
Abstract
The authors aimed in the present study to assess the protective effect of Rosmarinus officinalis essential oils (ROEO) and Lavandula stoechas essential oils (LSEO) against reproductive damage and oxidative stress in alloxan-induced diabetic male rats. Essential oil samples were obtained from the aerial parts of the plants by hydrodistillation and analyzed by the gas chromatography–mass spectrometry (GC-MS). Rats were divided into four groups: healthy control (HC); diabetic control (DC); healthy+ROEO (H+ROEO), healthy+LSEO (H+LSEO), diabetic+ROEO (D+ROEO), and diabetic+LSEO (D+LSEO). The use of GC-MS allowed to the identification of 15 and 22 compounds in ROEO and LSEO, respectively. In addition, the 2,2-diphenyl-1-picrylhydrazyl (DPPH) test showed that ROEO and LSEO had an important antioxidant capacity. In vivo, we initially found that ROEO and LSEO treatment protected against the decrease in alloxan-induced body weight gain, relative reproductive organ weights, testosterone level, as well as sperm quality decline. On the other hand, we showed that alloxan administration was accompanied by an oxidative stress status assessed by an increase of malondialdehyde (MDA) and hydrogen peroxide (H2O2) levels, as well as a depletion of sulfhydril group content (-SH) and antioxidant enzyme activities as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) in testis, epididymis, and sperm. More importantly, ROEO and LSEO treatment significantly protected against oxidative damage of the male reproductive organ systems in alloxan-induced diabetic rats. These findings suggested that ROEO and LSEO exerted a potential protective effect against alloxan-induced reproductive function damage and oxidative stress in male rat. The beneficial effect of ROEO and LSEO might be related, in part, to their antioxidant properties.
Introduction
D
Rosmarinus officinalis and Lavandula stoechas are well-known medicinal plant species in the world, including Tunisia, for their beneficial health effects. However, due to their richness in therapeutically active compounds, 10 these plants present many beneficial health effects as antioxidant 10 and antidiabetic 11 activities. Recently, we have shown that Lavandula stoechas essential oils (LSEO) attenuate hyperglycemia and protect against liver and kidney oxidative stress in alloxan-induced diabetic rats. 10
Hence, the present study aimed to investigate the possible implication of oxidative stress in the protective effect of Rosmarinus officinalis essential oils (ROEO) and LSEO against damage induced by alloxan treatment in the reproductive function of male rats.
Materials and Methods
Chemicals
5,5′-dithiobis(2-nitrobenzoic acid) (DTNB), trichloroacetic acid (TCA), acetylcholine iodide, S-butyrylcholine, butylhydroxytoluene (BHT), KOH, ethanol, ether, Coomassie G250, bovine serum albumin (BSA), orthophosphoric acid, and NaCl were obtained from Sigma-Aldrich Co.
Plant collection
L. stoechas and R. officinalis aerial parts were collected in March, 2010, from the area of Ain Draham (North-West of Tunisia) and identified by the laboratory of taxonomy in the Faculty of Sciences of Tunis (FST), Tunisia. The voucher specimens (No. R101 for R. officinalis and No. L101 for L. stoechas) have been deposited with the herbarium of the Higher Institute of Biotechnology of Béja and also in the Department of Biological Sciences, Faculty of Science of Tunis, Tunisia.
Plant extracts preparation
The dried aerial parts were submitted to hydrodistillation for 3 h using the Clevenger-type apparatus. Briefly, the plant was immersed in water and heated to boiling, after which the essential oils were evaporated together with water vapor and finally collected in a condenser. The distillate was isolated and dried over anhydrous sodium sulfate.
Free radical-scavenging activity on DPPH
The antioxidant capacity of the ROEO and LSEO was performed using 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical-scavenging activity (RSA) as previously described by Grzegorczyk et al.
12
Briefly, various concentrations of the essential oils (20, 50, 100, 150, and 200 μg/mL) were added to 1 mL of 0.1 mM methanol solution of DPPH and incubated at 27°C during 30 min. The absorbance of the sample was measured at 517 nm. DPPH RSA expressed as percentage was calculated using the following formula:
BHT was used as a reference molecule in the same concentration as the test extract.
