cyclone mill (UDY Co., Fort Collins, CO, USA). The powder from fruits (seed-free), seeds, stems, and roots of O. humifusa was extracted using methanol three times at RT; each extract was used for 2 days. Methanol extracts were subsequently filtered through Whatman No. 2 filter paper. Methanol-extracted samples were concentrated in a vacuum at 40°C. The yield was 16.4% from the fruit, 5.0% from the seed, 8.4% from the stem, and 5.9% from the root. Residues were suspended in water and then partitioned using hexane to produce a hexane-soluble layer. The aqueous layer was then partitioned using ethyl acetate to yield ethyl acetate-soluble and water-soluble residues. 24 All extracts were stored at −20°C until use.
Cell culture
HeLa cells were purchased from the Korean Cell Line Bank (KCLB, Seoul, Korea). Human fibroblast BJ cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA).Cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% (v/v) fetal bovine serum (Gemcell, Gemini, CA, USA), 1% penicillin/streptomycin (Caisson, North Logan, UT, USA), and L-glutamine (Caisson). Cells were grown at 37°C and 5% CO2 in humidified air. 25
Assay for cytotoxicity
Cell cytotoxicity was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,3 diphenyltetrazolium bromide (MTT) assay (Roche, Mannheim, Germany). This assay is used to indirectly determine cytotoxicity by measuring the activity of mitochondrial enzymes. MTT, a yellow tetrazolium salt, is metabolized by mitochondrial dehydrogenases to form blue formazan (methyltriazole). Briefly, cells were plated in 96-well plates at an initial density of 1×104 cells per well. After incubation for 24 h at 37°C, cells were treated with various concentrations of O. humifusa extracts and incubated for 24 h. Solubilized formazan products were quantified using an enzyme-linked immunosorbent assay (ELISA) reader (PowerWaveXS; Bio-Tek) at 550 nm with a reference wavelength of 690 nm. 11
Cell cycle analysis
HeLa cells were seeded into six-well plates at 3×105 cells per well. After 24 h of exposure to O. humifusa extracts, cells were harvested by trypsinization and combined with floating cells collected from the media. Cells were then fixed in 70% ethanol and resuspended in 200 μL of 100 μg/mL propidium iodide (Sigma, St. Louis, MO, USA) in phosphate buffered saline. The cell cycle stage was determined by flow cytometry using a FACSCalibur (Becton Dickinson, Franklin Lakes, NJ, USA) equipped with CellQuestPro software. 11
RNA isolation and quantitative real-time PCR
The total RNA of cells and tumor tissues was extracted using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Isolated RNA was further purified by using the RNeasy Mini Kit (Quiagen, Valencia, CA, USA) with RNase-free DNase (Quiagen) and quantified by the use of a Nanodrop spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA). Synthesis of cDNA was performed using the iScript cDNA synthesis kit (BioRad, Hercules, CA, USA). The synthesized cDNA was subjected to quantitative PCR using CFX-96 (BioRad) according to the manufacturer's specifications (KapaBiosystems, Woburn, MA, USA). The PCR cycle conditions were as follows: 95°C for 3 min, 45 cycles of 95°C for 10 sec, 61°C for 15 sec, and 72°C 15 sec. The Ct values were obtained from the BioRad CFX-Manager software (BioRad) and adjusted for GAPDH Ct values for each sample to determine ΔCt and relative expression of the target mRNA. Data were presented 2−ΔΔCt. 26 The primers used in this study included the following:
p53, forward: GGCCATCTACAAGCAGTCACAGCAC and reverse: CTCATTCAGCTCTCGGAACATCTCG
p21, forward: CCCGTGAGCGATGGAACT and reverse: CGAGGCACAAGGGTACAAGA
Cdk4, forward: TGTGGAGCGTTGGCTGTATC and reverse: TGGTCGGCTTCAGAGTTCC
Cyclin D1, forward: GCTGGAGGTCTGTGAGGAGCA and reverse: CTCCTCGGGC CGGATAGAGTT
Cdk2, forward: CCGGCTCGACACTGAGACTGAAG and reverse: GAGGAATGCCCGTGAGAGCAGA
Cyclin E1, forward: GCGCGCGCAGAGGTTTCATAG and reverse: TGCAAGGGCTGATTCCTCCAGA
Housekeeping GAPDH, forward: CTACACTGAGCACCAGGTGGTCTC and reverse: GATGGATACATGACAAGGTGCGGC.
Western blot assay
Whole cells and tumor tissues were lysed in a lysis buffer (20 mM Tris-HCl [pH 8.0], 1% NP-40, 150 mM NaCl, 2 mM EDTA, 50 mM NaF, and 1 mM Na3VO4) supplemented with protease inhibitors cocktail (Roche, Applied Science, Indianapolis, IN, USA). After incubation for 20 min on ice, lysed tissues were centrifuged, and equal amounts of protein were dissolved in 4×sodium dodecyl sulfate (SDS) sample buffer. Samples were boiled for 5 min before loading onto 10% SDS-polyacrylamide gels. After electrophoresis, proteins were transferred onto a polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA) and blocked with Tris-buffered saline/0.1% Tween 20 (TBST) containing 5% nonfat dried milk. The membrane was incubated with the appropriate primary antibody in 5% nonfat dried milk at 4°C overnight. The membrane was washed with TBST, followed by incubation with a horseradish peroxidase-conjugated secondary antibody in TBST. Bands were visualized using the Amersham ECL system (Amersham Pharmacia Biotech, Arlington Heights, IL, USA). 25
In vivo tumor xenograft study
Female Balb/c nude mice (4 weeks old) were purchased from the Animal Resources Centre (Canning Vale, Australia) and housed in a specific pathogen-free system with 12-h light–12-h dark cycles and free access to regular rodent feed (Purina rodent chow 38057; Purina Korea, Inc., Seoul, Korea) and water. The diet contained 20% protein, 4.5% fat, 3.7% fiber, 7.3% ash, and 64.5% nonfibrous carbohydrate. HeLa cells (2×106 cells) were injected subcutaneously into the flanks of nude mice. 25 Tumors were allowed to develop until they reached a size of 0.6 cm3, after which treatment (ethyl acetate extracts of O. humifusa fruits [OHF]) was initiated. Sixteen mice were randomly divided into four groups. The control group was treated with an equal volume of DW vehicle. Mice in the treatment group were orally administered 100 mg/kg body weight/day of OHF (OHF100), 200 mg/kg body weight/day of OHF (OHF200), and 500 mg/kg body weight/day of OHF (OHF500) through zonde. The OHF was made to a final volume of 300 μL. According to the human mouse equivalent dose conversion principle, 27 the experimental dosages for OHF used in this study were equivalent to the corresponding clinical prescription dosage used for a 60 kg human subject. After transplantation, the tumor size was measured using vernier calipers, and tumor volume was calculated as (L×W 2)/2, where L is the length and W is the width of the tumor. At the end of the study, after recording body weights, animals were euthanized, and the uterus, liver, spleen, lungs, and kidneys were carefully removed, washed with saline, and immediately weighed. Furthermore, tumors were harvested for analysis after tumor weights have been measured. All experimental designs and procedures were approved by the Institutional Animal Care and Use Committee of Korea University.
