
Abstract
Garlic and its lipid-based extracts have played an important medicinal role in humans for centuries that includes antimicrobial, hypoglycemic, and lipid-lowering properties. The present study was to investigate the effects of crude garlic extract (CGE) on the proliferation of human breast, prostate, hepatic, and colon cancer cell lines and mouse macrophageal cells, not previously studied. The human cancer cell lines, such as hepatic (Hep-G2), colon (Caco-2), prostate (PC-3), and breast (MCF-7), were propagated at 37°C; air/CO2 (95:5 v/v) using the ATCC-formulated RPMI-1640 Medium and 10% fetal bovine serum (FBS), while the mouse macrophage cell line (TIB-71) was propagated at 37°C; air/CO2 (95:5 v/v) using the ATCC-formulated DMEM and 10% FBS. All cells were plated at a density of ∼5000 cells/well. After overnight incubation, the cells were treated with 0.125, 0.25, 0.5, or 1 μg/mL of CGE an additional 72 h. Inhibition of cell proliferation of 80–90% was observed for Hep-G2, MCF-7, TIB-71, and PC-3 cells, but only 40–55% for the Caco-2 cells when treated with 0.25, 0.5, or 1 μg/mL. In a coculture study of Caco-2 and TIB-71 cells, inhibition of cell proliferation of 90% was observed for Caco-2 cells compared to the 40–55% when cultured separately. CGE also induced cell cycle arrest and had a fourfold increase in caspase activity (apoptosis) in PC-3 cells when treated at a dose of 0.5 or 1 μg/mL. This investigation of CGE clearly highlights the fact that the lipid bioactive compounds in CGE have the potential as promising anticancer agents.
Introduction
O
Over many decades, researchers have been studying plants and vegetables for identifying naturally occurring chemopreventative agents highly effective in preventing carcinogenesis, which could be used in combination with existing drug therapies. 6 –11 Garlic, also known as Allium sativum, is a vegetable that is currently used as a flavoring agent in cooking and has played an important medicinal role in humans for centuries. 12 Some chemical constituents of garlic have previously been shown to be active as antimicrobial, hypoglycemic, and lipid-lowering agents. 13 –17 Various studies indicating garlic as a dietary anticarcinogen have been carried out with aged garlic extract (AGE), aqueous garlic extract, and dehydrated garlic powder. 14,18 –20 Garlic contains a mixture of organosulfur compounds (OSC) that are oil or water soluble and are known to suppress tumor growth in vitro and in vivo. 6 Several studies have shown that some of the garlic-derived OSC can inhibit cell proliferation of cultured cancer cells by causing apoptosis and cell cycle arrest. 21 –26 A study carried out by Pinto et al. 27 investigated the growth inhibitory effects of AGE on anchorage-dependent human breast cancer cell MCF-7 and showed that AGE exhibited anticancer properties against breast cancer cells that had retained the properties of human breast cancer clinically. Seki et al. 28 demonstrated the growth inhibition of human promyelocytic leukemia HL-60 cells with garlic oil. The mechanisms of cancer prevention by garlic include but are not limited to increased DNA repair, changes in gene expression, decreasing inflammation, suppression of cell growth, and induction of apoptosis. 29
The primary objective of this study was to investigate the effects of a lipid-based crude garlic extract (CGE) on the proliferation of human breast, prostate, hepatic, and colon cancer cell lines and mouse macrophageal cells, not previously studied. A coculture experiment of colon cancer cells with mouse macrophageal cells was also designed to help understand the behavior of the CGE in a tumor-like microenvironment, as well as the measures examining the induction of apoptosis and its effects on cell cycle in prostate cancer cells were designed to enhance our understanding of the anticancer effects of CGE.
Materials and Methods
Cell culture line, growth conditions, and reagents
Human liver cancer (Hep-G2), colon cancer (Caco-2), prostate cancer (PC-3), breast cancer (MCF-7), and mouse macrophage cell (TIB-71) lines were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). The human cancer cell lines Hep-G2, Caco-2, PC-3, and MCF-7 were propagated at 37°C in a humidified air/CO2 (95:5 v/v) atmosphere using the ATCC-formulated RPMI-1640 Medium and 10% fetal bovine serum (FBS). The mouse macrophage cell line TIB-71 was propagated at 37°C in a humidified air/CO2 (95:5 v/v) atmosphere using ATCC-formulated DMEM and 10% FBS.