All the analyses were done in triplicate. The efficacy concentration 50 (EC50) value was defined as the concentration (in μg/mL) of the compound required to scavenge 50% of the DPPH radical.
Gas chromatography–mass spectrometry
ROEO and LSEO were subjected to gas chromatography–mass spectrometry (GC-MS) analysis using the Trace GC ULTRA/Polaris Q (GC-MS; Thermo Electron). The column was a VB-5 (5% phenyl/95% dimethylpolysiloxane) with film thickness of 0.25 μm, a length of 30 m, and an internal diameter of 0.25 μm; helium was used as carrier gas. The GC oven temperature was kept at 50°C for 5 min and programmed to 250°C for 3 min at the rate of 4°C/min and programmed to 300°C at the rate of 25°C/min. The injector temperature was set at 250°C. Split flow was adjusted at 50 mL/min. MS were taken at 70 eV. Mass range was from uma 20 to 350. A library search was carried out using the Wiley GC-MS Library, NIST, and PMW. The sample was dissolved in hexane.
Animals and treatment
Adult male Wistar rats (weighing 220–230 g, 15 weeks old, and housed five per cage) were purchased from SIPHAT and used in accordance with the local ethics committee of the Tunis University for use and care of animals in conformity with the NIH recommendations. The animals were housed in standard cages (40×28×16 cm) under controlled conditions: 12-h light–12-h dark, 20–22°C, food and water are ad libitum. After diabetes induction, rats were divided into 6 groups of 12 animals each:
Group I: nondiabetic rats (control) treated with NaCl (0.9%, i.p.) during 15 days.
Group II: diabetic rats (alloxan, 160 mg/kg b.w.) treated with NaCl (0.9%) during 15 days.
Group III: nondiabetic rats treated with ROEO (150 mg/kg b.w., i.p.) during 15 consecutive days.
Group IV: nondiabetic rats treated with LSEO (50 mg/kg b.w., i.p.) during 15 consecutive days.
Group V: diabetic rats (alloxan, 160 mg/kg b.w., i.p.) treated with ROEO (50 mg/kg b.w., i.p.) during 15 consecutive days.
Group VI: diabetic rats (alloxan, 160 mg/kg b.w., i.p.) treated with LSEO (50 mg/kg b.w., i.p.) during 15 consecutive days.
Twenty-four hours after the last injection, animals were sacrificed, and blood was collected in heparinized tubes. After centrifugation at 3000 g for 15 min, plasma was processed for biochemical parameter determinations. The testis as well as the epididymis were rapidly excised and homogenized in phosphate-buffered saline and the sperm was collected in 5-cc tubes. After centrifugation at 10,000 g for 10 min at 4°C, supernatants were used for biochemical determination.
Induction of experimental diabetes
Experimental diabetes was induced in 12-h fasted rats by single i.p. injection of alloxan (160 mg/kg body weight) dissolved in the citrate buffer (100 mM, pH 4.5). To prevent fatal hypoglycemia, as a result of massive pancreatic insulin release, the rats were supplied with 10% glucose solution after 6 h of alloxan administration for the next 24 h. After 7 days, rats with glycemia ≥14 mM were selected for the experiment.
Body and reproductive organ weights
Initial (weight at the starting point) and final (weight at the end point) body weights were recorded. The reproductive organs were stripped from fatty tissues and blood vessels, blotted, and their absolute weights were determined. Clinical signs of body and reproductive organs were evaluated for toxicological criteria. To normalize the data for statistical analysis and to obtain relative weight, data were expressed per 100 g body weight.
Glycemia assays
Glucose was measured by the glucose oxidase and peroxidase using quinoneimine as a chromogen. The amount of glucose is related to the amount of quinoneimine, which is measured spectrophotometrically at 505 nm. 13
Plasma acetylcholinestrase and butyrylcholinesterase activities
The cholinesterase activities are assayed in plasma by the Ellman's method 14 using acetylthiocholine iodide as a substrate. The rate of hydrolysis of acetylthiocholine was measured at 405 nm by the reaction of thiocholine with dithiobis nitrobenzoic acid to give the yellow 5-thio-2-nitrobenzoate anion with the spectrophotometer. The enzyme activity was expressed as nmol of substrate hydrolyzed/min/mg protein.
