
Abstract
Impaired fasting blood glucose is one of the landmark signs of metabolic syndrome, together with hyperinsulinemia, dyslipidemia, hypertension, and a chronic proinflammatory, pro-oxidative, and prothrombotic environment. This study investigates the effect of wild blueberry (WB) consumption on blood glucose levels and other parameters involved in glucose metabolism in the obese Zucker rat (OZR), an experimental model of metabolic syndrome. Sixteen OZRs and 16 lean littermate controls (lean Zucker rat [LZR]) were fed an 8% enriched WB diet or a control (C) diet for 8 weeks. Plasma concentrations of glucose, insulin, glycated hemoglobin GHbA1c, resistin, and retinol-binding protein 4 (RBP4) were measured. Expression of the resistin, RBP4, and glucose transporter GLUT4 genes was also determined both in the liver and the abdominal adipose tissue (AAT). Plasma glycated hemoglobin HbA1c, RBP4, and resistin concentrations were significantly lower in OZRs following the WB diet (−20%, −22%, and −27%, respectively, compared to C diet, P<.05). Following WB consumption, resistin expression was significantly downregulated in the liver of both OZRs and LZRs (−28% and −61%, respectively, P<.05), while RBP4 expression was significantly downregulated in the AAT of both OZRs and LZRs (−87% and −43%, respectively, P<.05). All other markers were not significantly affected following WB consumption. In conclusion, WB consumption normalizes some markers related to glucose metabolism in the OZR model of metabolic syndrome, but has no effect on fasting blood glucose or insulin concentrations.
Introduction
T
Impaired fasting blood glucose is one of the landmark signs of the metabolic syndrome, together with hyperinsulinemia, dyslipidemia, hypertension, and a chronic proinflammatory, pro-oxidative, and prothrombotic environment. 2
Increasing evidence indicates that diet and dietary bioactive compounds can play a fundamental role in preventing and reversing most risk factors associated with the metabolic syndrome, 3 suggesting diet as a possible alternative to pharmacotherapy.
Wild blueberries (WB) are one of the richest fruit sources of anthocyanins (ACN), 4 a class of phenolic bioactive compounds that have been shown to have insulin-sensitizing effects and improve glucose utilization. 5
The obese Zucker rat (OZR) is an excellent experimental model to study the metabolic syndrome. 6 As a result of a spontaneous mutation, it develops obesity, insulin resistance, hyperglycemia, hyperinsulinemia, hypertriglyceridemia, and hypercholesterolemia within the first weeks of its life and without the need for feeding it high-fat diets. 6
In previous studies conducted in our laboratory, we have used the OZR model to study the effect of WB consumption on several parameters of the metabolic syndrome. 7 –9 Eight-week consumption of a diet containing 8% WB powder was able to decrease markers of inflammation, 7 improve blood lipid profile, 8 affect the expression of genes involved in the inflammatory response and in lipid metabolism, 7,8 and modulate endothelial function. 9
In this study, we use the same experimental approach to test the effect of WB consumption on glucose metabolism, another key component of the metabolic syndrome. In particular, we measure plasma fasting concentrations of glucose, insulin, glycated hemoglobin, resistin, and retinol-binding protein 4 (RBP4). We also measure the expression of the resistin, RBP4, and glucose transporter GLUT4 genes in the liver and the abdominal adipose tissue (AAT).
Materials and Methods
Zucker rats
Sixteen male OZRs and 16 male lean littermate controls (lean Zucker rat [LZR]) were purchased from Charles River Laboratories at 8 weeks of age. Rats from each group were randomly assigned to a WB-enriched diet or a control (C) diet for 8 weeks. Rats were housed in mesh-bottomed cages, in a room maintained at 22°C with a light cycle of 12-h light–12-h dark. Food and tap water were provided ad libitum. Food intake was measured daily, and rats were weighed every week. The experimental protocol was reviewed and approved by the University of Maine Institutional Animal Care and Use Committee (IACUC) and complied with the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments.
