
Abstract
Oxidative stress (OS) promotes bone loss after menopause, and there is evidence that dietary antioxidants may reduce the level of OS in vivo. This study examined dose-dependent effects of blackberries (BBs) containing mainly cyanidin 3-O-β-
Introduction
P
The principal goal of osteoporosis therapy is to reduce fracture risk. To this end, drug therapies such as alendronate, risedronate, zoledronic acid, denosumab, and strontium ranelate used with calcium and vitamin D have been effective. 13 Nonetheless, due to factors such as cost, fear for side effects of medication, and experiences of side effects associated with these drugs, about 50% of patients discontinue medication or fail to adhere to the treatment regimens in the first year of use, which results in increased fracture risk. 13,14 Hence, in addition to drugs and lifestyle modifications, practical diet-based approaches to reduce the level of OS and improve bone health are necessary. Growing evidence shows that increasing the intake of fruits and vegetables, as part of a nutrient-balanced diet, may be beneficial to bone health. 15 –19 Additionally, experiments with dietary antioxidants (e.g., N-acetyl cysteine, ascorbic acid, alpha-tocopherol, and β-carotene) have demonstrated their potential to promote bone formation 20 and increase levels of enzymatic antioxidants. 8,16 These antioxidants may stimulate osteoblast differentiation, suppress osteoclast formation, decrease TNF-α production, and hence attenuate the bone resorptive effects of OS. 8,20,21
Apart from the traditional dietary antioxidants such as vitamins A, C, and E, phenolic compounds from plums and blueberries have demonstrated protective effects against bone resorption due to their strong superoxide scavenging capacity.
22
–24
Akin to these fruits is the blackberry (BB) fruit (Rubus sp.) that has a total phenolics range of 114–1056 mg/100 g fresh weight.
25
The composition of BB includes anthocyanins,
26
ellagitannins,
27
flavonols, flavan-3-ols,
28
procyanidins,
29
phenolic acids,
25
and low amounts of lignans.
30
Most berries have several anthocyanins in their phenolic profile; however, BB contains mostly the anthocyanin, cyanidin 3-O-β-
Using an Ovx-rat model of postmenopausal osteoporosis, effects of supplementation with C3G-rich BBs at the level of 5% (w/w) and 10% (w/w) were assessed on (i) bone mineral density (BMD), (ii) trabecular microarchitecture, (iii) bone biochemical markers, and (iv) plasma enzymatic antioxidants. An incomplete experimental model design of two factors and three level models was considered sufficient in this study because the object was to examine the effect of berries in the prevention of postmenopausal bone loss. The sham group served two purposes used to (i) verify that ovariectomy caused bone loss and (ii) investigate the extent to which the berries prevented bone loss.
Materials and Methods
Animal care and diet
Nine-month-old female (N=38) Sprague-Dawley rats (Harlan, Indianapolis, IN, USA), an established model for investigation of postmenopausal osteoporosis, 35 were used. Animals were housed in an environmentally controlled animal care facility with standard temperature and light:dark (12:12) cycles. Following a week of acclimatization, the animals were randomly assigned to four groups. One group was sham operated (Sham) and three groups were ovariectomized (Ovx). Baseline BMD and bone mineral content (BMC) were obtained using DXA dual X-ray absorptiometry (DXA; GE Lunar DXA, Waukesha, WI, USA) equipped with enCore software. The groups and corresponding diets were as follows: Sham+control diet (n=12), Ovx+control diet (n=12), Ovx+5% BB (n=7), and Ovx+10% BB (n=7). The control diet was an AIN-93M rodent diet, and the Ovx+5% BB and Ovx+10% BB were control diets modified to contain powdered and freeze-dried BB at levels of 5% and 10% (w/w). BB diets were formulated based on proximate analysis of freeze-dried BBs to contain the same amounts of carbohydrate, fiber, fat, and protein (data not shown). Rats were weighed on a weekly basis, received free access to deionized water, and were match fed to the weekly mean food intake of the sham group. Deionized water was used in this study to control the variation in the mineral content of the water as minerals in tap water may influence bone metabolism.