Evaluation of serum testosterone
The concentration of plasma testosterone was measured by commercial enzyme-linked immunosorbent assay (ELISA) purchased from the Demeditec (Ref, DEV 9911), radioimmunoassay, and ELISA using commercial kits as per the manufacturer's instructions.
Evaluation of sperm characteristics
Sperm collection
After the test period, the laparotomy was conducted for all exposed and control male rats after body weight measurement. Testis and epididymis were carefully excised. The sperm count was assessed from right cauda epididymis, whereas sperm motility and morphology were analyzed from the left one. Epididymis was excised and minced in 1 mL of RPMI to obtain spermatozoa suspension. The other testis and epididymis were frozen in −20°C until use.
Sperm count
The cauda epididymal sperm count was performed according to the method of Vega et al. 15 Epididymal sperm counts were expressed as number of spermatozoa per epididymis. To minimize the error, the count was repeated three times on each sample. 16
Sperm motility
Ten microliters of sperm suspension was layered onto a warmed microscope slide. Sperm motility was assessed by counting all progressive motile, nonprogressive motile, and immotile spermatozoa. The number of motile spermatozoa in each field was divided by the total number and the average of the fields was assayed. The percentage of motile spermatozoa was thereafter determined. 17
Sperm viability
Sperm viability was assessed using the eosin stain. 18 The staining was performed with one drop of freshly collected semen (10 μL) and two drops of solution (20 μL) of eosin. The living spermatozoa remain intact so that the dead are purple with red stain. The dye exclusion was evaluated in 100 spermatozoa. Sperm viability was defined as the percentage of dead sperm cells. Viability was evaluated according to the WHO guidelines. 19
Sperm morphology
A drop of sperm suspension was smeared on a slide and air-dried and made permanent. The smeared slide was stained with 1% eosin. Morphological sperm defects were evaluated and examined on an optical microscope. 20 At least 100 spermatozoa from different fields in each slide were examined and classified for criteria of morphological abnormalities (head, tail, and tail-head) according to Filler. 21 Abnormal sperm cells were counted and the percentage calculated.
Assessment of sperm production
Sperm content per gram was determined using the method previously described by Vega et al. and Narayana et al. with slight modifications. 15,22 Briefly after thawing at temperature (25–27°C), the whole epididymal and the testicular tissues were homogenized for 5 min in 5 mL of physiological saline (0.9% NaCl) containing 0.05% (v/v) Triton X-100 using a manual homogenizer. The homogenates were diluted with 1.5 mL of the saline solution; spermatozoa and spermatids were counted. Three counts per sample were averaged. 23 These count values were used to obtain the total number of spermatids per testis or epididymis, which was then divided by the weight of the testis or epididymis to determine the number per gram of testis or epididymis.
Lipid peroxidation
Lipid peroxidation was detected by the determination of malondialdehyde (MDA) production determined by the method of Begue and Aust. 24 Briefly, homogenates of testis, epididymis, and sperm were centrifuged at 1000 g for 10 min at 4°C to sediment cell debris. Supernatants were suspended in phosphate-buffered saline, pH=7.4, mixed with BHT-TCA solution (1% BHT dissolved in 20% TCA w/v), centrifuged at 1000 g for 35 min and finally mixed with 0.5 N HCl and 120 mM thiobarbituric acid (TBA) in 26 mM Tris, and heated in water bath at 80°C for 10 min. After cooling, the absorbance of the resulting chromophore was measured at 532 nm. MDA levels were determined by using an extinction coefficient for MDA-TBA complex of 1.56×105 M−1 cm−1.
Thiol groups measurement
Total concentration of thiol groups (-SH) was performed according to the Ellman's method. 25 Briefly, aliquots from liver tissue was mixed with 100 μL of 10% sodium dodecyl sulfate and 800 μL of 10 mM phosphate buffer (pH 8), and the absorbance was measured at 412 nm (A0). Then, 100 μL of DTNB were added and incubated at 37°C during 60 min. After incubation, the absorbance of the sample was measured at 412 nm (A1). The thiol group concentration was calculated from A1 to A0 subtraction using a molar extinction coefficient of 13.6×103 M−1 cm−1. Results were expressed as nmol of thiol groups per mg of protein.