Wild blueberries
WB were provided as a composite from Wyman's, and subsequently freeze-dried, powdered, and vacuum packed by FutureCeuticals. The WB powder was stored in the dark at −20°C until use. Analysis of the WB powder has been previously reported. 10 Total ACN content was 1.5% w/w, with malvidin 3-galactoside and peonidin 3-glucoside as the most abundant ACN.
Diets
The control diet was prepared mixing 691 g/kg dextrose, 200 g/kg egg white solids, 60 g/kg corn oil, 35 g/kg mineral mix (AIN-93M), 10 g/kg vitamin mix (AIN-93), 4 g/kg D-L-methionine, and 2 mg/kg biotin. It provided ∼71% carbohydrate, 6% fat, 17% protein, and 403 kcal/100 g. The WB diet was prepared in the same way, but incorporated 80 g/kg (8%) of WB powder substituting for an equivalent amount of dextrose. It provided ∼68% carbohydrate, 6% fat, 17% protein, 1.5% fiber, 0.12% ACN, and 393 kcal/100 g. Diets were stored at 4°C and used within 1 week of preparation. Using the body surface area calculation, 11 the amount of WB provided to rats corresponds to approximately two daily cups of fresh WB for an average human adult.
Sample collection
At the end of the experimental period, animals were fasted overnight, anesthetized with 95% CO2/5% O2 for 90 sec, and exsanguinated by intracardiac puncture. Blood was collected with 200 μL of a 5% EDTA solution and centrifuged at 1500 g for 15 min at 4°C. Plasma was collected and stored at −80°C until analysis. Liver and AAT samples were immediately excised, snap frozen in liquid nitrogen, and stored at −80°C for subsequent mRNA extraction.
Plasma biochemical markers
The plasma glucose concentration was measured using a colorimetric method with the Glucose Colorimetric Assay Kit (#10009582; Cayman). The plasma insulin concentration was determined using the Rat/Mouse Insulin ELISA Kit (#EZRMI-13K; Millipore). The plasma concentration of glycated hemoglobin GHbA1c was measured using the Rat GHbA1c ELISA Kit (#MBS883244; MyBioSource).
Insulin resistance
The homeostasis model assessment (HOMA), an index of insulin resistance, was calculated as glucose (mM)×insulin (μU/mL)/22.5. 12
Gene expression
Messenger RNA from frozen liver and AAT fragments was isolated using the RNeasy Mini Kit (Qiagen) and the RNeasy Lipid Tissue Mini Kit (Qiagen) following the instructions provided by the manufacturer. Purity and concentration of RNA were estimated spectrophotometrically by measuring absorbance at 260 and 280 nm in UV transparent cuvettes. cDNA was synthesized from mRNA using the QuantiTect Reverse Transcription kit (Qiagen) and used as a template for a two-step, real-time reverse transcription polymerase chain reaction (RT-PCR) amplification on a quantitative PCR System (CFX96; Bio-Rad) using SYBR Green master mix (SSoFast EvaGreen, #172-5202; Bio Rad) and the following rat-specific primers: Slc2a4 (RefSeq NM_012751, #QT00175931; Qiagen) to target GLUT4; Retn (RefSeq NM_144741, #QT00186886; Qiagen) to target resistin; and Rbp4 (RefSeq NM_013162, XM_001080229, XM_215285, #QT00428680; Qiagen) to target RBP4. For each primer and tissue, the analysis was performed in triplicate with a reaction volume of 20 μL per well (1.5 μL reverse transcription product, 10 μL SYBR Green Mix, 2 μL primers forward+reverse, and 6.5 μL RNAse-free water). After an enzyme activation step (95°C for 30 sec), 45 amplification cycles were performed (denaturation at 95°C for 2 sec, annealing/extension at 60°C for 5 sec), followed by a melting curve (75–95°C in 0.5°C increases, 2 sec per step) to verify amplification specificity. Relative expression of the genes of interest was determined by the ΔΔCt method 13 relative to the housekeeping gene beta-actin (Actb, RefSeq NM_031144, #QT00193473; Qiagen) and expressed as fold variation compared to the LZR on the control diet.
