
Abstract
Lutein is a carotenoid pigment present in fruits and vegetables that has anti-inflammatory and antitumor properties. In this study, we examined the effect of lutein on proliferation and survival-associated genes in prostate cancer (PC-3) cells. We found that in vitro culture of PC-3 cells with lutein induced mild decrease in proliferation that improved in combination treatment with peroxisome proliferator-activated receptor gamma (PPARγ) agonists and other chemotherapeutic agents. Flow cytometry analyses showed that lutein improved drug-induced cell cycle arrest and apoptosis in prostate cancer. Gene array and quantitative reverse transcription-polymerase chain reaction analyses showed that lutein altered the expression of growth and apoptosis-associated biomarker genes in PC-3 cells. These findings highlight that lutein modulates the expression of growth and survival-associated genes in prostate cancer cells.
Introduction
D
Current treatment for prostate cancer includes surgery, radiation, hormone and chemotherapy, which yield poor prognosis and toxic side effects. Doxorubicin and Taxol are two chemotherapeutic agents commonly used in treating prostate cancer. 9,10 Temozolomide (TMZ) is an alkylating agent that induces O6-methylguanine formation leading to apoptosis and has been commonly used to treat patients with malignant glioma. 11 However, the influence of lutein on the therapeutic efficacy of TMZ in prostate cancer is not known. Peroxisome proliferator-activated receptor gamma (PPARγ) is a nuclear receptor transcription factor that regulates cell growth, differentiation, and homeostasis. 12 PPARγ agonists modulate lipid metabolism and glucose homeostasis and have been used as prescription drugs for type 2 diabetes. We and others have shown earlier that PPARγ agonists induce growth arrest and apoptosis in different types of tumors. 13 –15 Other studies have also demonstrated that PPARγ agonists induce growth arrest by modulating the expression and activation of PPARγ in prostate cancer. 15 –17 However, the influence of lutein on the inhibition of prostate cancer by these therapeutic agents is not known.
In this study, we examined the antitumor effect of lutein in combination with chemotherapeutic agents and PPARγ agonists in prostate cancer. We found that lutein induces antitumor effect by modulating growth and survival-associated genes in PC-3 cells, suggesting its significance in the treatment of prostate cancer.
Materials and Methods
Reagents
RPMI 1640 medium, fetal bovine serum (FBS), penicillin, streptomycin, propidium iodide, and RNase A were purchased from Life Technologies (Grand Island, NY, USA). Lutein (≥90% pure isolated from marigold, Fig. 1) was purchased from Sigma Chemicals (St. Louis, MO, USA) and diluted in phosphate buffered saline (PBS) or RPMI medium. Doxorubicin, Paclitaxel (Taxol), Temozolomide (TMZ), Pioglitazone, 15-Deoxy-Δ 12,14 -prostaglandin J2 (15d-PGJ2), and DMSO were obtained from Sigma (St. Louis, MO, USA). Ciglitazone was purchased from EMD Biosciences (Billerica, MA, USA). The human prostate cancer biomarker array kit was obtained from SuperArray/SA Bioscience (Valencia, CA, USA). The reagents and primer sets specific to TGFβ-2, IGF1R, EGFR, CDK-9, CDK-7, BRCA1, BCL-2, and GAPDH for quantitative reverse transcription-polymerase chain reaction (qRT-PCR) were obtained from Applied Biosystems (Foster City, CA, USA). WST-1 reagent (4-[3-(4-iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio]-1,3-benzene disulfonate) and the Annexin-V-FLUOS staining kit were purchased from Roche (Indianapolis, IN, USA).
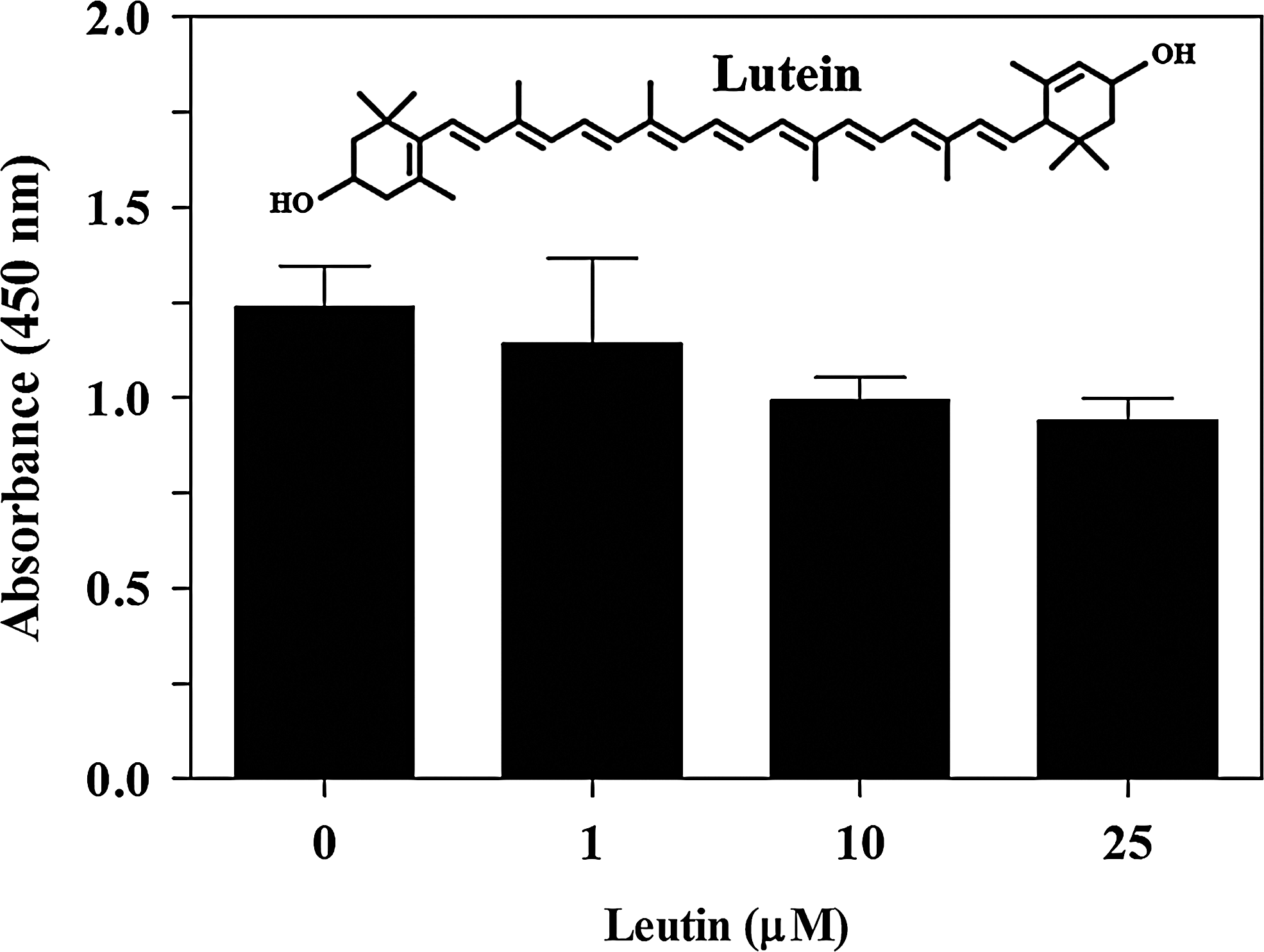
Effect of lutein on the proliferation of prostate cancer cells. PC-3 cells were cultured with increasing doses of lutein. After 48 h WST-1 reagent was added and the absorbance at 450 nm was detected as a measure of proliferation. The values are mean±SD of triplicates and the figure is a representative of three independent experiments. The chemical structure of lutein is shown as an insert.