Antioxidant activities assays
Superoxide dismutase (SOD) activity was determined by using modified epinephrine assay of Misra and Fridovich. 26 At alkaline pH, superoxide anion O2 − causes the autoxidation of epinephrine to adrenochrome, whereas competing with this reaction, SOD decreased the adrenochrome formation. One unit of SOD is defined as the amount of the extract that inhibits the rate of adrenochrome formation by 50%. Enzyme extract was added to a 2 mL reaction mixture containing 10 μL of bovine catalase (0.4 U/μL), 20 μL epinephrine (5 mg/mL), and 62.5 mM sodium carbonate/bicarbonate buffer pH 10.2. Changes in absorbance were recorded at 480 nm. Characterization of SOD isoforms was performed using KCN (2 mM), which inhibits Cu/Zn-SOD or H2O2 (5 mM), affecting both Cu/Zn-SOD and Fe-SOD, whereas Mn-SOD was insensitive to both inhibitors. 27
Catalase (CAT) activity was assayed by measuring the initial rate of H2O2 disappearance at 240 nm. 28 The reaction mixture contained 33 mM H2O2 in 50 mM phosphate buffer with pH 7.0 and the CAT activity was calculated using the extinction coefficient of 40 mM−1 cm−1 for H2O2.
Glutathione peroxidase (GPx) activity was measured by the procedure of Flohé and Günzler. 29 Briefly, 1 mL of the reaction mixture containing 0.2 mL of sample supernatant, 0.2 mL of phosphate buffer 0.1 M pH 7.4, 0.2 mL of reduced glutathione (GSH) (4 mM), and 0.4 mL of H2O2 (5 mM) was incubated at 37°C for 1 min and the reaction stopped by the addition of 0.5 mL TCA (5%, w/v). After centrifugation at 1500 g for 5 min, aliquot (0.2 mL) from supernatant was mixed with 0.5 mL of phosphate buffer 0.1 M pH 7.4 and 0.5 mL DTNB (10 mM) and absorbance was recorded at 412 nm. GPx activity was expressed as nmol of GSH consumed/min/mg protein.
Protein determination
Protein concentration was determined according to the Bradford method 30 using BSA as standard.
Statistical analyses
Data were analyzed by unpaired Student's t-test or one-way analysis of variance (ANOVA) and were expressed as means±standard error of the mean. Data are representative of 10 independent experiments. All statistical tests were two-tailed, and a P-value of .05 or less was considered significant.
Results
Chemical composition of ROEO and LSEO
The results obtained by GC-MS analysis of the ROEO are presented in Table 1 and revealed the presence of 15 compounds. The principal compounds detected are: 1,8-Cineole (34.38), trans7caryophyllene (14.47), Borneol (9.66), Camphor (8.97), α7pinene (7.83), α7thujone (6.92). The LSEO are also previously analyzed by GC-MS and allowed to the identification of 22 compounds. 10
IR, retention index.
Antioxidant capacity of ROEO and LSEO
Concerning the antioxidant capacity, we showed that ROEO and LSEO present an important RSA (EC50=221.43±4.27 and 247.26±2.63 μg/mL, respectively), but lesser than BHT (EC50=108.24±3.78 μg/mL) used as reference molecule (Table 2).
EC50 is the effective concentration of sample that can decrease DPPH concentration by 50%.
BHT, butylhydroxytoluene; DPPH, 2,2-diphenyl-1-picrylhydrazyl; LSEO, Lavandula stoechas essential oils; ROEO, Rosmarinus officinalis essential oils; RSA, radical-scavenging activity.
Mortality and macroscopic symptoms of toxicity
In the present study, no mortality was noted until the end of the experiment in control and treated groups. Concerning the testis as well as the reproductive accessories, no macroscopic alterations were found throughout the experiment. However, cholinergic signs of adverse toxicological effects were observed such as sluggishness, muscular tremors, irregular movements, and abdominal tremble in the diabetic group. The progression of these signs proceeds to the last week of treatment. ROEO and LSEO administration reversed all adverse cholinergic effects induced by alloxan intoxication.