Animal necropsy
After the 100-day treatment period, the animals were placed in metabolic cages for a 24-h period to collect a 24-h urine sample. The urine sample was centrifuged at 3000 g for 20 min aliquoted and stored at −20 °C for deoxypyridinoline (DPD) and creatinine analyses. After a12-h fast, animals were anesthetized with a mixture of ketamine and xylazine (70 mg and 3 mg/kg body weight, respectively), to measure final BMD and BMC using DXA. The rats were then sacrificed by exsanguination through cardiac puncture. Blood samples were collected and centrifuged at 3000 g for 20 min at 4 °C. Serum was aliquoted and stored at −80 °C until analysis. Bone specimens, the tibiae, the fourth lumbar vertebrae, and femur, were collected, cleaned, and stored at −20 °C for BMD and microarchitectural analyses. Uteri were collected and weighed for confirmation of ovariectomy.
All animal procedures used in this study were according to the University of Arkansas Institutional Animal Care and Use Committee guidelines.
Assessment of bone mineral area, content, and density
Using procedures previously published 36 and DXA equipped with suitable software for assessing bone indices in small laboratory animals, bone mineral area (BMA), BMC, and BMD of the tibiae, the fourth lumbar vertebrae, and femur were measured.
Assessment of bone microarchitecture using microcomputed tomography
To assess the effect of C3G-rich BB treatment on the bone trabecular structure, the tibiae, fourth lumbar vertebrae, and femur were scanned using microcomputed tomography (μCT) and methods previously reported. 37 After analyzing the volume of interest, the following bone morphometric parameters were obtained: bone volume over total volume (BV/TV), trabecular number (Tb.N.), separation (Tb.Sp.), thickness (Tb.Th.), connectivity density, and structure model index (SMI).
Assessment of OS biomarkers
Effects of ovariectomy and C3G-rich BB treatments on OS levels were assessed using four assays. Thiobarbituric acid reactive substances (TBARS) were measured in plasma and used to determine lipid peroxidation, 38 an indicator of overall OS. TBARS were determined in duplicate using the TBARS assay kit (ZeptoMetrix Corporation, Buffalo, NY, USA) according to manufacturer's instructions and absorbance of 532 nm read on a Synergy HT (BioTek, Winooski, VT, USA) microplate reader.
The activity of antioxidant enzymes, glutathione peroxidase (GPx), superoxide dismutase (SOD), and catalase were assessed in plasma using BioVision's assay kits (BioVision, Inc., Mountain View, CA, USA) following manufacturer's instructions. All samples were assayed in duplicate and read using a Synergy HT (BioTek) microplate reader.
Assessment of systemic biomarkers bone metabolism
The effects of ovariectomy and the two doses of C3G-rich BB treatment were evaluated in four systemic biomarkers of bone metabolism. Alkaline phosphatase (ALP), which is required for the hydrolysis of phosphate esters, was assessed in serum as an indicator of bone formation using an Alfa Wassermann Diagnostic Technologies Kit and an ACE Alera Clinical Chemistry System (Alfa Wassermann Diagnostic Technologies, West Caldwell, NJ, USA). Serum osteocalcin (OC), a reliable indicator of bone turnover, 39 was measured using the Rat-MID™ Osteocalcin enzyme immunoassay (EIA) (Immunodiagnostic Systems, Inc., Fountain Hills, AZ, USA). Serum insulin-like growth factor-1 (IGF-1), a recognized modulator of bone metabolism involved in the proliferation of osteoblasts, 40 was quantified using an EIA kit from ALPCO immunoassays (ALPCO Diagnostics, Salem, NH, USA). In addition, a resorption bone biomarker urinary DPD was quantified using a competitive EIA kit specific for rats (Quidel, Mountain View, CA, USA). The assay is established in the DPD molecule specificity, and hence, cross-reaction with other collagen crosslinks such as pyridinoline is highly insignificant. 41 Urinary creatinine was measured colorimetrically using an ACE Alera clinical chemistry system and a commercially available kit from Alfa Wassermann Diagnostic Technologies (Alfa Wassermann Diagnostic Technologies). The final creatinine-corrected DPD results were expressed as nmols DPD/mmols creatinine. All serum and urine samples were assayed in duplicates for the above biomarkers.
