
Abstract
Khz (a fusion mycelium of Ganoderma lucidum and Polyporus umbellatus mycelia) is isolated from ganoderic acid and P. umbellatus and it exerts antiproliferative effects against malignant cells. However, no previous study has reported the inhibitory effects of Khz on the growth of human colon cancer cells. In the present study, we found that Khz suppressed cell division and induced apoptosis in HCT116 cells. Khz cytotoxicity was measured using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Khz reduced cell viability and mitochondrial membrane potential levels and it also induced disruption of the mitochondrial membrane potential and increased calcium concentration and reactive oxygen species generation. Khz increased caspase 3, PARP, caspase 7, and caspase 9 levels, but reduced Bcl-2 protein levels. Flow cytometry showed that the percentage of HCT116 cells in the sub-G1 phase of the cell cycle increased in response to Khz treatment.
Introduction
T
In recent years, Ganoderma mushrooms have been used to treat bronchitis, allergies, hepatitis, immune disorders, and cancer, and to prevent high blood pressure. G. lucidum extracts are used to treat colorectal cancer, prostate cancer, colon cancer, acute myelogenous leukemia, and breast cancer, and the antiproliferative effects of its extracts have been shown in many tumor cell lines. 1 –11 Previously, we reported that Grifola umbellata inhibited cancer cell proliferation and induced apoptosis. 12 Khz is an extract from the mycelia of a G. lucidum (Fig. 1A) and P. umbellatus (Fig. 1B) nuclear fusion. The anticancer effect of this fusion product has been previously reported. In this study, we investigated the mechanism underlying Khz-induced cell death in human colon carcinoma HCT116 cells. Cancer cells abnormally proliferate. Thus, the inhibition of proliferation and induction of apoptosis in cancer cells are effective strategies to prevent the occurrence of cancer. Programmed cell death, also known as apoptosis, occurs in cells in response to normal or pathological factors.
Cell shrinkage, nuclear enrichment, chromatin condensation, and DNA fragmentation are indicators of apoptosis. Apoptosis is triggered by a series of external signals and is regulated by the Bcl-2 family of proteins and the cysteine-aspartic acid protease (caspase) family of proteases. Defects in the mechanism or regulation of apoptosis can result in cancer because of the uncontrolled proliferation of cells. Many anticancer drugs act by inducing apoptosis in various types of cancer cells. Thus, apoptosis can be considered as a major anticancer mechanism, and the induction of apoptosis in cancer cells by treating them with natural products is a potential treatment strategy.
The anticancer effects and underlying mechanism of action of the Khz extract were explored in human colon carcinoma HCT116 cells.
Materials and Methods
Cell lines
Human colon carcinoma HCT116 cells were purchased from ATCC and maintained in a RPMI 1640 medium supplemented with 10% fetal bovine serum, 100 U/mL penicillin G sodium, 100 μg/mL streptomycin sulfate, and 0.25 μg/mL amphotericin B. Unless otherwise indicated, all cells were treated with a 1:2 dilution of the Khz extract.
Khz extraction method
First, 1 kg of powder was dissolved in 8.5 L of clean water and extracted at a temperature of 115°C under pressure for 60 min. The remnant from the first extraction was dissolved in 7.5 L of clean water and extracted at a temperature of 115°C for 60 min under pressure. The first and the second extracts were then mixed and boiled for 5 min at 100°C.
Cell viability assay
To determine cell viability, the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used. HCT116 cells were seeded in 96-well plates at a density of 2000 cells/well. For the MTT assay, 10 μL/100 μL MTT was added to each well to reach a final concentration of 500 μg/mL. The mixture was further incubated for 1 h, and the liquid in the wells was removed thereafter. Fifty microliters of dimethyl sulfoxide was then added to each well, and the absorbance was read with a UV max microplate reader (Molecular Device) at 595 nm.