Column chromatography of the OHF
The OHF was subjected to a liquid chromatography (glass column [5×40 cm] packed with silica gel [230–400 mesh]) using CHCl3:MeOH (100:0, 50:1, 40:1, 30:1, 20:1, 10:1, 8:1, 6:1, 4:1, 3:1, 2:1, 1:1) gradient mixtures as eluents. The eluent fractions OHF1–OHF12 were obtained from this initial column chromatographic separation. The fractions OHF3 and OHF4 were chosen for further workup. The OHF3 was separated by the column chromatography using a glass column (1×40 cm) packed with silica gel (230–400 mesh) in hexane. The column was then eluted with hexane-EtOAc (10:0, 9:1, 8:2, 7:3, 6:4, 5:5, 4:6, 3:7, 2:8, 1:9, 0:10) resulting in seven subfractions (OHF3-1–OHF3-7). Compound 1 (10.2 mg) was isolated from OHF3-3 (48.7 mg) by preparative HPLC (column: YMC J'sphere ODS-H80, 4 μm, 250×20 mm ID, flow rate: 8.0 mL/min, 50–70% MeOH, 50 min). The fraction OHF-4 (223.7 mg) was separated by the column chromatography using a glass column (4×130 cm) packed with sephadex LH-20 gel in CHCl3: MeOH (1:1). The column was then eluted with CHCl3:MeOH (1:1) resulting in 12 subfraction (OHF4-1–OHF4-12). Compound 2 (4.2 mg) was isolated from OHF4-9 by preparative HPLC (column: YMC J'sphere ODS-H80, 4 μm, 250×20 mm ID, flow rate: 8.0 mL/min, 50–70% MeOH, 50 min).
Compounds 1 and 2 exhibited the following data
Compound 1: Yellowish powder; 1 H-NMR (CD3OD, 500 MHz) δ: 7.32 (2H, d, J=8.6 Hz, H-3′ and 5′), 6.83 (2H, d, J=8.5 Hz, H-2′ and 6′), 5.92 (1H, d, J=2.0 Hz, H-8), 5.88 (1H, d, J=2.0 Hz, H-6), 4.97 (1H, d, J=11.6 Hz, H-3), 4.54 (1H, d, J=12.1 Hz, H-2); 13 C-NMR (CD3OD, 125 MHz) δ: 198.7, 168.9, 165.5, 164.7, 159.4, 130.5, 129.5, 116.3, 102.0, 97.5, 96.4, 85.1, 73.8; ESI-MS: m/z 287.04 [M-H]−. Compound 2: Yellowish powder; 1 H-NMR (CD3OD, 500 MHz) δ: 7.06 (1H, d, J=1.8 Hz, H-2′), 6.91 (1H, dd, J=8.1 Hz, H-5′), 6.85(1H, d, J=8.1 Hz, H-6′), 5.98 (1H, d, J=2.0 Hz, H-8), 5.95 (1H, d, J=2.0 Hz, H-6), 5.01 (1H, d, J=11.7 Hz, H-3), 4.60 ((1H, d, J=11.2 Hz, H-2); 13 C-NMR (CD3OD125, MHz) δ: 197.3, 167.0, 164.1, 163.3, 145.7, 144.9, 128.9, 119.9, 115.0, 114.9, 100.6, 95.2, 95.2, 83.6, 72.3; ESI-MS: m/z 305.99 [M+H]+.
Statistical analyses
All experiments were replicated a minimum of three times. Data were presented as means±standard error. Differences between the means of individual groups were assessed by one-way analysis of variance with the Duncan's multiple range tests. The statistical software package SAS version 9.2 (SAS Institute, Cary, NC, USA) was used for the analysis. The level of statistical significance was based on the P-values (P<.05).
Results
Antioxidant contents of O. humifusa
The antioxidant contents of O. humifusa are presented in Table 1. The β-carotene content in the plant stem was 22.88 mg/100 g; no β-carotene was detected in the seed or root. The fruit and stem contained 445.40 and 260.94 mg/100 g ascorbic acid, respectively. According to these results, fruit and stem have higher amount of vitamins than other parts. As shown in Table 1, the total flavonoid levels of fruit and stem were similar, whereas total polyphenol was present at higher amounts in the fruit (5.05 mg gallic acid equivalents [GAE]/g) compared with other parts.
Values are means±standard error (n=3).
Total polyphenol contents are expressed as gallic acid equivalents (GAE). Total flavonoid contents are expressed as quercetin equivalents (QE). ND, not detected.
Extracts of O. humifusa inhibit cell proliferation in human cervical cancer cells
HeLa cells were treated with the extracts for 24 h at concentrations ranging from 100 to 1000 μg/mL and the number of viable cells was determined using the MTT-based assay. Hexane extracts of O. humifusa significantly suppressed (P<.05) cell proliferation (Fig. 1). Additionally, treatment with the ethyl acetate extracts (100 μg/mL) of the fruit, stem, and root significantly decreased the proliferation of HeLa cells (P<.05). The effects of O. humifusa extracts on cell growth were further examined using BJ cells. Treatment of the cells with the ethyl acetate extracts markedly increased the number of viable cells (Fig. 2). Compared to the control, no significant difference in viability was observed for BJ cells treated using hexane extracts (100 and 500 μg/mL) of the seed (Fig. 2). Based on these results, we used an extract concentration of 100 μg/mL in proliferation studies. Therefore, hexane extracts of seeds and ethyl acetate extracts of fruits and stems were used in subsequent experiments since they induced a reduction in HeLa cell proliferation without affecting the proliferation of BJ cells.