Statistical analyses
The data analyses involved estimation of mean and standard error of the mean using JMP 8 (2009 SAS Institute, Inc., Cary, NC, USA). The effects of treatment were analyzed by one-way analysis of the variance model followed by post hoc analysis using the Fisher's least squares means separation test when F values were significantly different (P<.05).
Results
BMA, content, and density
Baseline BMC of the whole body was not different among groups, however, final BMC of the sham group was significantly lower than all groups (P<.0026). The cause for this finding remains unclear as typically ovariectomy should decrease BMC.
Baseline BMD of the whole body was not different among groups (Table 1). However, mild effects of Ovx were observed although insignificant in the final BMD values of Ovx rats when compared with the sham group (Table 1).
Values are mean±SEM. The effects of treatment were analyzed by one-way ANOVA model followed by post hoc analysis using the Fisher's least squares means separation test when F values were significantly different (P<.05). Within a row, values that do not share the same superscript letters (a or b) are significantly different from each other (P<.05).
Sham, sham operated; OVX, ovariectomized; OVX+BB 5%, ovariectomized+blackberry 5% (w/w); OVX+BB 10% (w/w), ovariectomized+blackberry 10% (w/w); BMC, bone mineral content; BMD, bone mineral density; SEM, standard error of the mean; ANOVA, analysis of variance.
There were no statistically significant differences in BMA of the tibia, fourth lumbar vertebra, and femur among groups. Results show that tibial BMC of the Ovx group was lower compared with the sham group, as expected (Table 2). In addition, although not to a statistically significant level, BMA and BMC of tibia, fourth lumbar vertebra, and femur of the Ovx+control group were lower to some degree compared with the sham group. Results also show that tibial, vertebral, and femoral BMDs were significantly reduced by Ovx, 5%, 5%, and 6%, respectively. Results of treatment show that a C3G-rich BB diet at the level of 5% and not 10% (w/w) increased BMD of the tibia, lumbar vertebra, and femur by 2.4%, 2.7%, and 4.3% (P<.0013; .0437; .0004), respectively, compared with the Ovx+control, bringing it up to the level of the sham group.
Values are mean±SEM. The effects of treatment were analyzed by one-way ANOVA model followed by post hoc analysis using the Fisher's least squares means separation test when F values were significantly different (P<.05). Within a row, values that do not share the same superscript letters (a, b, or c) are significantly different from each other (P<.05).
BMA, bone mineral area.
μCT analysis of tibia and fourth lumbar vertebra
Four cortical parameters were assessed at the proximal tibia, including the cortical area, porosity, thickness, and medullary area. Tibial cortical bone microarchitecture was markedly compromised by Ovx (Table 3). Ovx also resulted in decreased cortical thickness and cortical area by 7% and 8%, respectively. Effects of Ovx also resulted in an increase in the cortical percent porosity of 8.5%.
Values are mean±SEM. The effects of treatment were analyzed by one-way ANOVA model followed by post hoc analysis using the Fisher's least squares means separation test when F values were significantly different (P<.05). Within a row, values that do not share the same superscript letters (a or b) are significantly different from each other (P<.05).
Cort. Th., cortical thickness; Cort., cortical; Med., medullary.
The effect of C3G treatment at the level of 5%, but not at 10% (w/w), is demonstrated by increased cortical thickness by 5%, bringing it up to the level of the sham group. However, C3G treatments had no impact on the cortical area and cortical porosity. Neither Ovx nor C3G treatments had any effects on the cortical–medullary area. Data analysis of trabecular microarchitecture showed that Ovx considerably compromised all parameters of trabecular microarchitecture of the proximal tibia and fourth lumbar vertebra except tibial Tb.Th. (Fig. 1). Ovx-induced changes were characterized by decreased BV/TV (Fig. 1A), Tb.N. (Fig. 1B), and Tb.Th. (Fig. 1C) and increased Tb.Sp. (Fig. 1D). Ovx decreased tibial and vertebral BV/TV by 23% and 13%, respectively. C3G-rich BB treatment at the level of 5% (w/w) significantly increased tibial BV/TV by 37% and had no effect on lumbar BV/TV. The higher dose of BB 10% (w/w) had a negative impact to vertebral BV/TV significantly reducing BV by 28% compared to the Ovx+control.