Statistical analysis
Data were analyzed using two-way analysis of variance with diet (WB vs. C diets) and animal models (OZR and LZR) as independent factors. Significant main effects (diet and model) as well as the interactions diet x model were further evaluated using the Tukey honestly significant different post hoc test. Statistical analysis was performed using R statistical software version 2.15.1 (R Foundation for Statistical Computing). Results are expressed as mean±SEM and considered significant at P<.05.
Results
No statistically significant differences were found for average body weight, average weight gain, and average food intake during the experimental period between rats in the WB and the control group (data not shown).
A significant effect of the animal model was observed for fasting plasma glucose, insulin, glycated hemoglobin HbA1c, RBP4, and resistin concentrations, which were all significantly higher in the OZR group compared with the LZR group (+57%, +471%, +55%, +93%, and +34%, respectively), as reported in Table 1. No statistically significant interaction with WB consumption was observed for glucose or insulin concentrations either in the OZR group or the LZR group. In contrast, glycated hemoglobin HbA1c concentrations were significantly lower in the OZRs on the WB diet compared with those on the control diet (−20%). Calculation of the HOMA score for insulin resistance revealed a significantly higher insulin resistance in OZRs compared with LZRs, but no effect of WB consumption was observed.
Values are mean±SEM, n=8.
Different superscript letters indicate statistically significant differences (P<.05).
C, control; HOMA-IR, homeostasis model assessment scores-for insulin resistance; LZR, lean Zucker rat; OZR, obese Zucker rat; RBP4, retinol-binding protein 4; WB, wild blueberry.
Both plasma RBP4 and resistin concentrations were significantly lower in the OZRs following the WB diet compared with those on the control diet (−22% and −27%, respectively).
Liver expression of the GLUT4 gene was uniform in LZRs and OZRs, with no significant effect of diet (Fig. 1). In the AAT, expression of GLUT4 was significantly lower in the OZR group compared with the LZR group, with no significant effect of diet (Fig. 1).