Preparation of CGE
Garlic was purchased from a general supermarket. Briefly, seven garlic cloves were peeled, mixed with 1 mL of sterile water, and crushed in a blender (Cuisinart, Stamford, CT, USA) for 30 sec at room temperature fresh for every experiment. The crushed portion (∼1 g) was then passed through a 10-mL syringe (VWR, West Chester, PA, USA) with a 0.22-μm filter and the supernatant liquid obtained was weighed in a 1.5-mL sterile Eppendorf Tube (VWR) and used directly as the CGE for all experiments. This procedure was based on the method described previously by Boivin et al. 30
In vitro cytotoxicity assay
Hep-G2, Caco-2, MCF-7, PC-3, and TIB-71 cells were plated at a density of 5000 cells/well into 96-well, flat-bottomed tissue culture plates. After an overnight incubation at 37°C in a humidified air/CO2 (95:5 v/v) atmosphere, the cells were treated with 0.125, 0.25, 0.5, or 1 μg/mL of fresh CGE. The cells were then incubated for 72 h. Untreated cells were used as controls. After incubation, the cell number and viability were assessed using the MTS reagent (Promega Corporation, Madison, WI, USA), where 20 μL MTS reagent was added to each well. The cell plates were incubated for an additional 2 h at 37°C and the absorbance read at 490 nm using SpectraMAX plate reader (Molecular Devices, Sunnyvale, CA, USA). Cell inhibition was calculated using the following equation: Cell Inhibition (%)=(1 − (Abss/Absctrl))×100; where Abss is the absorbance of the cells tested with CGE and Absctrl is the absorbance of untreated cells.
Coculture experiments of Caco-2 and TIB-71 cell lines
Caco-2 and TIB-71 cells were plated at a total density of 5000 cells/well into a 96-well, flat-bottomed tissue culture-treated plate. The plating density was chosen such that the ratio of macrophageal cells to cancer cells was 1:4, thus, TIB-71 cells were plated at 1000 cells/well and Caco-2 were plated at 4000 cells/well. Both cell lines were plated using DMEM/10% FBS to maintain consistent experimental conditions from above. The cultures were then incubated overnight at 37°C. The next day the cultures were treated with the same doses of CGE as stated above. The cells were then incubated and examined at 24, 48, and 72 h. Untreated cells were used as controls. After each incubation the cell number and viability were assessed using the MTS reagent (Promega Corporation), where the MTS reagent was added to each well of the plate and the cell plates were incubated for an additional 2 h at 37°C in a humidified air/CO2 (95:5 v/v) atmosphere and then the absorbance was read at 490 nm using a SpectraMAX plate reader (Molecular Devices). Cell inhibition was calculated using the following equation: Cell Inhibition (%)=(1 − (Abss/Absctrl))×100; where Abss is the absorbance of the cells tested with CGE and Absctrl is the absorbance of untreated cells.
Cell cycle analysis
For cell cycle analysis, PC-3 cells were seeded at 2×105 cells/well into a six-well plate and after 24 h, cells were treated with 0.25 or 0.5 μg/mL of CGE for 24 h and the DNA content was determined using propidium iodide (PI) and measured using flow cytometry (BD Biosciences, San Jose, CA, USA). Untreated cells were used as control. The method for cell cycle analysis was previously described 31 ; briefly after treatment, the cells were washed with phosphate-buffered saline (PBS), trypsinized, and fixed in cold ethanol for 2 h. Cells were then washed and resuspended in PBS plus 20 μg/mL of PI (Invitrogen, Carlsbad, CA, USA), plus 100 μg/mL RNase A (Sigma Aldrich, St Louis, MO, USA) for 30 min in the dark at room temperature. Flow Cytometric analysis was performed using the FACScan® cytometer (Becton Dickinson, Franklin Lakes, NJ, USA) and data analyzed using Modfit LT 3.0 software.