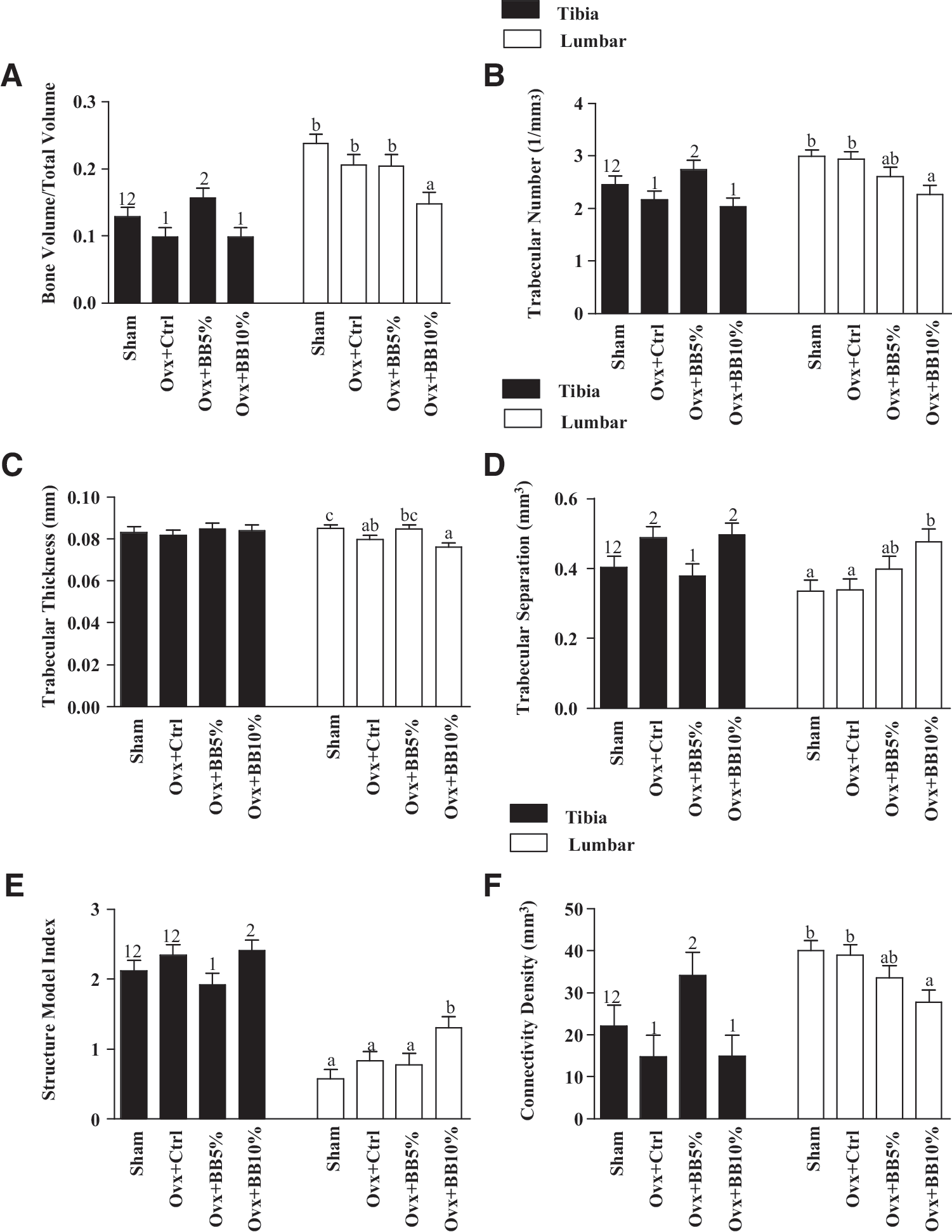
Effects of ovariectomy and dose-dependent C3G-rich blackberry treatments on the tibia, vertebrae, and trabecular bone. Graphs labeled
Similarly, tibial and vertebral Tb.N. was decreased in response to Ovx by 11% and 1.7%, respectively. C3G-rich BB at the level of 5% (w/w) resulted in significant positive effects on the tibial, but not the vertebral Tb.N. Tibial Tb.N. values significantly increased by 21% in the Ovx+BB 5% group, but the higher BB dose had no such effect. Ovx reduced Tb.Th. by 1.7% and 6.3% in the tibia and vertebra bones, respectively. In regard to treatments, the low BB diet dose increased vertebra Tb.Th. by 6% to the level of the sham group, but the higher dose had no effect, and neither treatment had an effect on tibial Tb.Th. Ovx also increased tibial and vertebra Tb.Sp. by 17% and the slight by 0.6%, respectively. C3G-rich BB treatment at the level of 5% (w/w) prevented tibial Tb.Sp. by reducing it by 22%, bringing it up to the sham level, but the same dose had no positive effects on the vertebral bone. Tibial and vertebral Tb.Sp. in animals fed BB at the level of 10% (w/w), similar to the Ovx control group.