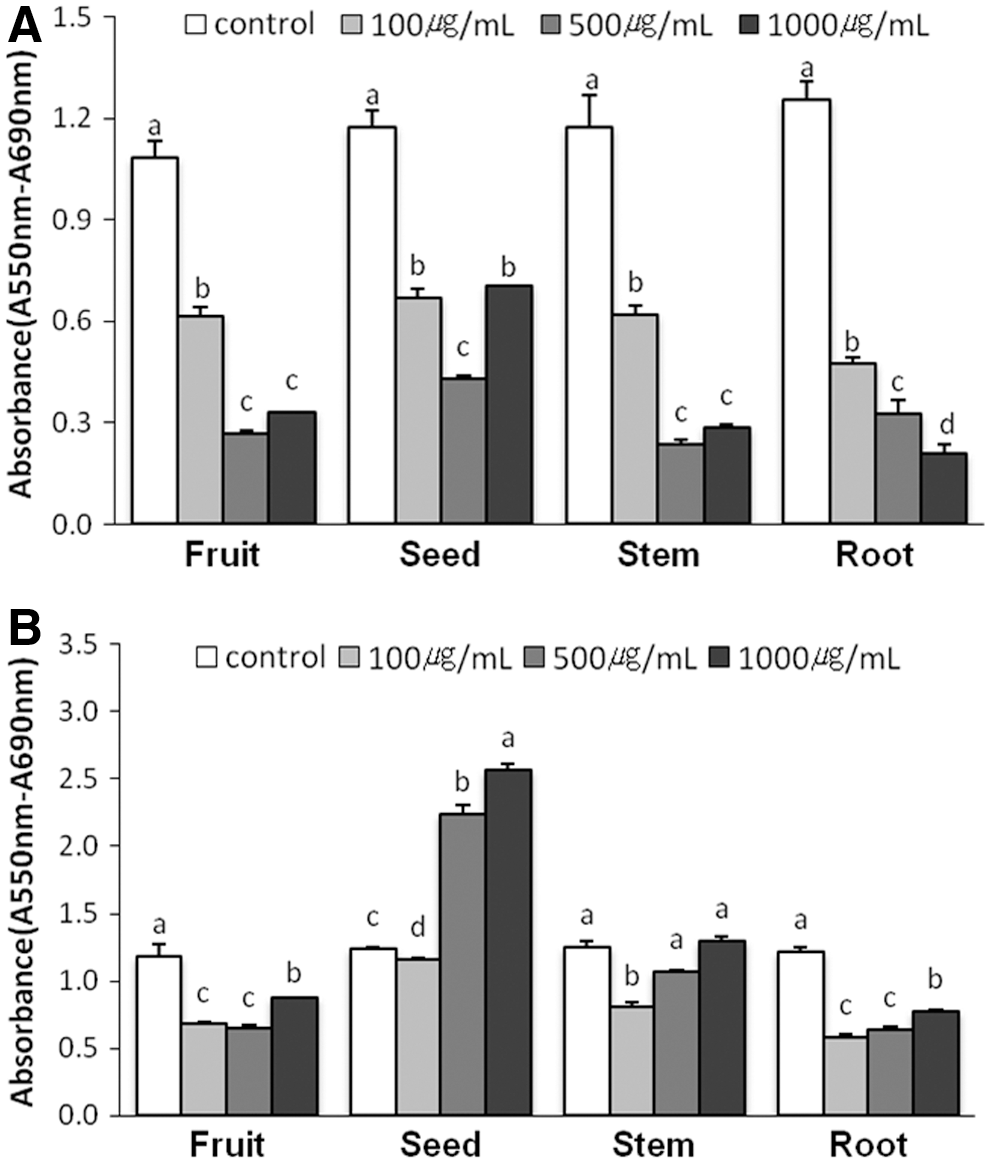
Inhibitory effect of O. humifusa extracts on the growth of HeLa cells. Cells were treated with the indicated concentration of the hexane extracts

Cytotoxicity of O. humifusa extracts toward BJ cells. Cells were treated with the indicated concentration of the hexane extracts
Extracts of O. humifusa induces G1 phase cell cycle arrest in HeLa cells
Based on the inhibitory effects of O. humifusa extract on cell growth, we investigated its effect on cell cycle progression. As shown in Figure 3, exposure of HeLa cells to seed hexane extracts, and fruit and stem ethyl acetate extracts significantly increased the number of cells in G1 phase, which was accompanied by a decrease in the number of cells in the S and G2-M phases compared with the control (P<.05). These results indicate that the inhibitory effects of O. humifusa extracts are likely due to G1 phase cell cycle arrest in HeLa cells.
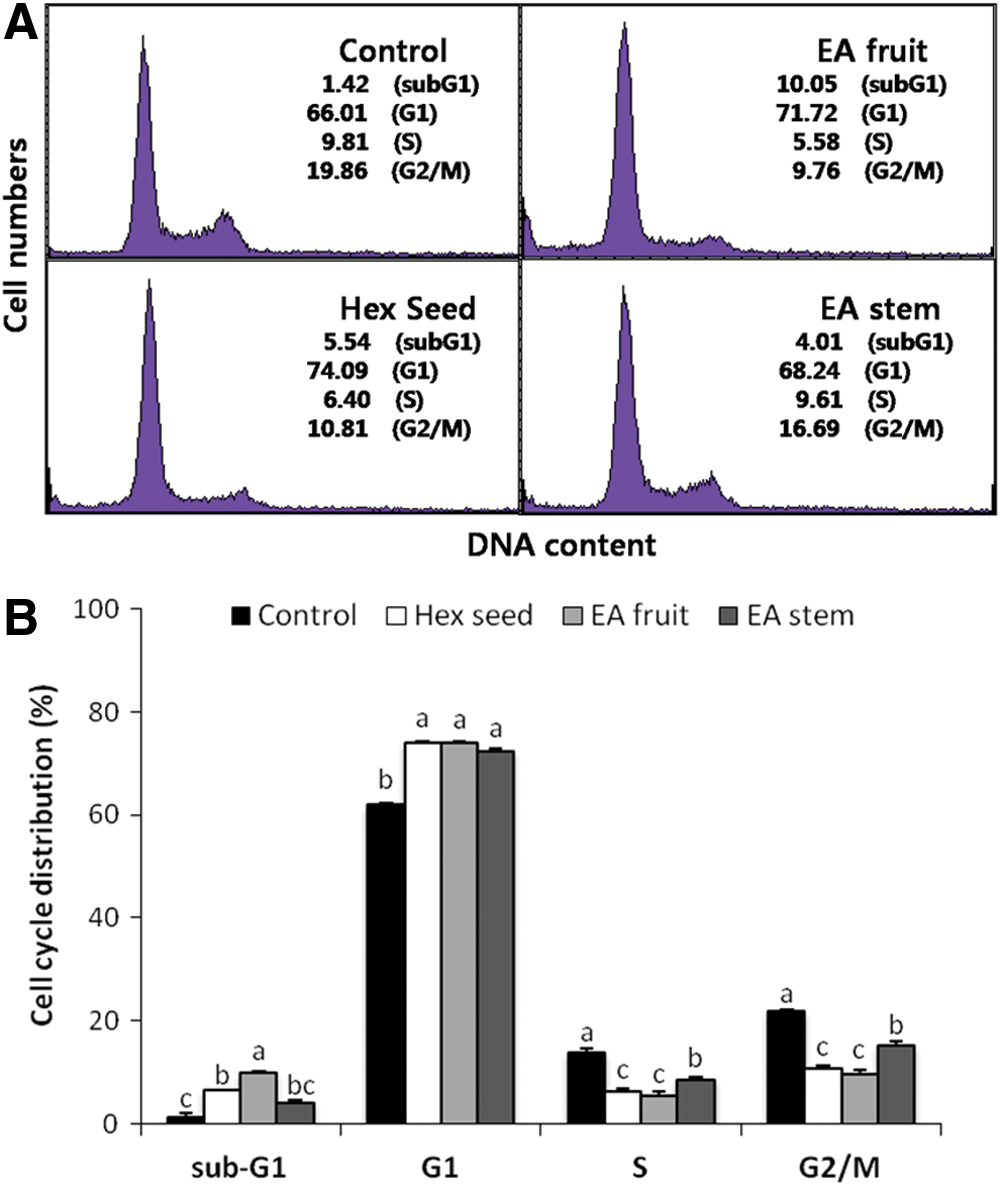
Effect of O. humifusa extracts on the cell cycle in HeLa cells.