Caspase-Glo® 3/7 activity assay
Activation of caspase-3 and −7 plays an important role in apoptosis. 32,33 The Caspase-Glo 3/7 assay kit (Promega) was used to measure the caspase-3/7 activity in PC-3 cells. The caspase-3/7 activity was measured by addition of luminogenic caspase-3/7 substrate, which is cleaved by the enzymes caspase-3/7 to release a substrate for luciferase resulting in the luciferase reaction. Briefly, PC-3 cells (4000 cells/well) were seeded in a white-bottom, tissue culture-treated 96-well plate and incubated for 24 h at 37°C. The PC-3 cells were then treated with 0.25, 0.5, or 1 μg/mL of CGE for 24 h at 37°C. After incubation, 100 μL of caspase-3/7 reagent was added to each well and incubated for 1 h in the dark at room temperature. Luminescence for each well was recorded using Wallac Victor 2 (Perkin Elmer, Boston, MA, USA).
Statistical analysis
SigmaStat software was used for all statistical evaluations (Jandel Scientific, San Rafael, CA, USA). 34 A one-way analysis of variance (ANOVA) or repeated measures ANOVA was used to analyze the data. When statistical significance was found by ANOVA, the Student–Newman–Keuls separation of means was used to determine group differences. All values are expressed as mean±SD of three independent experiments in triplicate. Statistical significance was set at the minimum P<.05.
Results
The growth rate at 72 h for the Hep-G2, PC-3, MCF-7, and TIB-71 cells was significantly inhibited by 80–90% (P<.05) when treated with 0.25, 0.5, or 1 μg/mL of CGE and by 59%, 55%, 48%, and 83% (P<.05), respectively, when treated with 0.125 μg/mL of CGE when compared to untreated cells (Table 1). Caco-2 cells were less responsive to CGE as cell proliferation was significantly inhibited by only 54% (P<.05) when treated with 0.5 and 1 μg/mL, and 44% (P<.05) when treated with 0.25 μg/mL of CGE when compared to untreated cells (Table 1). No statistically significant cell growth inhibition (12%) was observed with 0.125 μg/mL CGE compared to untreated cells after 72 h (Table 1).
Values are mean±SD of three independent experiments performed in triplicate.
Values in a column not sharing a common lowercase superscript letter are significantly different at P<.05.
Values in a row not sharing a common uppercase superscript letter are significantly different at P<.05.
We observed only a 54% inhibitory effect on Caco-2 cell proliferation at the highest dose (1 μg/mL) of CGE (Table 1). To follow up on this observation, we hypothesized that for CGE to be more effective on Caco-2 cells, it may require a tumor-like microenvironment, which could be simulated by coculturing Caco-2 cells with the mouse TIB-71 cell line. This experiment would not only help in improving the activity of CGE on Caco-2 cells but also provide a critical insight into the effectiveness of CGE on tumor reduction. In this coculture experiment, the proportion of Caco-2 cells to TIB-71 cells cultured was four to one. So taking into account their individual growth inhibitory effects 54% for Caco-2 and 90% for TIB-71 at 1 μg/mL after 72 h of treatment, the net growth inhibition obtained after coculture was postulated to be around 60%. At 24 h, the percent of cell proliferation inhibition at 0.25, 0.5, and 1 μg/mL was observed to be ∼60% (P<.05) when compared to untreated cells for all three doses (Fig. 1). However, at 48 h, an increase in percent inhibition of cell proliferation of ∼70% (P<.05) (Fig. 1) was observed with all three doses when compared to untreated cells. A significant increase in cell proliferation inhibition (90%; P<.05) was observed at all three doses, and 51% cell inhibition was observed at 0.125 μg/mL of CGE after 72 h when compared to untreated cells (Fig. 1). Thus, CGE inhibited the cell growth of Caco-2 at 0.25, 0.5, and 1 μg/mL in a time- and dose-dependent manner when cocultured with TIB-71 cells at 24, 48, and 72 h (Fig. 1).