Cell culture
Hormone-refractory human prostate cancer (PC-3) cell line was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). PC-3 cells were cultured in RPMI-1640 medium supplemented with 10% FBS, 100 U/mL of penicillin, and 100 μg/mL of streptomycin in 5% CO2 incubator at 37°C. Cells were passaged in T-75 tissue culture flasks as and when they were ∼85% confluent. Cells from passage 2 and 3 were used in the experiments.
Proliferation assay
The effect of lutein on prostate cancer cell proliferation was measured by WST-1 assay. PC-3 cells were cultured in RPMI medium (1×104/200 μL/well) in 96-well tissue culture plates in 5% CO2 incubator at 37°C. Increasing doses of lutein alone or in combination with chemotherapeutic agents and PPARγ agonists were added at the initiation of culture. After 48 h, 10 μL of WST-1 reagent was added to each well and the absorbance measured at 450 nm using a titer-plate reader (Alpha Diagnostics, San Antonio, TX, USA).
Cell cycle analysis
The effect of lutein on cell cycle progression of prostate cancer cells was determined by propidium iodide staining and flow cytometry. PC-3 cells were cultured in RPMI medium (5×105/mL) in the absence or presence of 10 μM lutein alone and in combination with 100 μM of chemotherapeutic agents or 25 μM of PPARγ agonists in 12-well plates. Cells treated with vehicle or 10 μM of lutein were used as controls. Cells were harvested after 24 h of culture and fixed in 70% ethanol overnight. Cells were then incubated in PBS containing 100 μg/mL of propidium iodide, 0.6% NP-40, and 20 μg/mL RNase (Sigma Chemicals) at 4°C for 1 h. The percentage of cells at different cell cycle stages (G0/G1, G2/M, S, and subG0) were determined based on the DNA content by flow cytometry using the FACS Calibur Flow Cytometer (BD Biosciences, San Jose, CA). The data were analyzed using the ModFit LT2.0 software (Verity Software House, Topsham, ME, USA).
Apoptosis assay
The effect of lutein on apoptosis of prostate cancer cells was determined by the Annexin V-FITC staining and flow cytometry. PC-3 cells were cultured in RPMI medium (5×105/mL) in the absence or presence of 10 μM lutein alone and in combination with 100 μM chemotherapeutic agents or 25 μM PPARγ agonists in 12-well plates. Cells treated with vehicle or 10 μM lutein were used as controls. Cells were harvested after 24 h of culture and washed three times in PBS containing 0.1% bovine serum albumin. Cells were then stained with Annexin V-FITC (Roche) in a binding buffer (0.1 M Hepes/NaOH, pH 7.4, 1.4 M NaCl, 0.2 mM) containing 100 μg/mL propidium iodide according to the manufacturer's instruction (Roche). The cells were incubated at room temperature for 30 min in the dark, acquired using the FACS Calibur Flow Cytometer (BD Biosciences) and analyzed using the FlowJo 8.2.6 software (Tree Star, Inc., Ashland, OR, USA).
DNA microarray
The effect of lutein on gene expression profile of PC-3 cells was determined by SuperArray containing 263 biomarker genes associated with the diagnosis and prognosis of prostate cancer (SuperArray/SA Bioscience). In brief, PC-3 cells were cultured in RPMI medium in the absence or presence of 10 μM lutein. The cells were harvested after 18 h and total RNA was extracted using Tri-Reagent according to the manufacturer's instruction (Sigma). Equal quantities of RNA samples were reverse transcribed into cDNA and then converted to cRNA using the SuperArray TrueLabeling-AMP™ 2.0 kit (SuperArray Bioscience Corp., Frederick, MD, USA). The cRNA samples were labeled with Biotin (Roche) and hybridized with SuperArray for 24 h. The arrays were then developed using a chemiluminescence detection system and visualized by exposing to X-ray films. The images were uploaded to GEArray® Expression Analysis Suite 2.0 computer software that reports gene expression profile normalized to GAPDH as internal control. The microarray experiments were conducted in triplicates and a mean fold change in gene expression between vehicle and lutein-treated PC-3 cells was calculated after normalizing to GAPDH. Heat map was constructed using the Excel software (Mirosoft Corporation, Redmond, WA, USA). Box plot and scatter plot were generated using the GraphPad Prizm 5.0 software (GraphPad, La Jolla, CA, USA).
Quantitative RT-PCR
The effect of lutein on the expression of selected genes in PC-3 cells was quantified using qRT-PCR technique. In brief, PC-3 cells were cultured in RPMI medium in the absence or presence of 10 μM lutein. The cells were harvested after 18 h and total RNA was extracted using Tri-Reagent. Equal quantities of RNA samples were reverse transcribed into cDNA using the TaqMan reverse transcription kit (Applied Biosystems). qRT-PCR was performed using primer sets and probes (Taqman™) for eight selected genes (EGFR, IGF1R, BRCA1, CDK-5, KLK14, PCA3, GSTP1, RASSF1) and GAPDH obtained from Applied Biosystems in 96-well format with iCycler MYIQ Real-Time PCR detection system (Bio-Rad, Hercules, CA, USA). The data were analyzed using the MYIQ Real-Time PCR quantification (delta-delta-Ct) study software and the gene expression levels were normalized to GAPDH and presented as relative fold change (RQ) compared with control. The qRT-PCR experiments were conducted in triplicates and the fold change in gene expression between untreated and lutein-treated PC-3 cells was also calculated.