Effects of extracts of O. humifusa on Cdks, cyclins, and CKI expression
To examine the molecular basis of extract-mediated G1 arrest in HeLa cells, we examined the expression of known G1 phase-associated proteins. As shown in Figure 4, the mRNA levels of Cdk4 and cyclinD1 both decreased significantly in cells treated with extracts, compared with the results for the control. Additionally, O. humifusa extracts reduced the protein levels of cyclinD1 and Cdk4 (Fig. 5). The phosphorylation of Rb markedly decreased after treatment (P<.05). To further examine the effect of the O. humifusa extracts on CKI protein, the expression of p18INK4 and p21WAF1/Cip1, which are important CKIs in the G1 phase, was examined. The level of both proteins increased after 24 h treatment (Fig. 5). Additionally, the p21WAF1/Cip1 mRNA level was 15 times higher in cells treated with ethyl acetate extracts of fruits compared with the control (Fig. 4). We also evaluated p53 status upon treatment and found that the extracts increased p53 expression levels in HeLa cells (Fig. 5).

Gene expression of
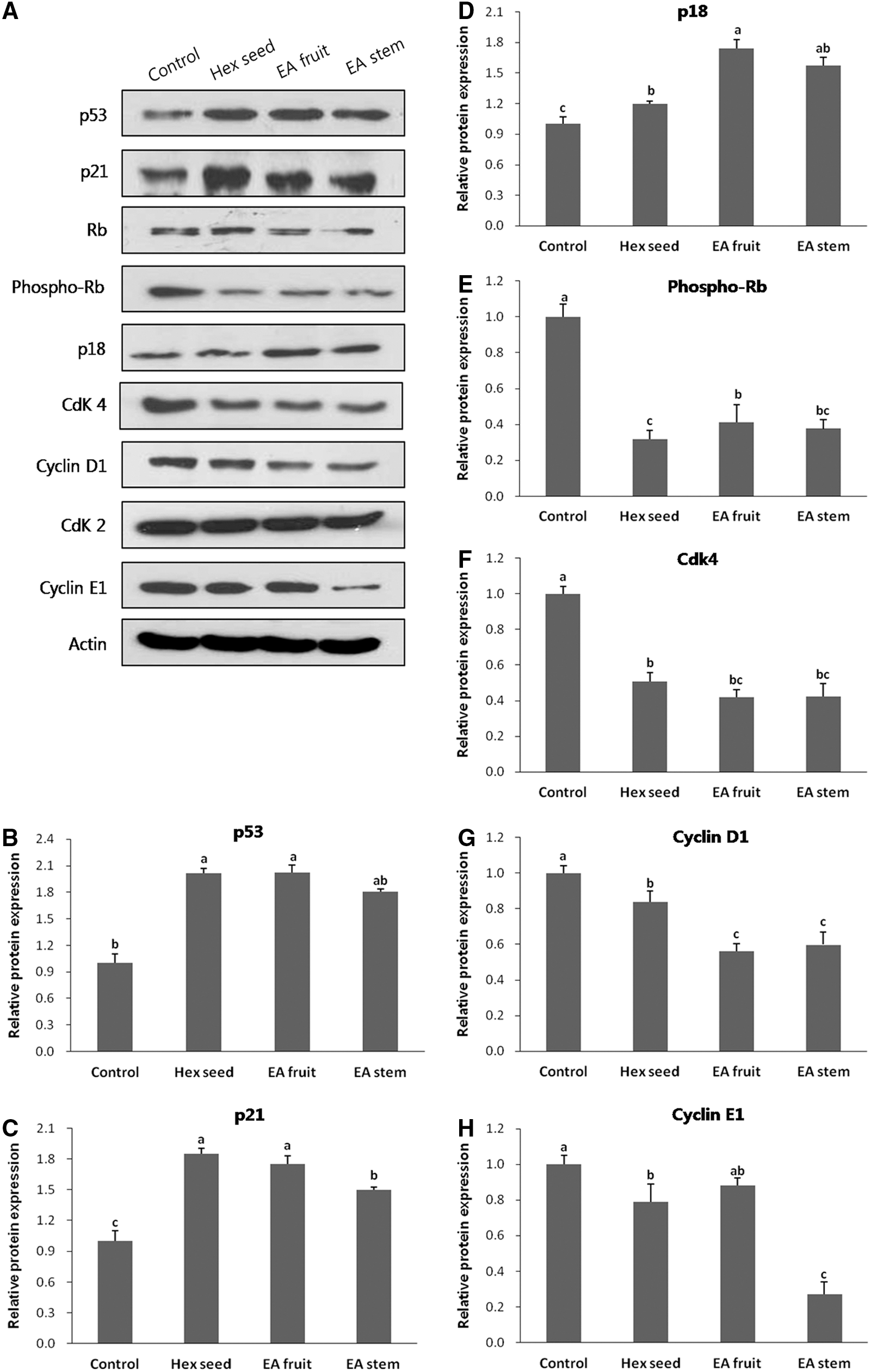
Expression of cell cycle regulatory proteins in HeLa cells treated with extracts of O. humifusa. Cells were treated with 100 μg/mL of O. humifusa extracts and incubated for 24 h. Total cell lysates were extracted from HeLa cells to examine the expression levels of cyclin D1, Cdk4, cyclin E1, Cdk2, phospho-retinoblastoma protein (Rb), Rb, p18, p53, and p21. Equal amounts of total protein from each preparation were separated by sodium dodecyl sulfate–10% polyacrylamide gel electrophoresis (SDS-PAGE), and for subsequent western blot analysis.