Relative expression of GLUT4 in the liver and abdominal adipose tissue (AAT) of the lean Zucker rat (LZR) and obese Zucker rat (OZR) following a control (C) or a wild blueberry (WB) diet. Values are mean±SEM, expressed as 2−ΔΔCt normalized to beta-actin and relative to LZR-C. n=8. a,bDifferent superscript letters indicate statistically significant differences (P<.05).
A significant effect of the animal model was observed for expression of the resistin gene both in the liver and the AAT (Fig. 2). In particular, resistin expression was significantly higher in the liver and significantly lower in the AAT of OZRs compared with LZRs. Furthermore, with WB consumption, resistin expression was significantly downregulated in the liver of both OZRs and LZRs, while no effect of diet was observed in the AAT (Fig. 2).
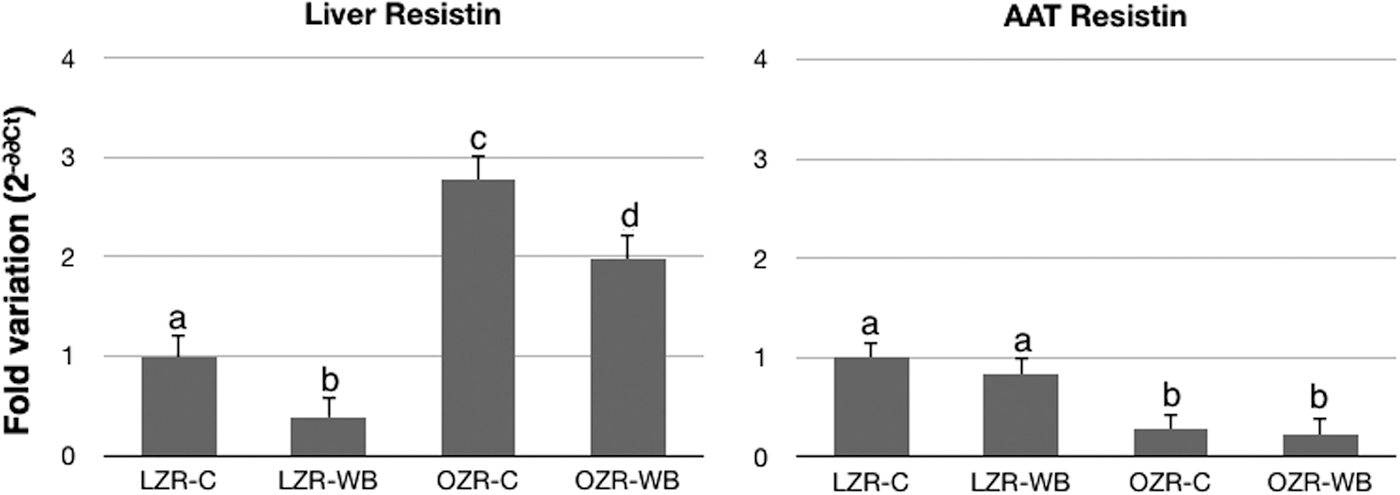
Relative expression of resistin in the liver and AAT of LZRs and OZRs following a control (C) or a WB diet. Values are mean±SEM, expressed as 2−ΔΔCt normalized to beta-actin and relative to LZR-C. n=8. a,b,c,dDifferent superscript letters indicate statistically significant differences (P<.05)
In the liver, expression of the RBP4 gene was significantly lower in the OZR group compared with the LZR group, with no effect of the diet. In the AAT, no significant effect of the animal model was noted, but a significant effect of the WB treatment was observed both in the LZR group and in the OZR group (Fig. 3). In particular, AAT expression of RBP4 was significantly downregulated both in OZRs and LZRs following WB consumption.
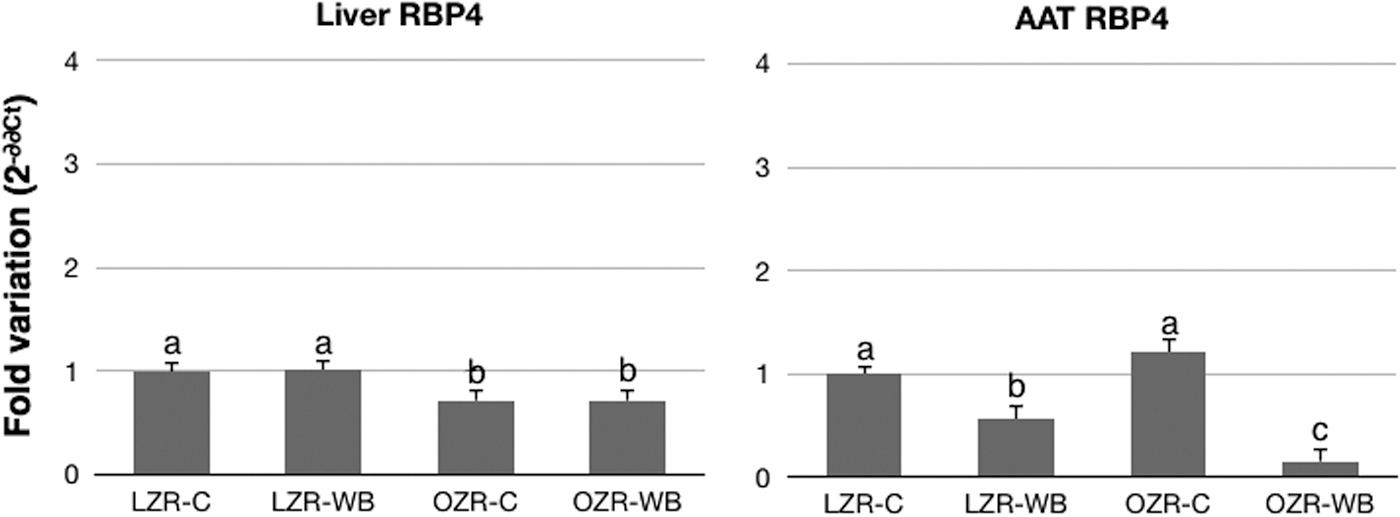
Relative expression of retinol-binding protein 4 (RBP4) in the liver and AAT of LZRs and OZRs following a control (C) or a WB diet. Values are mean±SEM, expressed as 2−ΔΔCt normalized to beta-actin and relative to LZR-C. n=8. a,b,cDifferent superscript letters indicate statistically significant differences (P<.05)
Discussion
This study investigated the effects of 8-week WB consumption on markers of glucose metabolism in the OZR, a model of the metabolic syndrome.