Statistical analysis
The experiments were repeated three or more times and the values are expressed as mean±SD/SEM. The differences between groups were analyzed by one way ANOVA using the GraphPad Prism 5.0 software (GraphPad) and the values with *P<.05, **P<.01, and ***P<.001 were considered significant.
Results
Lutein induces antiproliferative response in prostate cancer
To determine the antitumor effect of lutein in prostate cancer, we first examined its effect on the proliferation of PC-3 cells in culture. As shown in Figure 1, PC-3 cells cultured in RPMI medium with 10% FBS displayed normal growth and expansion. However, PC-3 cells cultured in the presence of lutein showed mild decrease in proliferation, reaching ∼15% inhibition at 10 μM. These findings suggest that lutein exerts mild antiproliferative effect on prostate cancer.
To further explore the beneficial effect of lutein, we examined its influence on Doxorubicin and Taxol, two clinically available chemotherapeutic agents for prostate cancer. As shown in Figure 2A, in vitro treatment of PC-3 cells with Doxorubicin resulted in a dose-dependent decrease in proliferation index, reaching 85% inhibition (P<.001) at 100 μM. At 10 and 25 μM doses, Doxorubicin showed a 40% and 47% decrease in proliferation index that further decreased to 65% and 75%, respectively with the addition of 10 μM lutein (P<.05). Similarly, treatment with Taxol induced a dose-dependent decrease in proliferation index, reaching 39% inhibition (P<.05) at 100 μM dose and that increased to 52% in combination with 10 μM lutein in culture (Fig. 2A). On the other hand, treatment with Temozolomide, a drug of choice for glioma, showed no significant antiproliferative effect on PC-3 cells. Interestingly, a combination treatment with 10 μM lutein resulted in a significant dose-dependent decrease in PC-3 cell proliferation, reaching 51% and 78% inhibition (P<.001) at 50 and 100 μM Temozolomide, respectively in culture (Fig. 2A). These results suggest that lutein potentiates the antiproliferative effect of Temozolomide and Doxorubicin in prostate cancer.
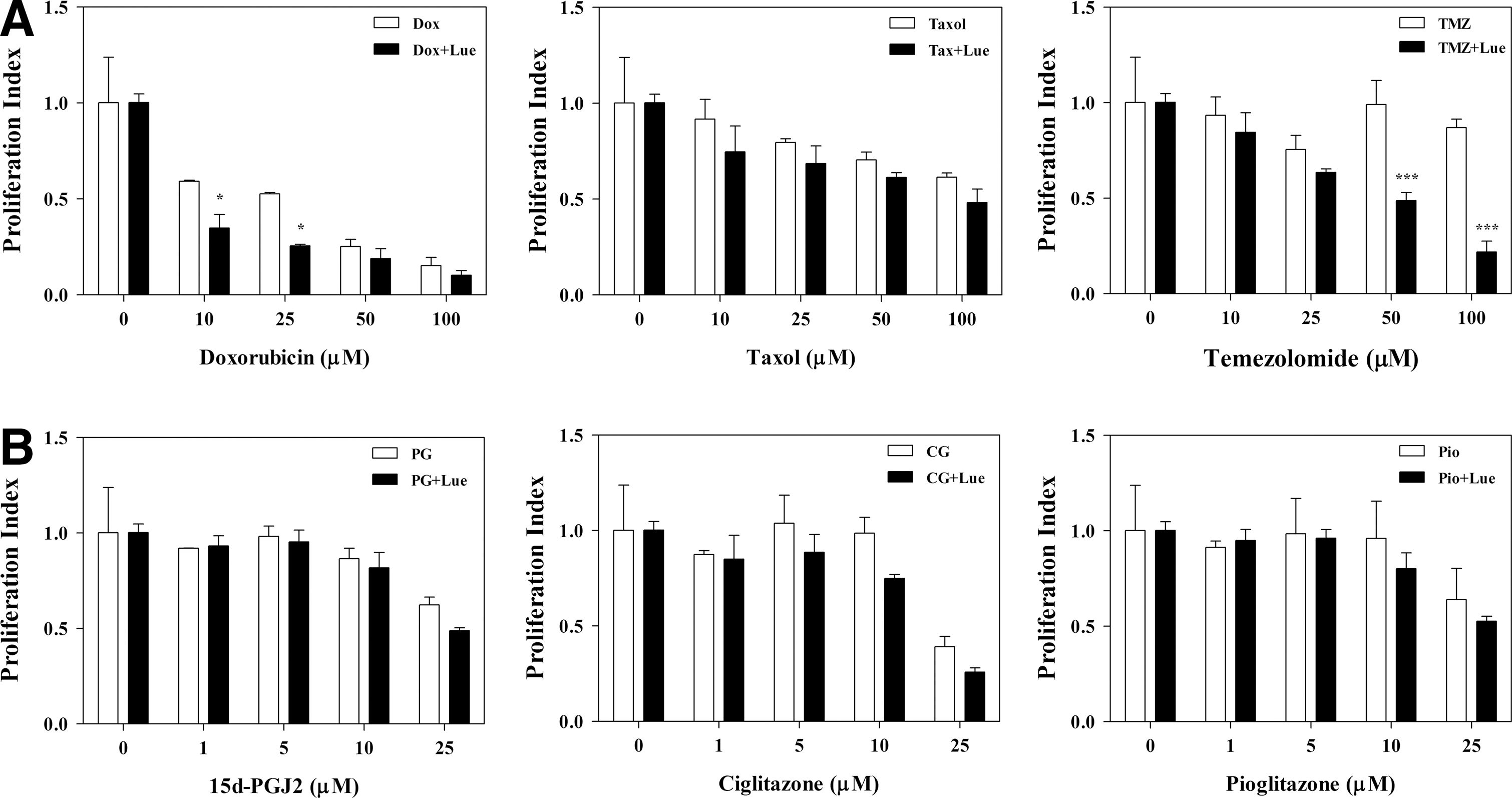
The potentiating effect of lutein on chemotherapeutic agents in prostate cancer cells. PC-3 cells were cultured with increasing doses of Doxorubicin, Paclitaxel, and Temozolomide
We have shown earlier that PPARγ agonists inhibit the growth and survival of brain tumor stem cells, whereas other studies demonstrated inhibition of prostate cancer cells. 13,14,17 In this study, we examined the potentiating effect of lutein on PPARγ agonists in PC-3 cells. We found that in vitro treatment of PC-3 cells with 15d-PGJ2, a natural ligand for PPARγ, decreased proliferation reaching 38% inhibition (P<.05) at 25 μM dose that further increased to 51% (P>.05) in combination with 10 μM lutein (Fig. 2B). Similarly, PC-3 cells treated with Ciglitazone, a synthetic agonist for PPARγ, showed a decrease in proliferation reaching 61% inhibition (P<.01) at the 25 μM dose that increased to 74% (P>.05) in combination with 10 μM lutein (Fig. 2B). Interestingly, the 10 μM dose of Ciglitazone failed to inhibit PC-3 cell proliferation, but the addition of 10 μM lutein led to a 25% inhibition in proliferation index. Moreover, in vitro treatment of PC-3 cells with Pioglitazone, a clinically available synthetic agonist for PPARγ, induced 35% decrease in proliferation (P<.05) at 25 μM dose that increased to 47% (P>.05) in combination with 10 μM lutein (Fig. 2B). These results suggest that lutein potentiates the antitumor effects of PPARγ agonists in prostate cancer.