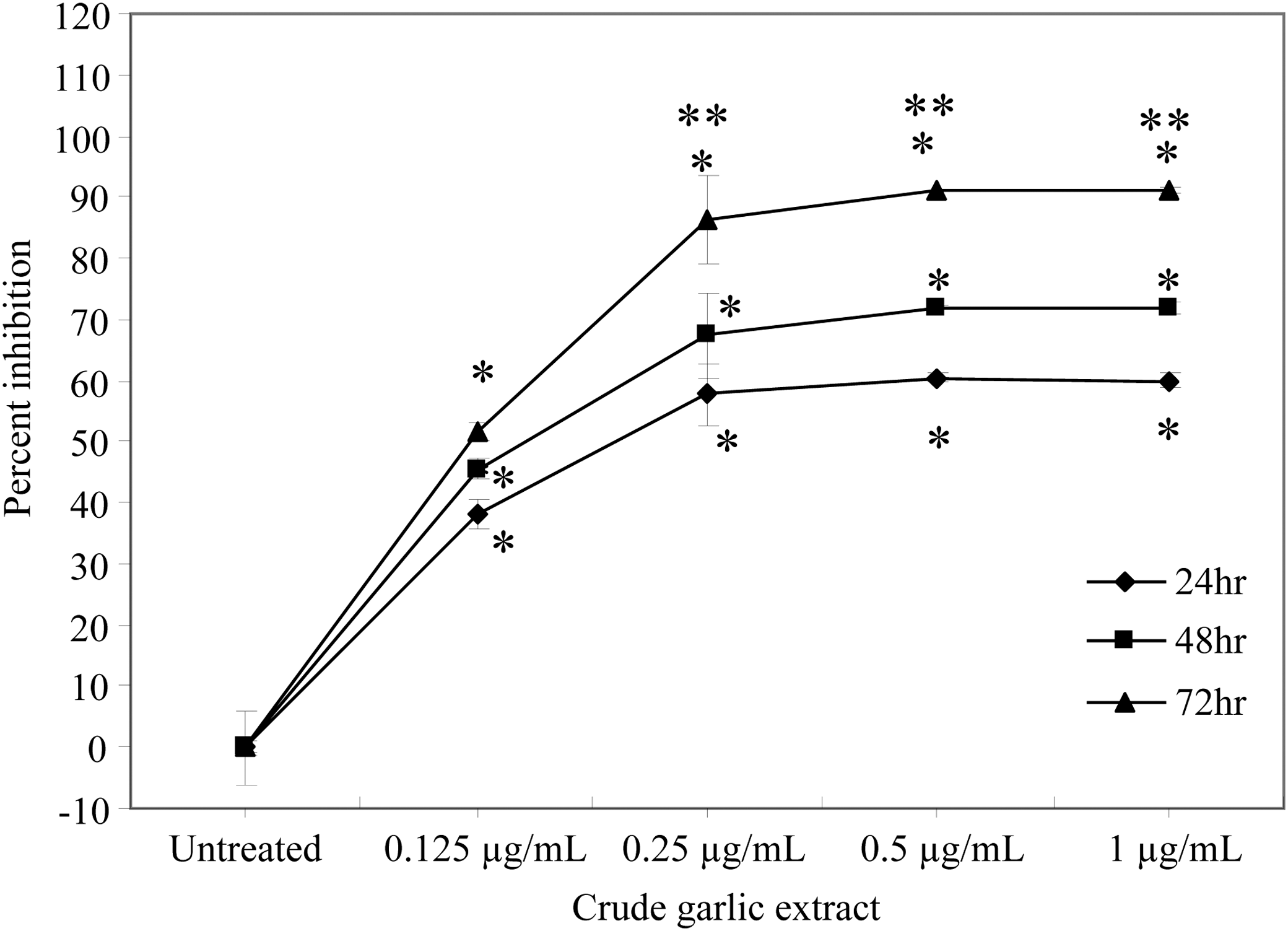
Effect of crude garlic extract (CGE) on cell viability of cocultured Caco-2 and TIB-7 cells. The cocultured Caco-2 and TIB-71 cells were treated with CGE (0.125, 0.25, 0.5, or 1 μg/mL) for 24, 48, and 72 h. Untreated cells were used as control. Cell proliferation inhibition was assessed using MTS assay. All the results are expressed as mean±SD of three independent experiments performed in triplicate. *Statistical significance at P<.05 for CGE treatments versus untreated control at the respective time points of 24, 48, and 72 h. **Statistical significance at P<.05 between 0.25, 0.5, and 1 μg/mL of CGE-treated cells at 24 versus 72 h.