Body and reproductive relative organ weights
Data from Table 3 showed that diabetic rats showed a significant decrease of body weight gain. Alloxan treatment also increased the relative weights of testis, epididymis, prostate, and seminal vesicles, respectively, by 58%, 17%, 13%, and 44%. ROEO and LSEO administration significantly protected against alloxan-induced body weight decrease as well as the weight increase of the reproductive related organs.
Animals were treated during 15 days with ROEO (150 mg/kg b.w., i.p.), LSEO (50 mg/kg b.w., i.p.), or vehicle (NaCl 0.9%) 7 days after diabetic induction with alloxan at 160 mg/kg b.w. Assays were carried out in triplicate.
P<.05 compared with HC group.
P<.05 compared with DC group.
DC, diabetic control; HC, healthy control.
Evaluation of reproductive performance quality
The effects of alloxan and essential oils on reproductive performance quality are reported in Table 4. As expected, alloxan administration induced a decrease of epididymal spermatozoa count and testicular spermatid enumeration, as well as a reduction of sperm motility, whereas the number of dead sperm increased significantly. ROEO and LSEO administration reversed all reproductive performance and semen abnormalities induced by alloxan treatment.
Animals were treated during 15 days with ROEO (150 mg/kg b.w., i.p.), LSEO (50 mg/kg b.w., i.p.), or vehicle (NaCl 0.9%) 7 days after diabetic induction with alloxan at 160 mg/kg b.w. Assays were carried out in triplicate.
P<.05 compared with HC group.
P<.05 compared with DC group.
Blood glucose level
Significant increase in blood glucose level was observed in alloxan-induced diabetic rats compared with healthy nondiabetic animals (Table 5). ROEO and LSEO treatment protected against this increase, whereas essential oils alone had no significant effect.
Animals were treated during 15 days with ROEO (150 mg/kg b.w., i.p.), LSEO (50 mg/kg b.w., i.p.), or vehicle (NaCl 0.9%) 7 days after diabetic induction with alloxan at 160 mg/kg b.w. Assays were carried out in triplicate.
P<.05 compared with HC group.
P<.05 compared with DC group.
AChE, acetylcholinestrase; BChE, butyrylcholinesterase.
Evaluation of testosterone level as well as AChE and BChE activities
Concerning the biochemical findings, we are also interested on the cholinesterase activities and the testosterone level in the plasma compartment (Table 5). We showed that alloxan treatment significantly reduced the plasma acetylcholinestrase (AChE) and butyrylcholinesterase (BChE) activities as well as the testosterone level. ROEO and LSEO administration significantly protected against alloxan-induced plasma AChE and BChE activities and testosterone level depletion.
Lipid peroxidation, thiol group's content, and H2O2 level.
We further looked at the effect of alloxan and essential oils on oxidative stress condition (Fig. 1). We initially showed that diabetes induced by alloxan treatment is accompanied by a significant increase of testis, epididymis, sperm MDA, and H2O2 levels, as well as a decrease of thiol group content. Treatment with ROEO and LSEO protected against lipoperoxidation, hydrogen peroxide increase, as well as (-SH) groups decrease induced by alloxan treatment.