Lutein modulates cell cycle progression in prostate cancer
To determine the mechanism by which lutein inhibits prostate cancer, we studied its influence on cell cycle progression. As shown in Figure 3, PC-3 cells cultured with lutein displayed significant changes in the distribution of cell cycle phases in the absence or presence of chemotherapeutic agents. When compared to control, the PC-3 cells cultured with 10 μM lutein showed a marked decrease in G0/G1 and G2/M phase in association with an increase in S phase without any change in SubG0 population compared to control (Fig. 3A). On the other hand, PC-3 cells cultured with 100 μM Doxorubicin showed 40% SubG0 cells that increased to 56% in the presence of lutein. Combination treatment with lutein also induced a marked decrease in G0/G1 and G2/M phase without altering S phase compared to Doxorubicin alone (Fig. 3A). Moreover, PC-3 cells cultured with Taxol showed 40% SubG0 cells that increased to 54% in combination with lutein. In addition, there was a marked decrease in S phase without altering G0/G1 and G2/M phase compared to Taxol alone (Fig. 3A). On the other hand, PC-3 cells cultured with Temozolomide showed 0% SubG0 cells that increased to 24% in combination with lutein. In addition, there was a marked decrease in G0/G1 without altering S and G2/M phase compared to Temozolomide alone (Fig. 3A).
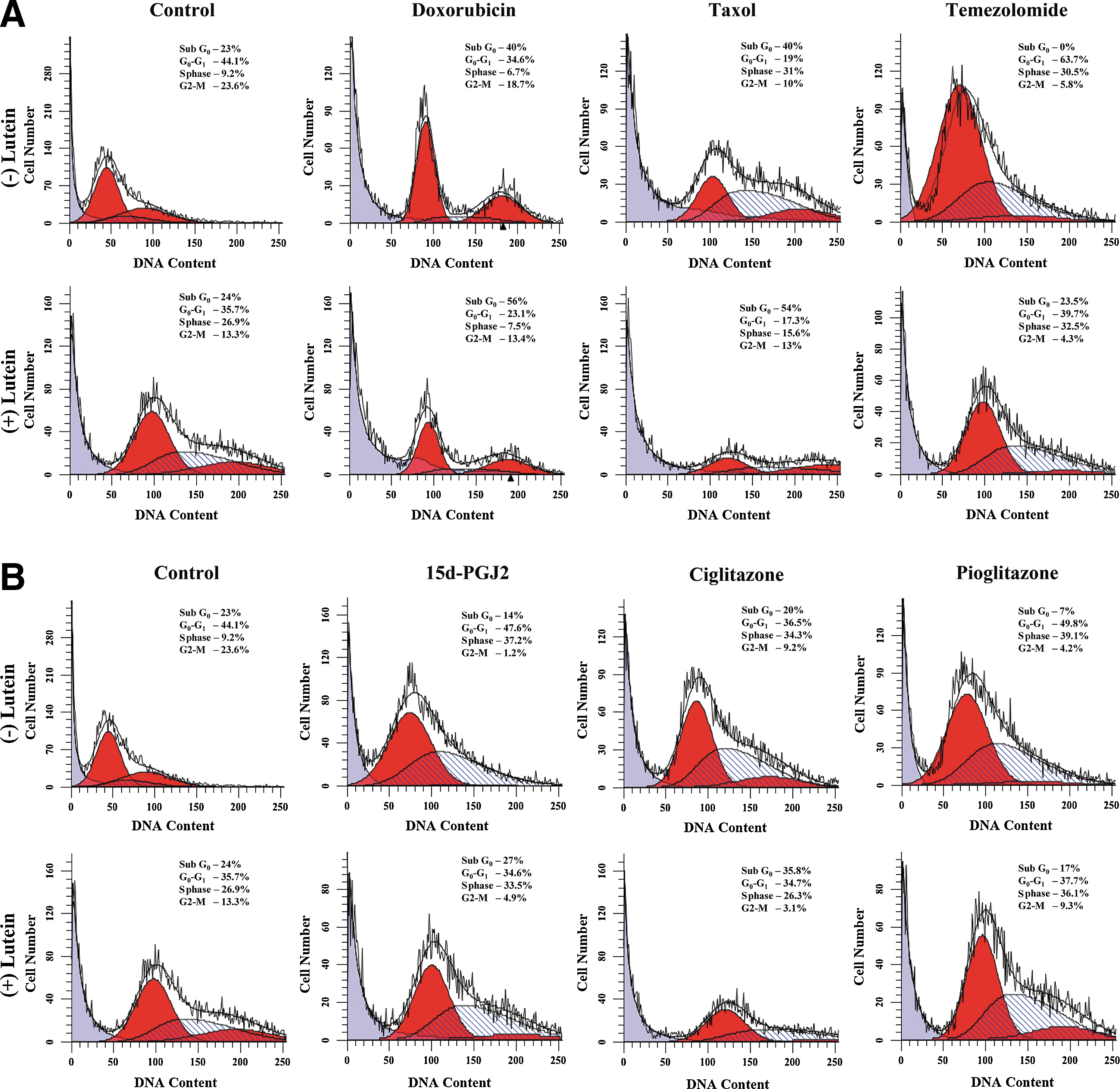
Effect of lutein on chemotherapeutic agent induced cell cycle arrest in prostate cancer cells. PC-3 cells were cultured with different chemotherapeutic agents
Further analyses showed that PC-3 cells cultured in the presence of 25 μM PPARγ agonists displayed significant decrease in G2/M and SubG0 phases with an increase in S phase and without altering G0/G1 phase, compared to control (Fig. 3B). Interestingly, the addition of 10 μM lutein increased SubG0 population in all PPARγ agonist-treated cells, while differentially regulating other cell cycle phases compared to PPARγ agonists alone (Fig. 3B). These results suggest that lutein alone or in combination with chemotherapeutic agents and PPARγ agonists induce marked changes in cell cycle progression in prostate cancer.