Nonmetric parameters of the tibial and lumbar trabecular bone, that is, SMI and connectivity density (Fig. 1E, F) were modified by Ovx, although not to a statistically significant degree. Higher SMI indicates a more biomechanically compromised trabecular bone pattern, which is more rod-like as opposed to the favorable plate-like pattern. Tibial SMI increased by 9.4%, and that of the vertebrae by 31%, while connectivity density decreased in both bones by 32% and 2.6%, respectively, due to Ovx. Although the impact of C3G-rich BB 5% (w/w) on tibial SMI did not reach significance level, this dose modestly prevented the change of trabecular to the unfavorable rod-like pattern by reducing the index by 18% compared with the Ovx+control. The vertebra SMI was not changed by the lower dose of BB treatment, but the high dose significantly increased SMI. Tibial but not lumbar connectivity density in animals fed a low dose of C3G-rich BB significantly increased to a value higher than the Ovx and sham groups. The higher dose of C3G-rich BB treatment had no positive effects on the connectivity density of both bones.
Biomarkers of OS and bone metabolism
Effects of Ovx and C3G-rich BB treatments were assessed on three antioxidant enzymes, SOD, GPx, catalase, and a biomarker of OS, TBARS. Neither Ovx nor C3G-rich BB treatments had any significant effects on these biomarkers.
Urinary DPD and serum OC, ALP, and IGF-I were measured to assess their response to Ovx and C3G-rich BB treatments on bone resorption and bone formation, respectively (Table 4). DPD expressed per unit of creatinine was increased by Ovx, but C3G-rich BB treatments did not suppress the urinary excretion of these collagen by-products. Serum OC was significantly increased in the Ovx+control group (37%) in response to Ovx, but not altered by C3G-rich BB treatments. ALP was significantly higher in the Ovx rats compared with the sham+control group and C3G-rich BB treatment at the level of 10%, but not 5% (w/w), decreased the ALP level compared with the level of the sham group. In response to Ovx, IGF-1 increased by 9%, and C3G-rich BB treatments further enhanced these levels in a dose-dependent manner, however, these changes represent only a trend and did not reach the level of statistical significance.
Values are mean±SEM. The effects of treatment were analyzed by one-way ANOVA model followed by post hoc analysis using the Fisher's least squares means separation test when F values were significantly different (P<.05). Within a row, values that do not share the same superscript letters (a or b) are significantly different from each other (P<.05).
SOD, superoxide dismutase; GPx, glutathione peroxide; TBARS, thiobarbituric acid reactive substances; DPD, deoxypyridinoline; ALP, alkaline phosphatase; IGF, insulin-like growth factor.