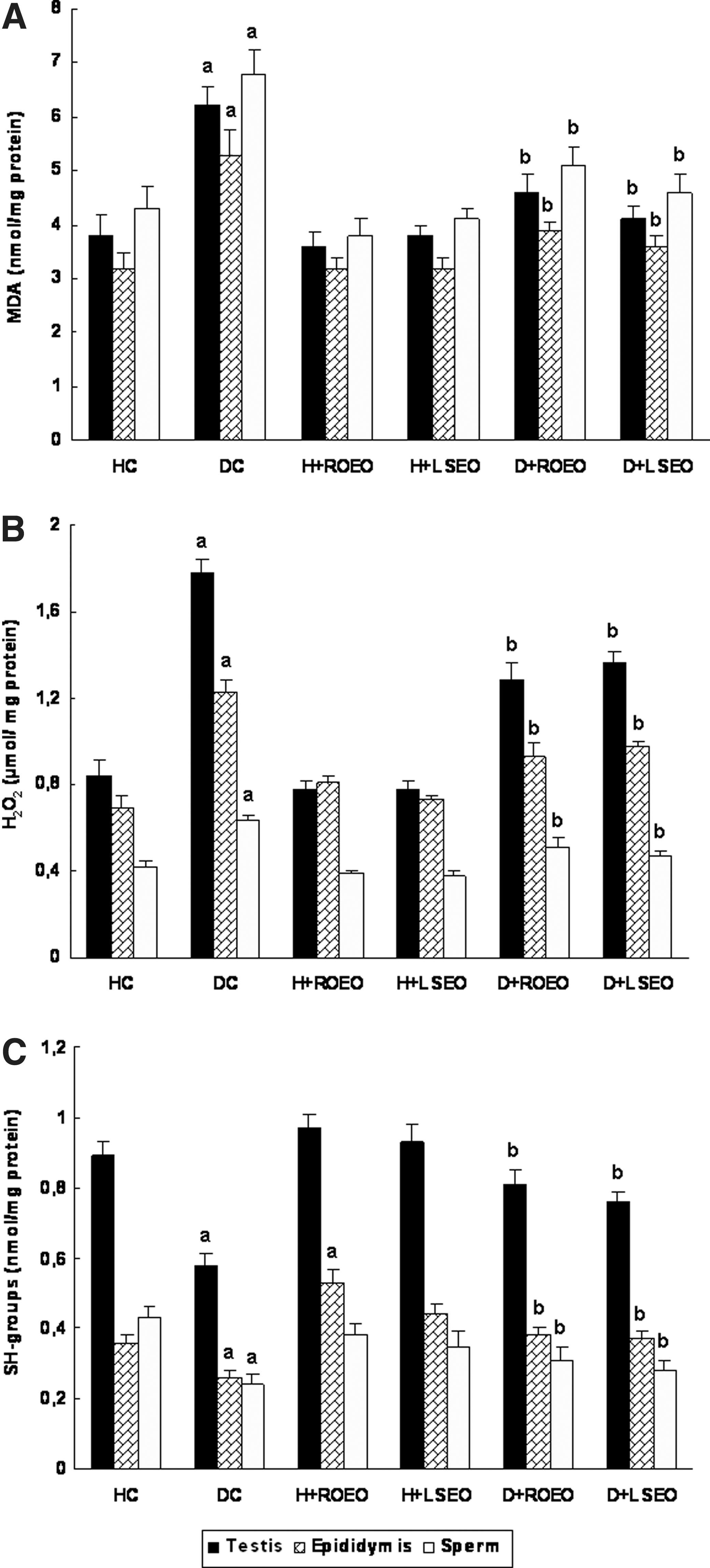
Subacute effect of Rosmarinus officinalis essential oils (ROEO) and Lavandula stoechas essential oils (LSEO) on acute alloxan-induced changes in malondialdehyde,
Antioxidant enzyme activities
The effect of alloxan and essential oils on antioxidant enzyme activities are reported in Figures 2 and 3. Alloxan per se significantly decreased testis and epididymis CAT, GPx, total SOD, Cu/Zn-SOD, and Mn-SOD activities. The subacute treatment with ROEO and LSEO protected against the depletion of antioxidant enzyme activities induced by alloxan intoxication. ROEO and LSEO alone significantly increased antioxidant enzyme activities.