Flow cytometry detection of mitochondrial membrane potential disruption in HCT116 cells
The mitochondrial membrane potential in HCT116 cells was determined by flow cytometry. Briefly, cells were treated with or without Khz for different times (0, 0.5, 1, and 2 h), harvested and washed twice, resuspended in 500 μL of DiOC6 (4 μM; Biocompare), and incubated at 37°C for 30 min. The changes in the mitochondrial membrane potential were then analyzed by flow cytometry.
Apoptosis assay
The cells were treated with Khz, washed twice in cold phosphate-buffered saline (PBS), and stained with annexin V-fluorescein isothiocyanate (FITC, A13199; Invitrogen) and propidium iodide (PI) according to the manufacturer's instructions. Briefly, annexin V-FITC was added to cells that were resuspended in 100 μL of a 1× binding buffer (10 mM HEPES, 140 mM NaCl, and 2 mM CaCld2; pH 7.4). The cells were then incubated at room temperature for 15 min, and PI was added to the cell suspension before flow cytometric analysis was performed (Becton-Dickinson).
Cell cycle analysis
The percentage of cells in the sub-G1, G0/G1, S, and G2/M phases was determined by flow cytometry. HCT116 cells (1×106/10-cm dish) were incubated with Khz for different times (0, 0.5, 1, and 2 h) and harvested by centrifugation. After harvesting, the cells were washed with PBS and fixed gently (drop by drop) in 70% ethanol (in PBS) on ice overnight. The cells were resuspended in PBS containing 40 μg/mL PI, 0.1 mg/mL RNase (Sigma-Aldrich), and 0.1% Triton X-100. After 30 min at 37°C in the dark, the cells were analyzed using a flow cytometer equipped with an argon laser with excitation at 488 nm.
Detection of reactive oxygen species
The levels of cytoplasmic reactive oxygen species (ROS) were estimated using the oxidation-sensitive fluorescent dye, H2DCF-DA (2,7-dichlorodihydrofluorescein diacetate; Invitrogen). For DCF staining, the cells were loaded with H2DCF-DA (100 nM) for 1 h at 37°C and washed once with PBS. After Khz treatment, the ROS levels were analyzed with a flow cytometer.
Detection of calcium increase
The Ca2+ levels in the HCT116 cells were determined by staining with fluo-4 AM. The cells were pretreated with the Khz extract for different times (0, 0.5, 1, and 2 h), harvested and washed twice, and then resuspended with fluo-4 AM. The cells were then incubated at 37°C for 30 min, and the changes in Ca2+ concentration were analyzed by flow cytometry.
Western blot analysis
The cells were lysed in an extraction buffer (31.25 mM Tris-HCl [pH 6.8], 1% sodium dodecyl sulfate [SDS], 10% glycerol, and 2.5% mercaptoethanol), and the whole cell lysates were subjected to SDS-polyacrylamide gel electrophoresis. The size-fractionated proteins on the gel were transferred onto a nitrocellulose membrane. The membrane was blocked in 5% skim milk in Tris-buffered saline containing 0.05% Tween 20 and incubated with a primary antibody. Antibodies against caspase 3, caspase 7, caspase 9, PARP, and Bcl-2 were purchased from Cell Signaling. After washing, the membrane was incubated with a peroxidase-conjugated secondary antibody. The protein bands of interest were detected using enhanced chemiluminescence reagents (Amersham).
Statistics
Cell viability is expressed as mean±standard deviation. Statistical comparisons were made between the control and treatment groups using Student's t-test. P≤.05 was considered statistically significant.