Untreated control cells exhibited 51% of their cells in the G1 phase, whereas the number of cells exhibited in the G1 phase increased to 75% (P<.05) after treatment with 0.5 μg/mL of CGE (Fig. 2). More importantly, we also observed a significant increase (21%; P<.05) in cells treated with 0.5 μg/mL of CGE in the sub-G1 population compared to untreated cells (9%) indicating occurrence of apoptotic events (Fig. 2). A reduced effect (32% in G1 phase) on the cell cycle arrest was observed in the cells treated with 0.25 μg/mL of CGE (Fig. 2).

Percentage of cells in the various phases of the cell cycle of PC-3 cells treated with different doses of CGE. Cell cycle distributions of CGE-treated PC-3 cells were analyzed by flow cytometry. The cells were treated with CGE (0.125, 0.25, 0.5, or 1 μg/mL) or without CGE for 48 h. All the results are expressed as mean±SD of three independent experiments. *Statistical significance at P<.05 when compared to untreated cells.
Compared to untreated control cells, 1 μg/mL of CGE caused a fourfold increase in caspase activity (P<.05) compared to the twofold increase (P<.05) in caspase activity at 0.25 and 0.5 μg/mL (Fig. 3). In addition, 1 μg/mL treated cells had significant caspase activation (P<.05) relative to 0.25 and 0.5 μg/mL treated cells (Fig. 3).
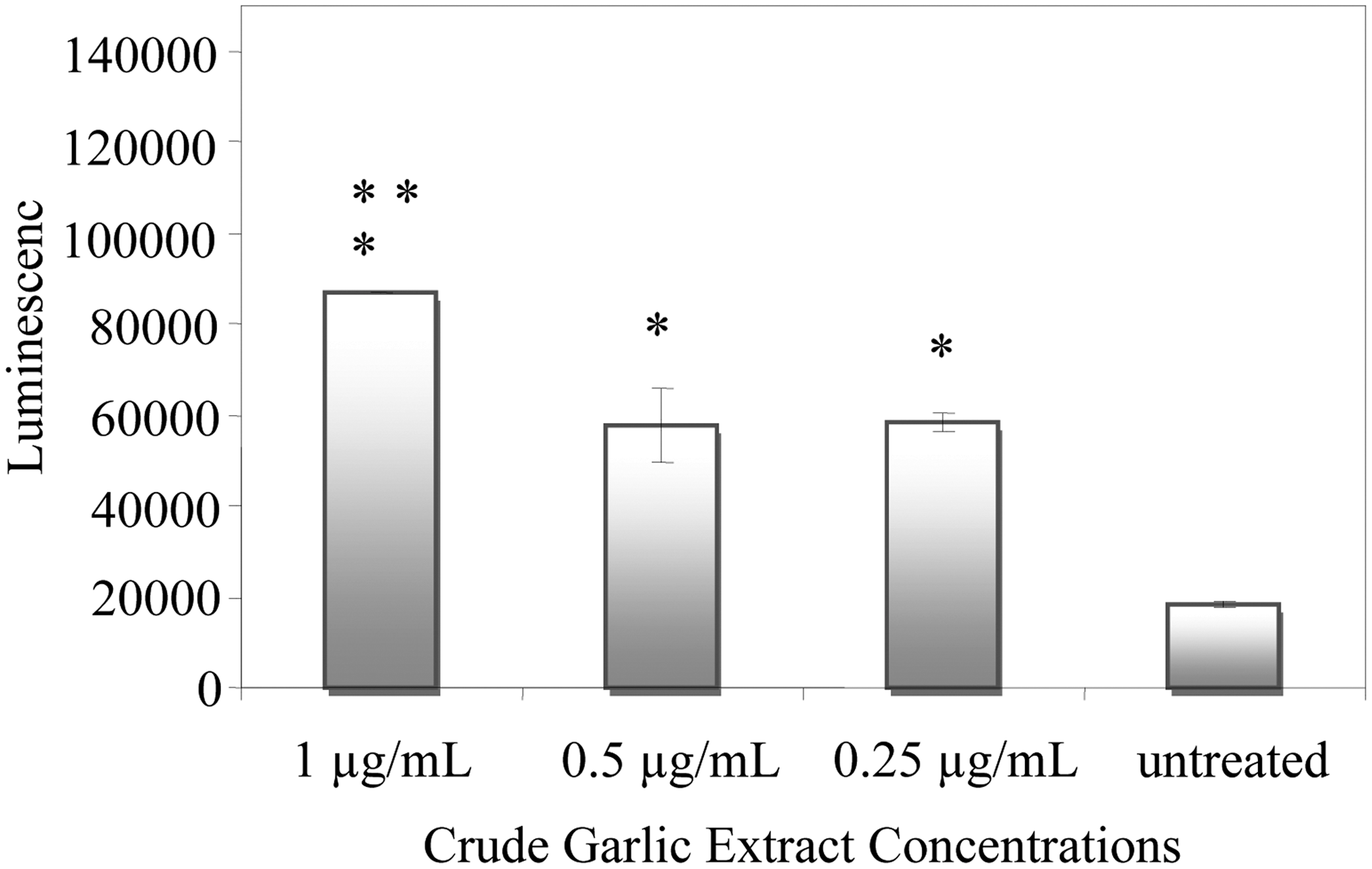
Caspase-3/7 activity in CGE-treated or untreated PC-3 cells after 24 h. Induction of caspase-3/7 was observed when PC-3 cells were treated with 1, 0.5, and 0.25 μg/mL of CGE for 24 h. Caspase activation was assessed using Caspase-Glo assay. *Statistical significance at P<.05 from untreated cells. **Statistical significance at P<.05 when 1 μg/mL CGE-treated cells are compared to 0.5 μg/mL and 0.25 μg/mL CGE-treated cells.
Discussion
Several previous studies have demonstrated that garlic extract exerts anticarcinogenic properties by inhibiting the growth of cancer cells and inducing apoptosis in certain cancer cells in vitro. 35 –37 In addition, epidemiological studies have also shown decreases in cancer incidence with garlic consumption and that high consumption of raw or cooked garlic offers potential protection against cancer development. 9,12,38 –40 Our study used CGE as the potential treatment source of lipid-based chemicals naturally found in garlic to examine its potential anticancer activity at doses that were similar to or slightly lower than previous work 38 –40 on human cancer cell lines: hepatic (Hep-G2), colon (Caco-2), prostate (PC-3), and breast (MCF-7), as well as on a mouse macrophage cell line (TIB-71) not previously published. We observed that as small a dose as 0.25 μg/mL of CGE was capable of inhibiting the proliferation of hepatic, prostate, and breast cancer cells by ∼80–90%. The same dose was also capable of inhibiting the proliferation of the mouse macrophage cell by 90% as well. Higher doses did not improve this anticancer activity. These results are similar to a study carried out by Howard et al., 41 where they observed that garlic-derived OSC showed