Discussion
Bone loss in response to Ovx is partially due to increased OS and impaired antioxidant systems of the bone tissue. 12,42 In this study, bone loss occurred at all sites measured in varying magnitudes. Changes are attributable to diet, the control diet was AIN-93M rodent diet, while experimental diets were control diet modified to contain powdered freeze dried-BB at levels of 5% and 10% (w/w). All other parameters were controlled. Study shows modest protection of tibial and femoral BMD by C3G-rich BB 5% (w/w) that could reduce fracture risk at these sites, the primary goal in osteoporosis therapy. Such results are encouraging considering that the risk of any fracture in women is about twice as high as that of men. 13 Similar bone protective effects have been demonstrated in blueberries, which have an equally high antioxidant capacity. 43
μCT analyses indicated Ovx-induced changes of the cortical and trabecular microarchitecture consistent with findings in other studies. 37,44,45 Moreover, evaluation of the metaphyseal region of the tibia demonstrated C3G-rich BB 5% (w/w) protection against loss of BV/TV and Tb.N. As a consequence of preserved Tb.N., less Tb.Sp. and more connectivity were observed. Tibial Tb.Th. remained unaltered by both Ovx and treatment. We speculate that no effect on Tb.Th. resulted from a compensatory mechanism to correct for loss in connectivity, as proposed previously, 37 and/or lack of statistical power due to the limited sample size in this study.
Rod-like trabeculae are more susceptible to fracture than plate-like patterns. 46 Therefore, by preventing rod-like pattern formation, as evidenced by lower SMI values, C3G-rich BB 5% (w/w) could reduce fracture risk. Such findings are important in providing evidence that supports epidemiological studies 18,47 that associate consumption of fruits and vegetables to improved bone health. Interestingly, response to treatment at the lumbar vertebrae was weak and these findings agree with previous studies. 37,48 However, compared to the proximal tibia, this site was also less vulnerable to the trabeculae deterioration effects of Ovx.
In terms of biomarkers, concentrations of urinary DPD increased the characteristic of bone turnover in favor of resorption due to Ovx. However, C3G-rich BB diets did not suppress the bone resorption rate, as would be expected. Also, serum markers of bone formation ALP and OC both of which indicated increased bone turnover following Ovx were unaltered by BB treatment. Ovx and BB treatments had no effect on IGF-1. Suppression of Ovx-induced ALP and OC has been reported in blueberry treatment, 43 but not in dried plum. 44 Similar to dried plum treatment, 44 C3G-rich BB diets were unable to alter bone mineralization raising the question as to the mechanism of action. Systemic markers appear to differ with treatments and mechanism of action, therefore further investigations are necessary to elucidate the mechanisms by which C3G-rich BB prevent Ovx-induced bone loss.
Following Ovx, rats suffer depleted intracellular antioxidants, such as glutathione, and experience increased osteoclast activity. 12 OS has been shown to result in marked regulation and expression of antioxidant enzymes, including SOD, GPX, and catalase. 5,49 However, this study found no significant changes in these enzymes as a result of either Ovx or C3G-rich treatments. Results on Table 4 show a slight dose-dependent trend in reduction of SOD, GPX, and catalase, which may be indicative of a reduced need for their expression as a result of reduced OS by C3G-rich BB. TBARS, an indicator of OS, was not statistically significant between groups.
The anthocyanin concentration in the BBs in this study as determined by high-performance liquid chromatography and high-performance liquid chromatography/electrospray ionization tandem mass spectrometry was 631.3 mg/100 g of dry weight and C3G was 87% of total anthocyanins. Therefore, we can hypothesize that the bone protective effects seen in this study are largely due to the high C3G content, which has one of the highest oxygen radical absorbance capacity activity compared to other anthocyanins. 50 It is plausible that the dose at 10% (w/w) was excessive, and therefore ineffective in combating OS. Future studies may experiment with freeze dried-BB at levels <5%, 5%, and 7.5% (w/w). Also, high fiber content in BBs may have inhibited the absorption of essential bone-building minerals such as calcium and phosphorous. However, the mechanisms by which C3G-rich BBs prevent bone loss need to be explored in future studies. To our knowledge, no other study has investigated the bone-protective effects of BB phenolic compounds.
Conclusion
In conclusion, BBs supplemented in the diet at the level of 5% (w/w) may modestly reduce ovarian hormone-induced bone loss evident by improved tibial, vertebral, and femoral BMD values, and tibial bone microstructural parameters. Bone protective effects may be as a result of the synergistic effects of phenolic compounds, however, further work is required to determine BBs' specific mechanisms of action.
Footnotes
Author Disclosure Statement
No competing financial interests exist.