Results
Effect of Khz on cell survival
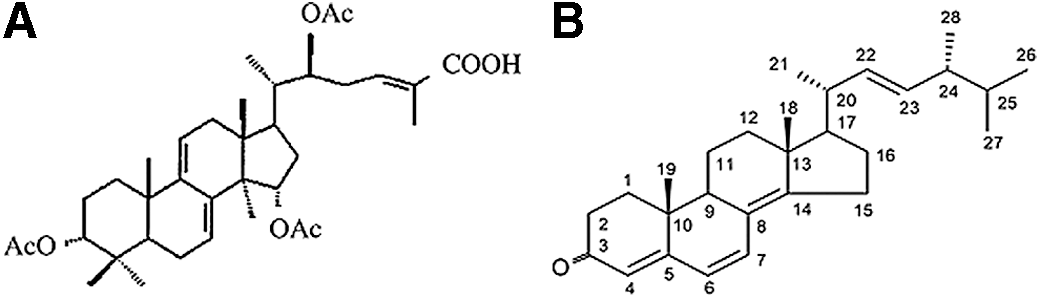
The extracted Khz product is shown in Figure 2. The aim of the present study was to examine whether Khz induced apoptosis in human cancer cells and, if so, to identify the signaling mechanisms involved in the process. We assessed the inhibitory effects of Khz on cell growth in HCT116 cells. The HCT116 cells were treated with a 1:2 dilution of Khz for 0 (control), 0.5, 1, and 2 h. These results indicate that Khz inhibits proliferation of cancer cells (Fig. 2).
HCT116 cells were treated with Khz for 0, 0.5, 1, and 2 h. The cytotoxicity of Khz was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as described in the Materials and Methods. Khz significantly inhibited HCT116 cell growth. Data in figure indicate mean±standard deviation, n≥3. **P<.01. Color images available online at
Effect of Khz on cell cycle progression
The effects of Khz on the growth and survival of HCT116 cells were tested by flow cytometry. In the HCT116 cells treated with Khz for 0.5, 1, and 2 h, the sub-G1 population increased by 4.24%, 8.38%, and 10.77%, respectively (Fig. 3). These results indicate that Khz induces apoptosis in HCT116 cells in a time-dependent manner.

HCT116 cells were exposed to Khz for 0.5, 1, and 2 h. The cells were harvested and the cell cycle analyzed by flow cytometry. The percentage of cells in the sub-G1 phase increased after Khz treatment for 0.5, 1, and 2 h.
Effect of Khz on the levels of caspase-3, PARP, caspase-7, caspase-9, and Bcl-2
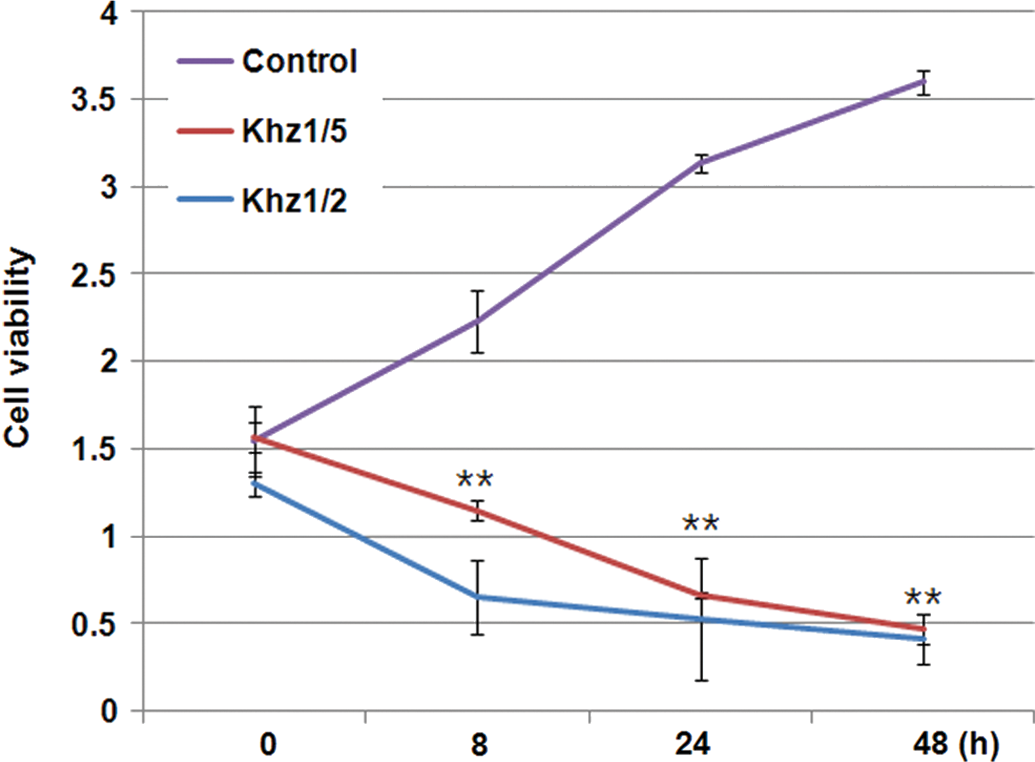
The levels of expression of caspase-3, PARP, caspase-7, caspase-9, and Bcl-2 in HCT116 cells after Khz treatment for 0, 0.5, 1, and 2 h were assessed by western blotting. The results showed that the levels of caspase-3, PARP, caspase-7, and caspase-9 increased in response to Khz treatment, whereas the level of the Bcl-2 protein decreased (Fig. 4). Therefore, Khz suppressed the expression of Bcl-2 and increased the expression of caspase-3, caspase-7, caspase-9, and PARP.