to be potential antimetastatic agents as they decreased invasion and cell motility of androgen-independent prostate cancer. Another study carried out by Song et al. 24 showed that the diallyl sulfide exhibited cell cycle arrest in colon cancer HCT-116 cells and also induced apoptosis, which was p53 dependent. While these previous studies were carried out with AGE, garlic-derived compounds, or other garlic products such as garlic oil, the use of CGE on other cancer cell lines provides another interesting approach on the anticancer properties of garlic and its various products.
Studies examining the use of garlic on colon cancer cell lines have proven to be very effective with in vitro as well as in vivo studies. 24,42,43 However, in our study, we did not observe as great an anticancer activity with the high dose (1 μg/mL) of CGE on the proliferation of colon cancer cells compared to the other cancer cell lines, although cell proliferation was still inhibited significantly (∼50% compared to control). The reason(s) for this decreased effect on inhibiting cellular proliferation of the Caco-2 colon cancer cell line and other colon cancer and our hepatic, prostate, and breast cancer cell lines is unknown at this time. It is possible that this particular type of colon cancer requires a slightly different environment for its potential growth and metastatic properties. For this reason, we designed a coculture experiment using the colon cancer cell line along with the macrophage cell line to create a microenvironment that might be similar to the environment found in true in vivo colon cancer. We observed that cellular proliferation in this coculture experiment was now inhibited similarly (∼80% with the 0.25 μg/mL dose) to the previous experiments with the other cell lines after the same incubation time period. This finding is novel as this is the first time a coculture experiment of CGE-treated colon cancer cells and macrophageal cells has been carried out and successfully demonstrated that CGE has potent effects on Caco-2 cells when cocultured with tumor-associated macrophages TIB-71, which are part of cellular microenvironment. Thus, CGE may prove to have a chemopreventive action on a stromal cell population as well. This might result in CGE being effective and playing an important role in reduction of tumor size and tumor survival by not only reducing the tumor cell growth but also effectively inhibiting the infiltrating macrophageal cell population in a tumor-like microenvironment. Interestingly, the cell proliferation inhibition obtained after coculturing the cells was 60% after 24 h. Subsequently, after 48 h, the cell proliferation inhibition increased to 70% and most importantly, after 72 h, we achieved 90% cell proliferation, thus demonstrating that CGE had a time-dependent inhibitory effect on Caco-2 cells after coculturing with TIB-71 cells. We must emphasize that, however, this experiment was carried out in vitro. Further in vivo investigations on the exact mechanisms of inhibition are warranted.