Lutein promotes cell death in prostate cancer
To understand the influence of lutein on prostate cancer survival, we examined cell death by apoptosis. As shown in Figure 4, PC-3 cells cultured with 10 μM of lutein displayed 10% apoptosis (P<.01) compared to 2.6% in the control. Interestingly, treatment with Doxorubicin, Taxol, and Temozolomide induced 13%, 3.3%, and 10% Annexin-positive cells that increased to 17%, 8% (P<.001), and 13% in combination with lutein, respectively (Fig. 4A). In addition, PC-3 cells cultured with 10 μM of lutein displayed 15% dead cells (apoptosis + necrosis) compared to 9% in the control. Moreover, treatment with Doxorubicin, Taxol, and Temozolomide showed 44%, 18%, and 16% dead cells that increased to 54%, 28% (P<.001), and 22% (P<.05) in combination with lutein, respectively (Fig. 4A). These results suggest that lutein promotes cell death by apoptosis and necrosis in prostate cancer.
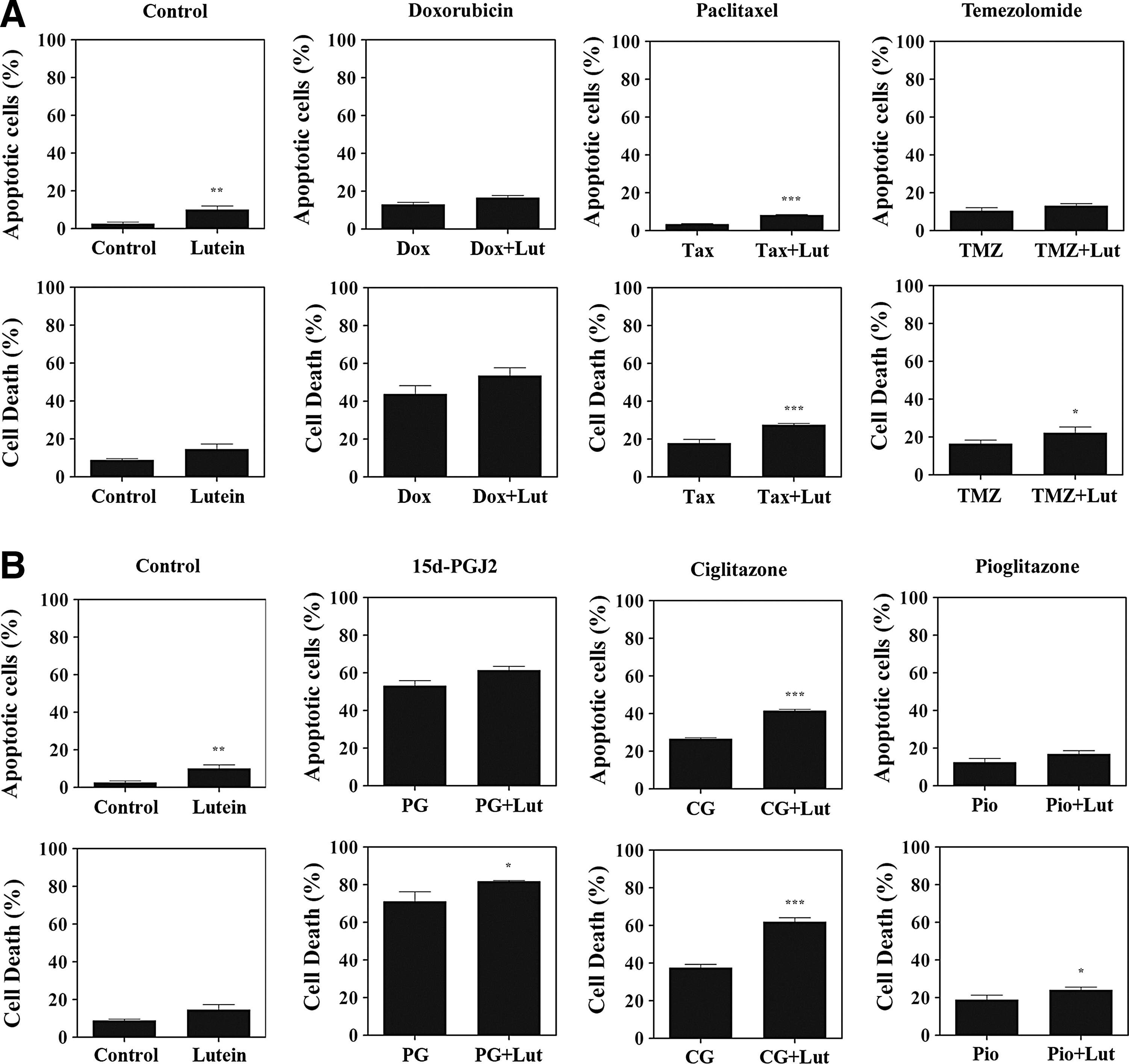
Effect of lutein on chemotherapy-induced apoptosis in prostate cancer cells. PC-3 cells were cultured with different chemotherapeutic agents
We have also studied the influence of lutein on apoptosis in combination with PPARγ agonists in prostate cancer. As shown in Figure 4B, we found that the PC-3 cells cultured with 25 μM Ciglitazone, 15d-PGJ2, and Pioglitazone displayed 27%, 53%, and 12% apoptosis that increased to 41% (P<.001), 61%, and 17% in combination with lutein (Fig. 4B). Further analyses showed that PC-3 cells treated with 25 μM Ciglitazone, 15d-PGJ2, and Pioglitazone displayed 38%, 71%, and 19% dead cells that increased to 62% (P<.001), 82% (P<.05), and 24% (P<.05) in combination with lutein (Fig. 4B). These results indicate that lutein augments PPARγ agonist-induced cell death in prostate cancer.
Lutein inhibits the expression of growth and survival-associated genes in prostate cancer
To understand the molecular mechanism by which lutein attenuates prostate cancer, we studied the expression of biomarker genes in PC-3 cells using SuperArray technique. From this analysis we found that in vitro treatment of PC-3 cells with 10 μM lutein resulted in significant alterations in the expression of several biomarker genes compared to control (Fig. 5). Among the 288 spots examined in the SuperArray, 6 spots including 3 GAPDH remained unchanged, whereas 3 other spots were undetected before and after treatment with lutein (Figs. 5 –7). After normalizing the remaining 279 data points to GAPDH, we found an increase in the expression of 96 genes and a decrease in the expression of 183 genes in lutein-treated PC-3 cells compared to control. Among the upregulated genes, 5 showed >2-fold increase, 15 showed 1.51- to 2-fold increase, 37 showed 1.21- to 1.5-fold increase and 38 genes showed 1.01- to 1.2-fold increase after treatment with lutein (Figs. 6 and 7). Moreover, among the downregulated genes, 66 showed 0.99 to 0.8, 81 showed 0.79 to 0.5, 29 showed 0.49 to 0.3, 6 showed <0.3-fold decrease and one gene became undetected following treatment with lutein (Figs. 6 and 7).