Effects of Khz on the protein levels of caspase 3, PARP, caspase 7, caspase 9, and Bcl-2 in HCT116 cells. Cells were cultured and treated with Khz for up to 2 h. Cell lysates were analyzed by immunoblotting with antibodies against caspase 3, PARP, caspase 7, caspase 9, Bcl-2, and β-actin.
Effect of Khz on the generation of ROS
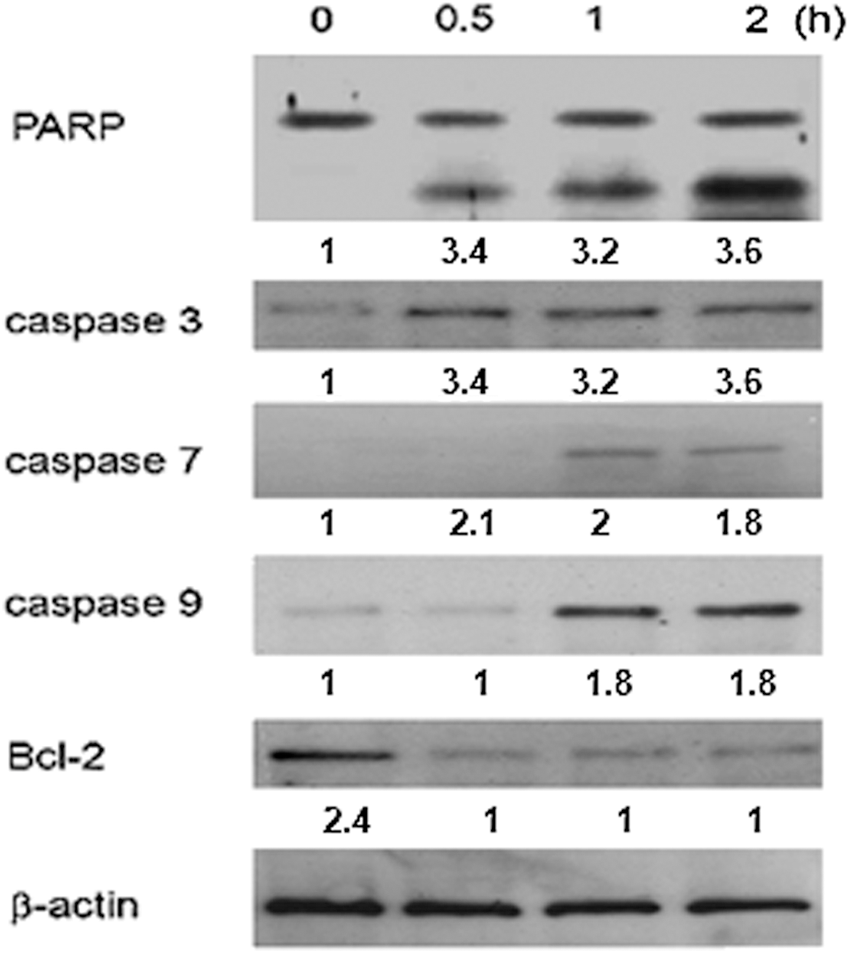
Oxidative stress has been associated with both apoptotic and nonapoptotic cell death, although a pro-oxidative condition is not a prerequisite for apoptosis. 13 –19 Free radicals are known to induce apoptosis. There was a quantifiable increase in the ROS concentration in response to Khz treatment. After 2 h, the ROS levels increased from 9.2% (in the control) to 11.4%, 41.7%, and 58.1% in the Khz-treated cells in a time-dependent manner. Increasing the incubation time resulted in the accumulation of ROS (Fig. 5), indicating that Khz-induced apoptosis is dependent on ROS generation.
Effects of Khz on the production of reactive oxygen species (ROS) in HCT116 cells. HCT116 cells were treated with Khz for 0, 0.5, 1, and 2 h, and the ROS level increased by 9.2%, 11.4%, 41.7%, and 58.1%, respectively. ROS were evaluated as described in the Materials and Methods. Increasing the time of incubation led to an increase in the level of ROS. Color images available online at
Effect of Khz on the increase of intracellular calcium
Recently, sustained elevation of the intracellular Ca2+ concentration ([Ca2+]i) has been associated with the induction of apoptosis. 20 Therefore, we investigated whether the effects of Khz were mediated by calcium-induced apoptosis. The concentration of calcium increased from 6.3% (in the control group) to 9.3%, 21.1%, and 51.1% in the Khz-treated cells in a time-dependent manner (Fig. 6), indicating that the effect of Khz on apoptosis is mediated by changes in the intracellular Ca2+ concentration. These results suggest that Khz induces apoptosis in HCT116 cells by increasing the Ca2+ concentration in the cells.