OHF inhibit HeLa tumor xenograft growth in nude mice
Based on the inhibitory effects of O. humifusa extracts on cell growth, we examined the effects of OHF on tumor growth by using nude mice subcutaneously injected with HeLa cells. Tumors readily formed in all nude mice, but substantial differences in tumor volume were observed beginning on day 12 of the treatment (Fig. 6A). Additionally, the magnitude of difference between control and OHF-treated groups were dependent on the length of the treatment. However, no dose-dependent effect was observed in the OHF-treated groups. After the 30-day treatment, the weights of tumors from OHF-treated groups (OHF100, OHF200, and OHF500) were significantly lower (P<.05) than those from the control group (Fig. 6B). The average tumor weight in the control group was 0.47 g, whereas the average tumor weight in the OHF-treated groups was 0.27 g. The four groups of mice had a similar mean body weight and feed intake (Fig. 7). Tissue weights of the lungs, liver, kidneys, spleen, and uterus did not significantly differ among the groups (Fig. 7).
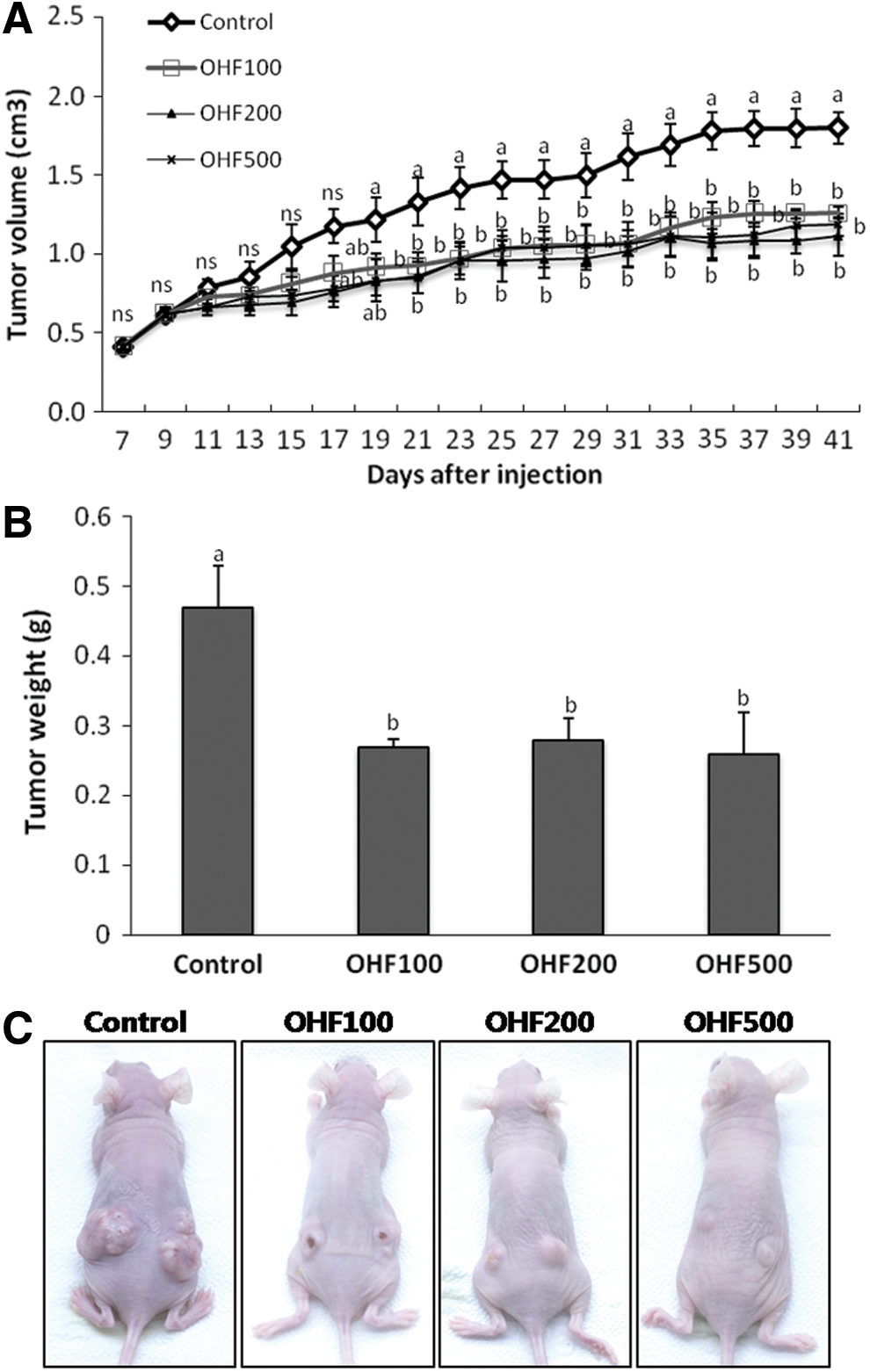
Inhibitory effect of ethyl acetate extracts of O. humifusa fruit (OHF) on the growth of transplantable tumors.