Inhibitory effects or induction of apoptosis are highly correlated to arrest cell cycle in the G1, S, G2, and M phase. 44,45 The antiproliferative effects of garlic on various cells have been linked to a block in the progression of cell cycle from the G1 phase to G2 phase or from G2 to M phase. 46,47 Our data specifically on cell cycle analysis of PC-3 cells treated with 0.5 μg/mL of CGE indicated a G1 arrest, with significantly greater numbers of cells in the G1 phase compared to untreated cells, along with an increase cell numbers in the sub-G1 population, thus suggesting a possible mechanism of apoptosis with CGE. 48,49 This effect was reduced when the cells were treated with 0.25 μg/mL of CGE indicating a dose-dependent response in the cell cycle arrest. Thus, it may be concluded that the cell cycle arrest in the G1 phase indicates that CGE has the ability to induce apoptosis in PC-3 cells.
One of the key mechanisms of targeted cancer therapy includes induction of apoptosis. Apoptosis is a form of programmed cell death that is characterized by a variety of morphological features, including loss of membrane symmetry, chromatin condensation, and DNA fragmentation. Caspase-3 activation is essential for the majority of apoptotic events in cells. 32,33 The activation of caspase-3 leads to apoptosis by inducing poly(ADP-ribose) polymerase (PARP) cleavage and breakage of chromosomal DNA. 33 Our study showed that CGE caused a greater response in caspase activity compared to untreated cells and that the best response was associated with the highest dose (1 μg/mL) used. These results show that the treatment of PC-3 cells with 1 μg/mL of CGE increases caspase-3/7 activation suggesting that CGE activation of caspase-3/7 activity might play a critical role in cell apoptosis in the present study. Thus, this observation for PC-3 cells along with the cell proliferation inhibition and cell cycle arrest by CGE could provide an effective strategy to control carcinogenesis.
These observations suggest that CGE possesses the characteristics of an anticancer agent and that consumption of garlic may offer chemopreventive effects against cancers. Thus, the activity of CGE against proliferation of multiple cancer cell lines clearly demonstrates that CGE has the potential of being effective not just against specific cancer types but rather be a universally applicable anticancer agent that is effective against multiple cancers. Furthermore, CGE can also be explored as a viable option for nutraceutical preparation of garlic that could be applied toward use of complementary medication with the currently available chemotherapies for cancer treatment with additional investigation. Although CGE contains potent various chemicals that may inhibit the cell growth of several cancer cell lines in vitro, the use of whole garlic or its extracts may provide a more synergistic effect compared to the individual chemical active components that have been previously studied. Furthermore, it was also observed that one of the possible mechanisms of the antiproliferative activity of CGE in prostate cancer cells could be related to activation of caspase-3/7 and cell cycle arrest, possibly leading to greater apoptosis. Future in vivo and in vitro studies will be needed to study the mechanisms and effects of CGE on cancer progression as well as identifying which of the chemical components of CGE could be a potentially safe and effective anticancer agent.
Footnotes
Acknowledgments
M.B. and S.K. designed the study and performed the laboratory analyses. All authors contributed in the interpretation and writing of the final article. The authors would like to thank Swetha Kambalapally, Dr. Robert J. Nicolosi, and Maureen Faul for their technical and administrative support with this study and article.
Author Disclosure Statement
No conflicts of interest exist for any of the authors. Internal funding was used by the corresponding author for all experiments.