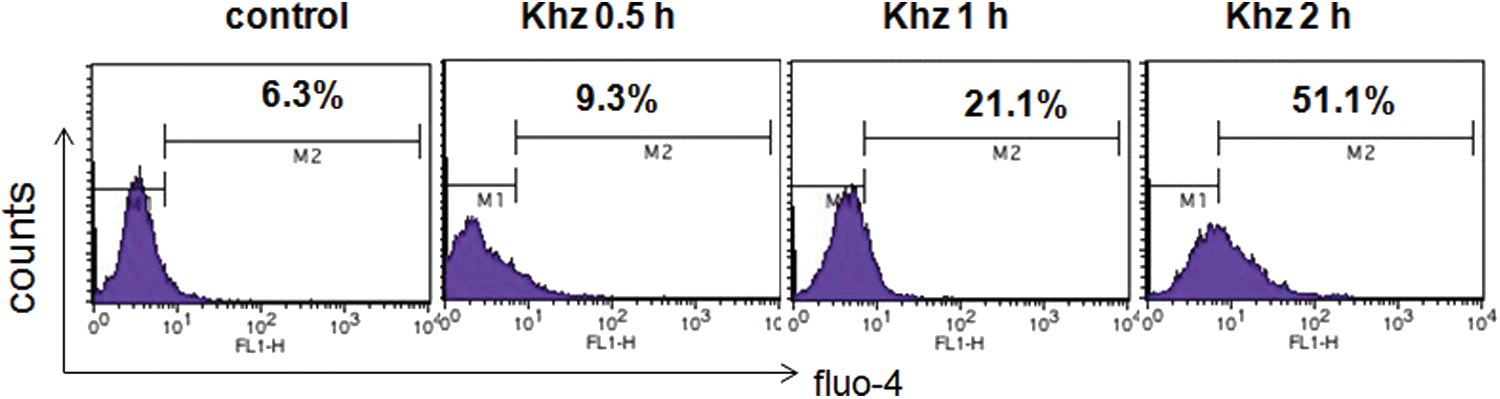
Effects of Khz on the release of Ca2+ in HCT116 cells. HCT116 cells were treated with Khz for the indicated times (0, 0.5, 1, and 2 h). The Ca2+ concentration was evaluated as described in the Materials and Methods. The Ca2+ concentration increased by 6.3%, 9.3%, 21.1%, and 51.1%, respectively. The Ca2+ concentration was different between the Khz-treated group and the control group. Increasing the time of incubation led to an increase in the concentration of Ca2+. Color images available online at
Effect of Khz on mitochondrial membrane potential disruption
Mitochondrial membrane potential is a marker for cell death. We determined the mitochondrial membrane potential in the Khz-treated HCT116 cells. The fraction of cells with a disrupted mitochondrial membrane potential increased from 13.8% (in the control group) to 19.8%, 34.0%, and 53.7% after Khz treatment (Fig. 7). These results suggest that Khz-induced apoptosis involves changes in the mitochondrial membrane potential. Thus, Khz induces apoptosis in HCT116 cells through a mitochondrial-dependent pathway.

Effects of Khz on the MMP in HCT116 cells. Treatment of HCT116 cells with Khz for 0, 0.5, 1, and 2 h increased the fraction of cells with a disrupted MMP by 13.8%, 19.8%, 34.0%, and 53.7%, respectively. The MMP was evaluated as described in the Materials and Methods. Increasing the time of incubation led to a decrease in the MMP.
Discussion