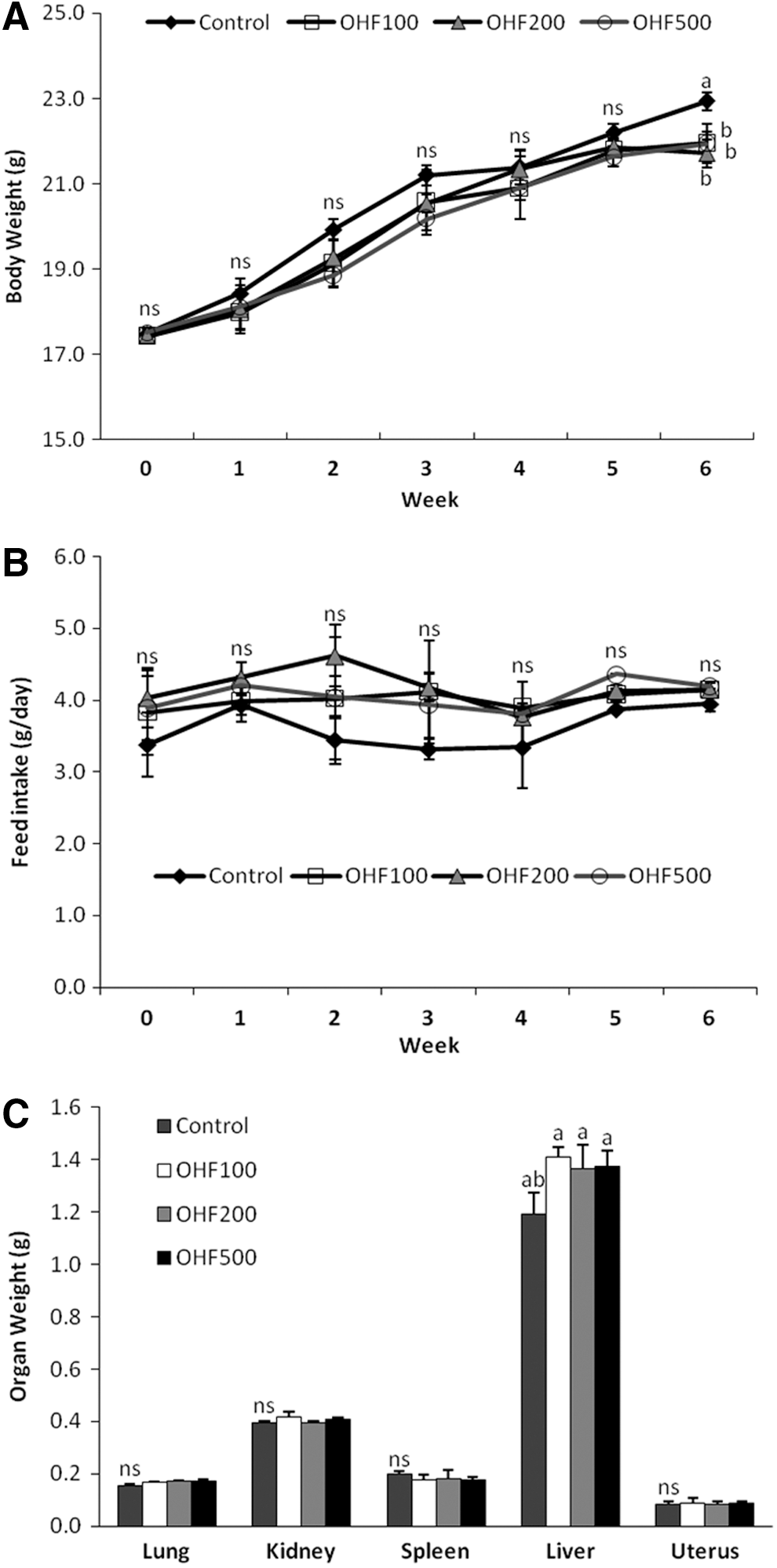
Effect of treatment with OHF on body weight changes, feed intake, and organ weight of nude mice injected subcutaneously with HeLa cells. Body weight
To obtain biochemical evidence for the in vivo results, the expression of key proteins involved in cell cycle arrest was examined. The p53 and p21WAF1/Cip1 mRNA expression were greater (P<.05) in tumors from mice in OHF-treated groups (OHF100, OHF200, and OHF500) compared with those in the control group (Fig. 8). In addition, protein expression of p53 and p21WAF1/Cip1 significantly increased (P<.05) in OHF-treated groups (Fig. 9). Among the OHF groups, OHF500 reduced the expression of phosphorylated Rb and Cdk4 in tumor tissues most effectively (P<.05) (Fig. 9). Furthermore, cyclinD1 expression was significantly decreased (P<.05) in tumors of OHF100 and OHF500 (Fig. 9). The expression levels of Cdk2 and cyclin E did not change significantly.
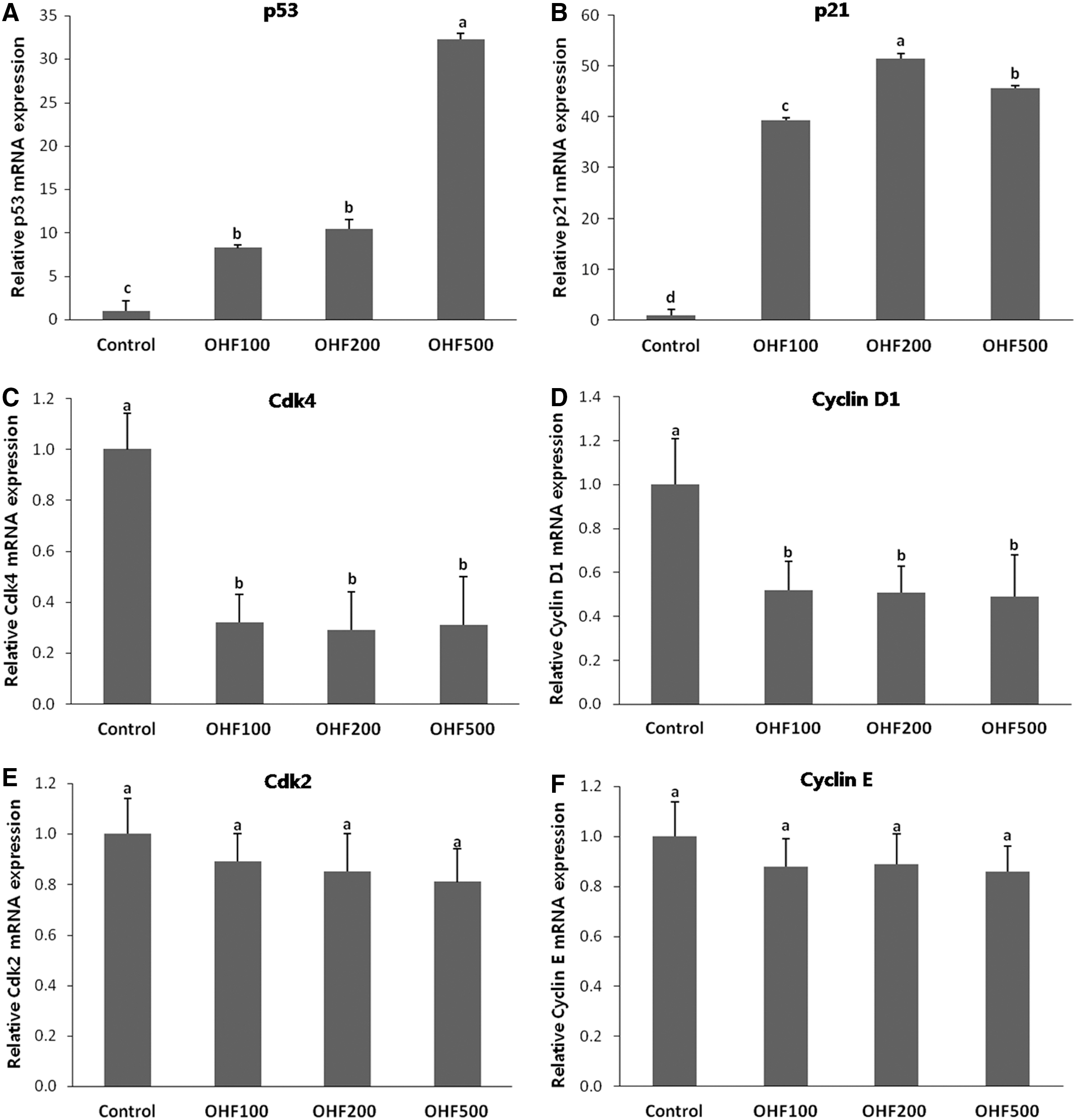
Gene expression of
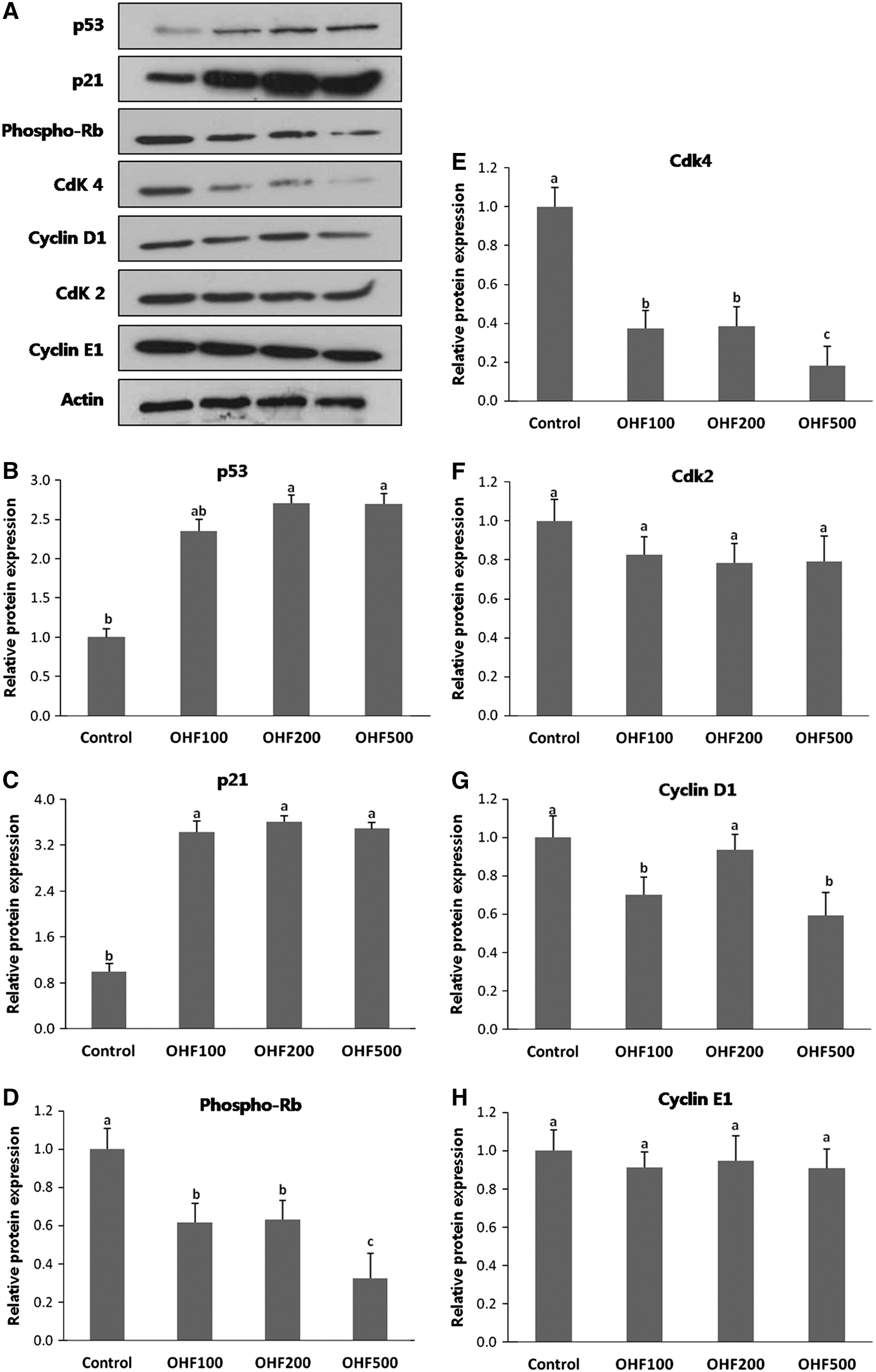
Expression of cell cycle regulatory proteins in tumor tissues of the mice HeLa cells xenografts. Equal amounts of total protein from each preparation were separated by SDS-PAGE, and for subsequent western blot analysis.