Subacute effect of ROEO and LSEO on acute alloxan-induced changes in catalase (CAT)
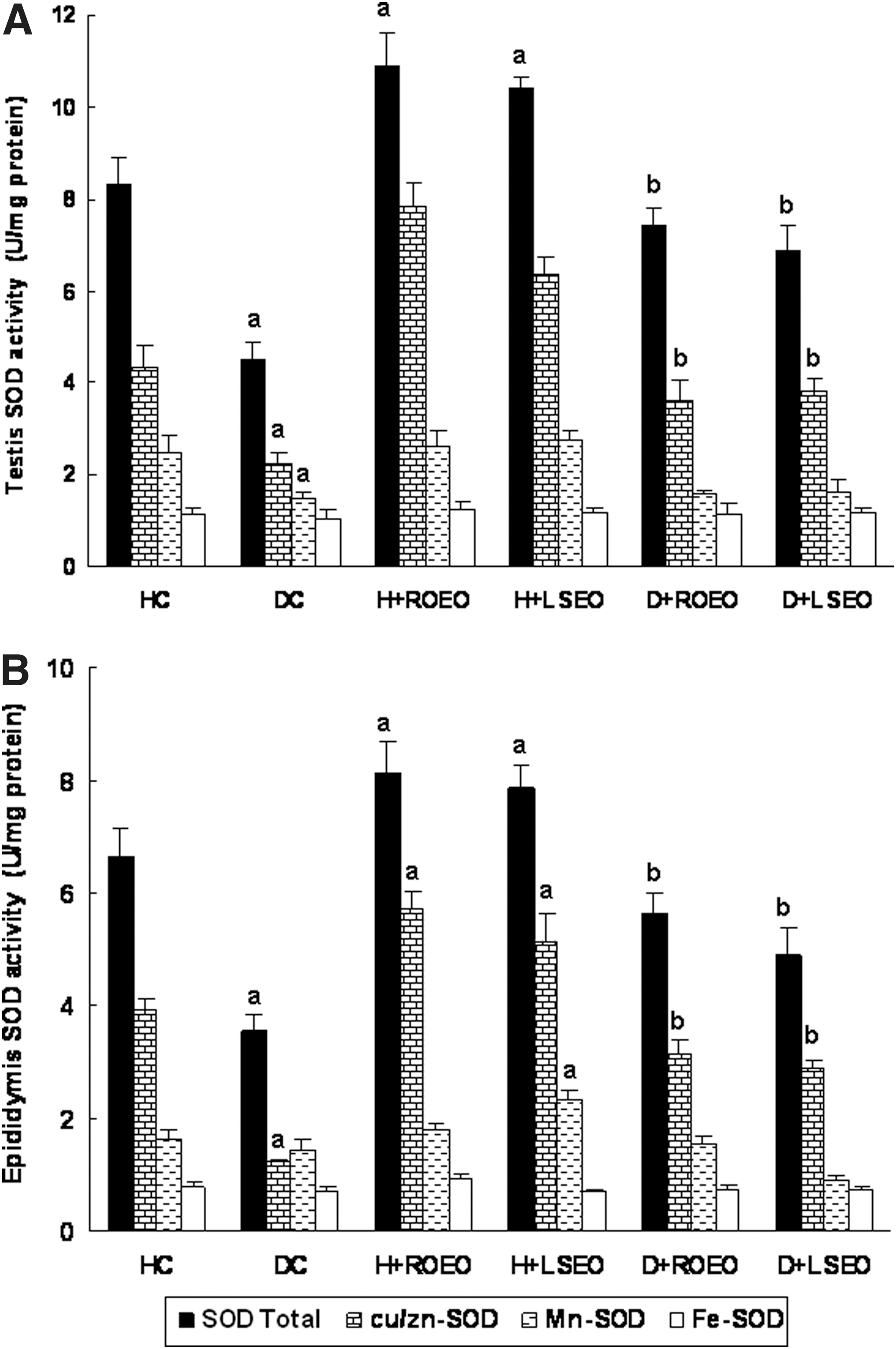
Subacute effect of ROEO and LSEO on acute alloxan-induced changes in testis
Discussion
In the present investigation, we studied the protective effects of ROEO and LSEO against alloxan-induced damage and oxidative stress in the reproductive function of male rats.