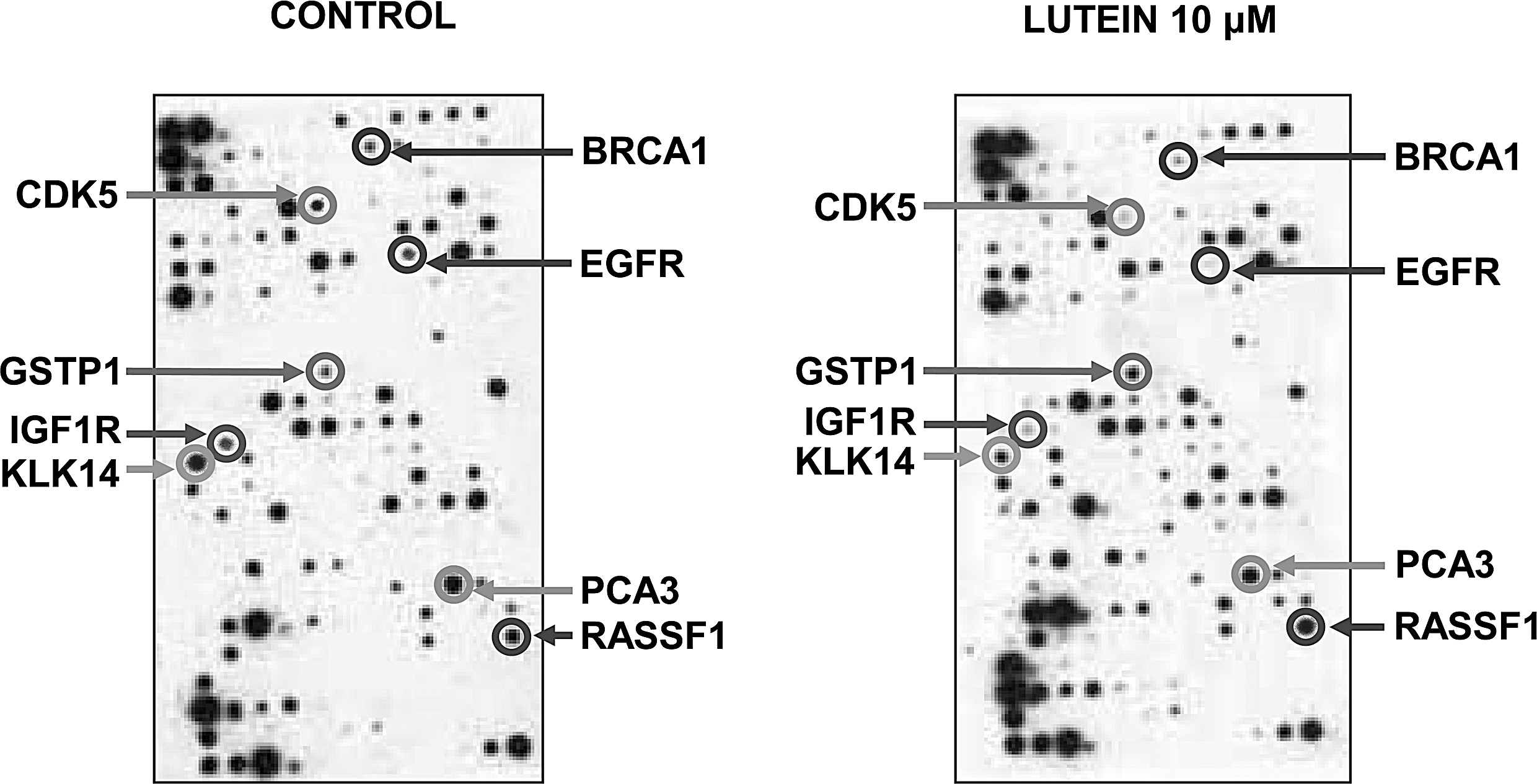
Effect of lutein on prostate cancer biomarker genes. PC-3 cells were cultured in the absence or presence of 10 μM lutein for 18 h and gene expression was analyzed using the SuperArray with 288 spots containing 263 prostate cancer biomarkers and other housekeeping genes. The array was developed using a chemiluminescence detection system and visualized by exposing to X-ray film. Arrows indicate selected biomarker genes with significant changes. The figure is a representative of three independent experiments.