Identification and structure determination of the isolates from O. humifusa fruits
Compound 1 was obtained as a yellow powder. The ESI-MS spectrum showed m/z: 287.04 [M-H]− (corresponding to C15H12O6: 288.2), and the presence of 15 carbon atoms was established from the 13 C-NMR spectra. In its 1 H-NMR spectrum, a meta-coupled proton signals at 5.92 (1H, d, J=2.0 Hz, H-8), and 5.88 (1H, d, J=2.0 Hz, H-6) were observed. Thus, the structure of this compound was assigned as dihydrokaempferol (Fig. 10A) based on a comparison of physical and spectroscopic data with reported values. 28 Compound 2 was obtained as a yellow powder. The ESI-MS spectrum showed m/z: 305.99 [M+H]+ (corresponding to C15H12O7: 304.2), and the presence of 15 carbon atoms was established from the 13 C-NMR spectra. The 1 H-NMR data were almost identical to those reported for taxifolin. 29 In particular, the signals for H-2 and H-3 appeared at δ 4.60 and 5.01 with a coupling constant (J value) of 11.2 Hz, which is well in accordance with that for the trans-form. Thus, the structure of this compound was elucidated as trans Taxifolin (Fig. 10B) (dihydroquercetin) based on a comparison of physical and spectroscopic data with reported values.