In a previous study, it was suggested that tanshinone II-A induced apoptosis in human lung cancer A549 cells through the induction of ROS and reduction of the mitochondrial membrane potential. 21 We have also shown that Khz induces apoptosis by increasing intracellular calcium levels and activating the JNK and NADPH oxidase-dependent generation of ROS. 12 We suggested that Khz induces apoptosis in A549 human lung cancer cells by generating ROS and decreasing the mitochondrial membrane potential. 22 We also demonstrated that Khz-cp (crude polysaccharide extract obtained from the fusion of G. lucidum and P. umbellatus mycelia) induces apoptosis by increasing intracellular calcium levels and activating P38 and NADPH oxidase-dependent generation of ROS in SNU-1 cells. 23
We fused G. lucidum and P. umbellatus mycelia to obtain Khz, which inhibited cell proliferation in human colon carcinoma HCT116 cells (Fig. 2). In addition, we investigated the cell cycle arrest by Khz treatment (Fig. 3) and elucidated the mechanism by which Khz induces apoptosis in cancer cells. The levels of cleaved, that is, activated caspases 3, 7, 9, and PARP were significantly increased in Khz-treated cells (Fig. 4). Oxidative stress is associated with apoptotic as well as nonapoptotic cell death, although pro-oxidative conditions are not a prerequisite for apoptosis (Fig. 5). It is widely accepted that calcium signaling plays an important role in apoptosis. We observed an increase in intracellular [Ca2+]i in response to Khz treatment (Fig. 6). Our results also showed that Khz increases the decreases in mitochondrial membrane potential (Fig. 7).
Taken together, these results suggest that Khz induces apoptosis by activating caspases and that apoptosis induction by Khz requires ROS generation and calcium release. The proapoptotic and cytotoxic effects of Khz shown in the present study suggest its potential as a chemotherapeutic agent for human colon cancer. Further investigation of the effects of Khz, including in vivo studies, is necessary to determine its potential for clinical use.
Footnotes
Author Disclosure Statement
No competing financial interests exist.