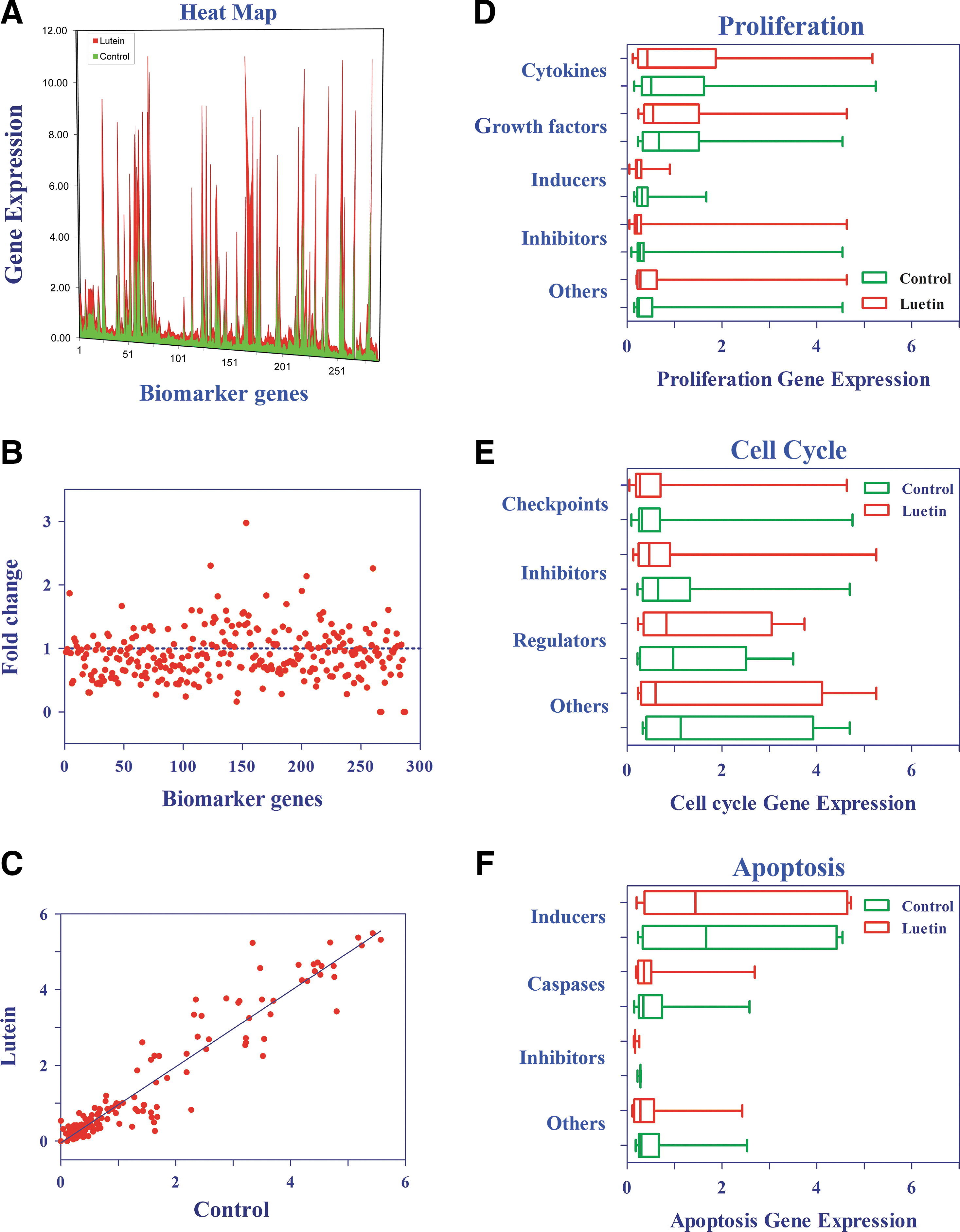
Analysis of lutein regulated prostate cancer biomarker genes. Heat map was constructed using Excel
Effect of lutein on growth and survival-associated genes in prostate cancer cells. Histograms showing selected genes altered by lutein in prostate cancer, as determined by SuperArray
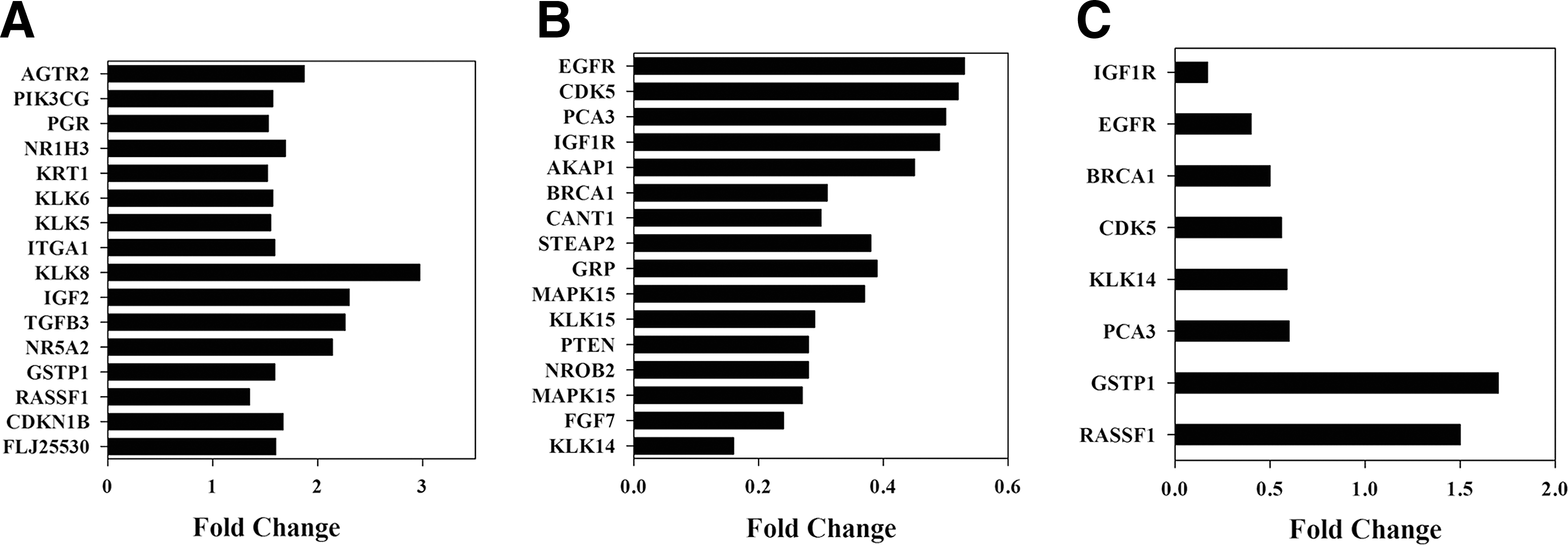
Further analysis of microarray using spot densitometry shows alterations in the gene expression profile of lutein-treated cells compared with control as demonstrated in the heat map (Fig. 6A). Similarly, scatter plot analysis (Fig. 6B) showed significant changes in lutein-treated cells, compared with control, with dots above 1 indicating increase and dots below 1 representing decrease in gene expression. Scatter plot showing gene expression levels comparing control and lutein-treated cells displayed an r 2 value of 0.94 (Fig. 6C). Gene cluster analyses also showed changes in the expression of proliferation, cell cycle, and apoptosis-associated genes in lutein-treated cells compared with control (Fig. 6D–F). An in-depth analyses of microarray data revealed that in vitro treatment with lutein resulted in a significant decrease in the expression of 15 selected growth and survival-associated genes in PC-3 cells. Among the upregulated genes, KLK8, IGF2, TGFB3, and NR5A2 showed greater than 2-fold increase and AGTR2, GSTP1, RASSF1, CDKNIB, FLJ, ITGA1, KLK5, KLK6, KRT1, NR1H3, PGR, and PI3KCG displayed 1.5- to 2-fold increase (Fig. 7A). Among the downregulated genes, KLK14, FGF7, MAPK15, NROB2, PTEN, and KLK15 showed more than 3-fold decrease and CANT1, BRCA1, MAPK6, STEAP2, GRP, AKAP1, IGF1R, PCA3, CDK5, and EGFR displayed 1.6- to 3-fold decrease after treatment with lutein (Fig. 7B). Further validation of selected genes by qRT-PCR confirmed that IGF1R, EGFR, BRCA1, CDK5, KLK14, and PCA3 showed a decrease, whereas GSTP1 and RASSF1 showed an increase in PC-3 cells treated with lutein compared with control (Fig. 7C). These results demonstrate that lutein inhibits the expression of growth and survival-associated biomarker genes in prostate cancer.