Chemical structures of dihydrokaempferol
Discussion
Natural products and health foods are increasingly recognized by health professionals and the general population for improving the overall well-being, as well as prevention of diseases. The antioxidant contents of O. humifusa are shown in the present study. The β-carotene level in the stem appears higher than in other parts of O. humifusa. Epidemiological studies have shown that high intakes of carotenoid-rich vegetables and fruits and high blood levels of β-carotene are associated with decreased incidence of some cancers. 30 Enger et al. 31 showed that β-carotene may provide more effective protection against the development of colonic adenomas compared with other dietary carotenoids. The ascorbic acid level in the fruit of O. humifusa (445 mg/100 g) is considerably higher than the average ascorbic acid levels in other common fruits, such as lemon 32 and apple. 33 Vitamin C is a water-soluble antioxidant that scavenges reactive oxygen and nitrogen species. Besides exerting antioxidant influence directly, vitamin C can promote the removal of oxidative DNA damage from the DNA and/or nucleotide pool through the upregulation of repair enzymes, perhaps induced by the vitamin's pro-oxidative properties. 34 Additionally, we show, in this study, that O. humifusa contains higher concentrations of total polyphenols as well as flavonoids when compared with other types of cactus 35 ; this is especially true for the fruits of the plant. The presence of phenolic compounds such as phenolic acids, quinines, flavonoids, and tannins is associated with the antioxidant and anticancer activities of extracts from medicinal plants and herbs. 36
The ethyl acetate extracts of fruit and stem significantly decreased the proliferation of HeLa cells. Similar result has been reported that the ethyl acetate extract of O. humifusa stem repressed the breast cancer cells proliferation. 37 The seed extracts of fruits and vegetables are generally used as an anticancer and antioxidant treatment. 38,39 It has been reported that the hexane extract of Nigella sativa seeds have killed HeLa cells through inducing apoptosis. 40 Also, methanol and hexane extracts of molluginaceae seed showed anticancer activity. 41 In the present study, hexane extracts of seeds have been shown to have an inhibitory effect on HeLa cells, whereas ethyl acetate extracts of seeds caused an increase in the cell proliferation. These data are interpreted to mean that biologically active compounds were separated by fractionation methods. Therefore, additional studies are needed to identify compounds of the seeds required for the opposite effect on cell proliferation.
Recently, many studies have shown that the cactus has antiproliferative activity against a wide range of cancers. 42,43 While several cactus plant extracts possess anticancer effects, the pharmacological effects of O. humifusa have not been systematically examined. In this study, extracts of O. humifusa were shown to inhibit the proliferation of human cervical cancer HeLa cells by arresting the cell cycle during the G1 phase by upregulation of p21WAF1/Cip1 and downregulation of cyclin D1 and Cdk4. The decreased proliferation observed in HeLa cells treated with O. humifusa extracts correlates with the growth arrest that typically occurs when cultured cells are treated with polyphenolic compounds, such as genistein, 44 curcumin, 45 and tea catechins. 46 The results of the flow cytometric analysis showed that the number of cells in the G1 phase increased with increase in the inhibition ratio. Zou et al. 47 reported a similar result, stating that cactus products inhibit cervical cancer cell proliferation and induce G1 arrest. Moreover, several cases of induced cell cycle arrest and cell death after 24 h of treatment have been observed in HeLa cells. 21,48 –50
We also examined cell cycle regulatory factors to identify the mechanisms by which O. humifusa extracts induce G1 cell cycle arrest. Cell cycle dysregulation is clearly associated with tumorigenesis. Generally, this is thought to be due to an increased level or activity of cyclins and their cognate Cdks or a reduction in the level or activity of CKIs. When cyclin D1 is overexpressed at the G1 stage, phosphorylation of Rb is accelerated, promoting cell proliferation. 51 The overexpression of cyclin D1 has been observed in various tumor cells. 52,53 Interestingly, we observed the downregulation of cyclin D1 in HeLa cells treated with extracts of O. humifusa in the present study. Furthermore, the level of p21WAF1/Cip1 and p18INK4 CKI proteins increased after treatment with either seed hexane extracts, or fruit and stem ethyl acetate extracts. Since inhibition of CKI can cause uncontrolled proliferation of tumor cells, a possible anticancer strategy is to induce CKI expression. 54 p18INK4 is tightly regulated during the cell cycle phase in cultured cell lines; it increases during the G1 phase and peaks during the S phase. 55 Deletions at the p18INK4 locus have previously been implicated in the pathogenesis of other tumor types. 56,57 p21WAF1/Cip1, a key molecule in cell cycle regulation, binds to Cdk proteins and inhibits their activity. 58 Given the importance of p21WAF1/Cip1 in regulating the G1-S and G2 checkpoints, this protein has been implicated in the response of many cancers to treatment. 59 In this study, we observed that treatment of HeLa cells with O. humifusa extracts induced an increase in levels of the tumor suppressor protein, p53. The p53 is an important regulator of progression through the G1 phase, as its activation leads to the upregulation of p21WAF1/Cip1. 60,61
We found that the OHF inhibited tumor growth in a HeLa cells xenograft model. A previous study has shown that flavonoids exhibit antitumor activity against HeLa xenografts, and had no effect on body weight. 62 The fruits of O. humifusa are rich in functional constituents, such as vitamin C, vitamin E, and polyphenol. Our study showed that OHF increases the expression of p53 and p21 both in vitro and in vivo. Consistent with our results, Yang et al. 63 reported that flavonoid-enriched extracts inhibit cancer cell proliferation by inducing p21 expression in vitro and in vivo. In the present study, it was revealed that trans Taxifolin and dihydrokaempferol constituted the flavonoids in the OHF. In other investigations on cladodes of Opuntia spp., quercetin, kaempferol, and methyl-3-quercetin were detected. 64,65 Moreover, several flavonoids, including isorhamnetin glucoside, isorhamnetin galactoside, and taxifolin, were isolated from the cladodes of O. humifusa. 66 However, fewer studies have reported about the fruits of O. humifusa. In this study, the presence of trans Taxifolin and dihydrokaempferol in OHF was obtained, which proved to be an efficient antioxidant activity. There were similar results that kaempferol and quercetin were found in fruits of O. ficus-indica and O. streptacantha. 67 In addition, previous studies have reported that kaempferol decreased the viability of the human bladder and colon cancer cells through apoptosis inducing. 68,69 Moreover, taxifolin inhibits the proliferation of HCT116 colorectal cancer cell as well as represses prostate cancer cell growth. 70,71 In response, taxifolin has been reported to support dual-compound treatment in cancer cells in conjunction with andrographolide (diterpenoid lactone).
In conclusion, based on the results of this study, we confirmed our primary hypothesis that extracts of O. humifusa would have a cytotoxic and antiproliferative effect on cancer cells. The present study suggests that hexane extracts of seeds and ethyl acetate extracts of fruits and stems of O. humifusa inhibit cell proliferation by causing G1 phase arrest in human cervical carcinoma cells. Moreover, this study provided mechanistic evidence that the G1 phase arrest induced by ethyl acetate extract from fruit is mediated by disrupting protein expression of cyclin D1, Cdk4, and phospho-Rb, and by the upregulation of p21WAF1/Cip1 and p53 both at the mRNA and protein levels. Our results indicate that this extract may be a promising candidate for treating human cervical carcinoma.
Footnotes
Acknowledgment
This work was carried out with the support of a Korea University Grant, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.