Phytochemical study of ROEO and LSEO by GC-MS analysis allowed to the identification of 22 and 15 compounds, respectively. The principal compounds detected in the ROEO are: 1,8-Cineole (34.38), trans7caryophyllene (14.47), Borneol (9.66), Camphor (8.97), α7pinene (7.83), α7thujone (6.92). However, the chemical composition of LSEO has been previously detailed in our previous report. 10 Our results generally corroborate the previous report with some discrepancies. 31 These changes in the essential oils composition might be related to the plant parts, seasons, and sites as previously described for Salvia officinalis. 32 On the other hand, using the DPPH radical-scavenging assay, we showed that ROEO and LSEO presented a higher scavenging capacity, which may be related to the presence of phenolic compounds, according to previous data. 31
In vivo, we initially found that alloxan administration increased glycemia. However, the major characteristics of DM are polydipsia, polyuria, polyphagia, weight loss, muscle weakness, and hyperglycemia. 33 Alloxan, a beta cytotoxin, destroys beta cells in pancreatic islets resulting in a decreased endogenous insulin secretion leading to a decreased utilization of glucose by body tissues. 34 It results also in the elevation of blood glucose level, decreases protein content, and increases the levels of cholesterol and triglycerides. 35
Importantly, we showed in the present study that ROEO and LSEO administration reversed all reproductive performance and semen abnormalities induced by alloxan treatment. However, to our knowledge, our study is the first one to deal with ROEO and LSEO effect on reproductive damage induced by alloxan administration. We also showed that ROEO and LSEO significantly reversed the depletion of plasma AChE and BChE activities induced by alloxan administration. However, our results corroborated the previous report indicating that alloxan treatment is implicated in the cholinesterase activity depletion. 36 The decrease of cholinesterase activity might be due, in part, to the inhibition of the enzyme synthesis by the altered brain cells. 37
We next sought to determine the putative involvement of oxidative stress conditions in the protective effect of ROEO and LSEO against alloxan-induced damage in the reproductive male function of rats. Alloxan administration induced oxidative stress as assessed by an increase of lipid peroxidation and hydrogen peroxide level, a decrease of sulfhydril content, as well as a depletion of antioxidant enzyme activities as CAT, GPx, total SOD, Cu/Zn-SOD, Mn-SOD, and Fe-SOD. Alloxan-induced oxidative stress has been previously reported in many organ systems such as liver, 10 kidney, 10 brain, 38 and testis. 39 Reactive oxygen species (ROS) play a relevant role in the pathogenesis of DM and its complications. Lipid peroxide-mediated tissue damage has been demonstrated in insulin-dependent and noninsulin-dependent DM. 40 As a strategy to counteract the negative effect of oxidative stress, antioxidant-based therapy is promising to minimize the complications in DM.
In this respect, our results showed that ROEO and LSEO protect against oxidative stress induced by alloxan administration in the reproductive organs of male rats. Alloxan-induced reproductive function damage and oxidative stress have been previously showed to be attenuated by Ajuga iva extract. 41 The chemical composition of ROEO and LSEO clearly demonstrated their richness in antioxidant molecules, which are the primal source of their antioxidant ability, by scavenging free radicals as hydroxyl radical (OH•), which is the major cause of lipid peroxidation. 42 It is well known that sperm cell membranes are rich in polyunsaturated fatty acids and are very susceptible to free radical attack. Lipid peroxidation of sperm cell membranes is one type of cell damage induced by ROS, which causes an increase in membrane permeability, an interruption in respiratory chain and ATP production, as well as a decrease in phosphorylation of axonemal proteins. 43 Alloxan-induced decrease of the sperm quality might be due to the depletion of antioxidant enzyme activities and/or ROS accumulation. Free radical-induced sperm motility decrease was presumably due to a rapid loss of intracellular ATP, which altered axoneme structure and caused tail abnormality. 44 Injury of cell DNA induced by excessive ROS led to the production of some peroxidation products such as 8-oxo-7,8-dihydroxyguanosine, which cause fragmentation and have a mutagenic effect. 45 High levels of sperm DNA damage have been reported to decrease male fertility and antioxidants could be used to alleviate male infertility. 46
In conclusion, our data clearly demonstrated that ROEO and LSEO exert protective effects against reproductive damage and oxidative stress in alloxan-induced diabetic rats owing, in part, to their antioxidant properties.
Footnotes
Acknowledgment
Financial support of the Tunisian Ministry of Enseignement Supérieur et Recherche Scientifique is appreciatively acknowledged.
Author Disclosure Statement
This research received no specific grant from any funding agency of the public, commercial, or nonprofit sectors.