Discussion
The health benefits of lutein have been explored to find alternative therapies for inflammation and cancer. 18 Earlier studies have shown that lutein inhibits the production of nitric oxide and reactive oxygen species in macrophages demonstrating its anti-inflammatory property. 18,19 Lutein also induced cell cycle arrest in prostate cancer cells. 6,20 However, the effect and mechanism of action of lutein in prostate cancer is not fully defined. In this study, we found only a mild inhibition of proliferation by lutein in PC-3 cell. Therefore, we then examined the potentiating effect of lutein on Doxorubicin and Taxol, 9,10 two clinically available chemotherapeutic agents for prostate cancer. We found that Doxorubicin and Taxol induce antiproliferative response in PC-3 cells and that further improved in combination treatment with lutein by augmenting cell cycle arrest and apoptosis. Moreover, Temozolomide, which has no antiproliferative effect on PC-3 cells, significantly inhibited proliferation in combination with lutein by augmenting cell cycle arrest and apoptosis. PPARγ is a nuclear receptor transcription factor that regulates growth and differentiation of many cell types. Earlier studies have also shown that PPARγ agonists exert antitumor effects in many tumors, including prostate cancer. 13 –17 We showed earlier that PPARγ agonists inhibit the growth and expansion of brain tumor stem cells by modulating stemness and differentiation genes in glioma. 13 In addition, we have also shown that PPARγ agonists induce growth arrest and apoptosis in combination with lycopene in prostate cancer. 21 In this study, we found that PPARγ agonists inhibited the proliferation of PC-3 cells and lutein improved their effect by augmenting cell cycle arrest and apoptosis.
To understand the mechanisms by which lutein attenuates prostate cancer, we examined the expression of biomarker genes using gene array and qRT-PCR techniques. We found that in vitro treatment with lutein induced greater than two-fold decrease in the expression of IGF1R and BRCA1 in prostate cancer cells compared to control. The type I insulin-like growth factor receptor (IGF1R) is known to initiate a cascade of events leading to cell proliferation, neoplastic transformation, and tumorigenesis and is overexpressed in prostate, lung, colon, and breast cancer cells. 22,23 Circulating levels of IGF1R are associated with increased risk of breast, colon, and prostate cancers and high plasma IGF-I levels predispose to prostate cancer. IGF1R was significantly upregulated at the protein and mRNA level in primary prostate cancer. Moreover, overexpression of mutated BRCA1 associates with an aggressive form of prostate cancer. 24,25 The relative risk of prostate cancer is higher in male BRCA2 and BRCA1 mutation carriers, meanwhile the histological characteristics of this malignancy in these groups have not been clearly defined. Since BRCA1/2 mutation carrier status is associated with more aggressive disease, it is a prognostic factor for prostate cancer outcome. Targeted screening of this population may detect disease at an earlier clinical stage, which may, therefore, be beneficial.
We have also found approximately a two-fold decrease in the expression of EGFR, KLK14, PCA3, and CDK5 in PC-3 cells treated with lutein compared to control. Earlier studies have also implicated the association of EGFR, 26 –28 KLK14, 29,30 PCA3, 31 –33 and CDK5 34 in the progression of prostate cancer. Amplification and overexpression of EGFR and c-erbB-2 have been implicated in prostate cancer progression, thus EGFR-targeted drugs could be of therapeutic relevance in prostate cancer. KLK14 is a newly discovered human kallikrein gene that is mainly expressed in the central nervous system and endocrine tissues. Many members of the human kallikrein gene family are differentially expressed in cancer and a few have potential as diagnostic/prognostic markers. KLK14 was found to be regulated by steroid hormones in prostate cancer cell lines and differentially expressed in prostate cancer compared to normal tissue. 29,30 Cyclin-dependent kinase 5 (CDK5), a known regulator of migration in neuronal development, plays an important role in prostate cancer motility and metastasis. 34 Earlier studies have identified the prostate cancer antigen 3 (PCA3) as a perfect biomarker gene for the early detection of prostate cancer. 31 –33
In addition, lutein enhanced the expression of RASSF1 and GSTP1 ∼1.5-fold in PC-3 cells compared to control. Earlier studies have shown that RASSF1 and GSTP1 are silenced in prostate cancer. 35,36 Prostatic carcinoma is characterized by the silencing of the glutathione S-transferase gene (GSTP1), which encodes a detoxifying enzyme. The silencing of GSTP1 results from aberrant methylation at the CpG island in the promoter-5′ and occurs in the vast majority of cases of high-grade prostatic intraepithelial neoplasia and prostate cancers. The loss of expression of the GSTP1 gene is the most common (>90%) genetic alteration reported to date in prostate cancer. 37 Earlier studies have also identified the RAS-association domain family 1, isoform A gene (RASSF1A) located at 3p21.3 as a tumor suppressor gene. RASSF1A falls into the category of genes frequently inactivated by methylation rather than mutational events. This gene is silenced and frequently inactivated by promoter region hypermethylation in many adult and childhood cancers, including lung, breast, kidney, gastric, bladder, neuroblastoma, medulloblastoma, gliomas, and prostate cancer. 35,36 RASSF1A plays an important role in cell cycle regulation, apoptosis, and microtubule stability, and inhibits tumor growth in the in vitro and in vivo systems. 35 In this context, the upregulation of GST1 and RASSF1 by lutein observed in this study is important to understand its mode of action in prostate cancer. It is possible that the methylation-specific changes in GSTP1 could be a mechanism by which lutein improves sensitivity to TMZ and other chemotherapeutic agents in prostate cancer.
A number of genes have been implicated in prostate cancer by genetic alterations and functional consequences of the genetic alterations. The genes that have been identified need additional genetic, functional, and/or biochemical examination. However, the identification and characterization of these genes will be a key step for improving the detection and treatment of prostate cancer. Overall, this study demonstrates that lutein attenuates prostate cancer by altering the expression of growth and survival-associated genes. Thus, lutein has the potential to treat prostate cancer in conjunction with existing therapies. In addition, targeting the lutein sensitive biomarker genes could be a novel strategy to decrease the risk, delay the onset, or prevent the progression of prostate cancer.
Footnotes
Author Disclosure Statement
The authors declare no competing interests exist.