Compared with their lean littermate controls (LZR), OZRs had significantly higher circulating concentrations of fasting plasma glucose, fasting insulin, and glycated hemoglobin HbA1c, and a higher insulin resistance HOMA score independent of WB consumption.
Concentrations of glycated hemoglobin HbA1c were significantly lower in OZRs in the WB diet group compared with OZRs in the control diet group. A raised glycated hemoglobin HbA1c concentration is a marker of long-term hyperglycemia linked with increased cardiovascular mortality, as recently reviewed by Piya et al. 14
Previously, DeFuria et al. 15 have shown that diet supplementation with 4% blueberry powder for 8 weeks reduced fasting glucose levels and attenuated insulin resistance in high-fat-fed mice. In our study, fasting plasma glucose and insulin levels were not significantly different between rats on the control and the WB diet, in spite of the HbA1c being lower following WB consumption. This apparent contradiction may be due to a circadian effect (differences in blood glucose levels may not occur in a fasting state, but in the postprandial state) or due to a temporal effect (the differences between diet groups in blood glucose and insulin concentrations may occur in the earlier weeks of WB treatment, but be abolished by the time the eighth week is reached).
Recent human intervention studies have investigated some effects of medium-term consumption of dietary achievable doses of blueberries on glucose metabolism. Basu et al. 16 have reported that in obese individuals consuming a freeze-dried blueberry drink (containing the equivalent of 350 g fresh blueberries) for 8 weeks, while improving blood pressure, it did not affect blood glucose concentrations. Riso et al. 17 reported no change in fasting blood glucose levels in individuals with at least one risk factor for cardiovascular disease following 6-week consumption of a freeze-dried WB drink providing 375 mg of ACN (equivalent to 148 g of fresh blueberries). Stull et al. 18 observed an improvement in insulin sensitivity in a group of obese insulin-resistant individuals consuming two freeze-dried blueberry drinks (containing the equivalent of 300 g fresh blueberries each) daily for 6 weeks.
The present study also measured the circulating levels and genetic expression in the liver and AAT of proteins involved in glucose metabolism and the following links between insulin resistance and inflammation: resistin, RBP4, and the glucose transporter GLUT4.
Resistin is a hormone whose elevations are associated with inflammation and dyslipidemia and are believed to lead to insulin resistance. 19 In the liver, resistin increases glucose production and release in the bloodstream, while in peripheral tissues, it interferes with the normal insulin signaling cascade, thus decreasing insulin sensitivity. 19 –21 In the present study, OZRs had significantly higher plasma circulating levels of resistin and significantly higher resistin expression in the liver compared with LZRs. WB diet significantly reduced plasma resistin in OZRs and downregulated liver resistin expression both in OZRs and LZRs. Previously, green tea polyphenols have been shown to inhibit resistin expression in isolated mouse adipocytes. 22 Studies on the effect of ACN on resistin are rather limited. Ten-week consumption of ACN-rich grape-bilberry juice resulted in improved lipid profile, but did not affect serum glucose or insulin in Fischer rats. 23 However, serum resistin significantly decreased. 23 In another study, male C57BL/6J mice fed a high-fat diet experienced increased serum resistin concentrations; however, this effect was not reversed by either purified black raspberry ACN or freeze-dried whole black raspberry juice for 10 weeks. 24 Thus, the associations between resistin levels and circulating levels of glucose or insulin have given mixed effects, as recently reviewed by Barnes and Miner. 20
RBP4 is the main vitamin A transport protein in the bloodstream, but it has also been recognized as one of the adipokines, which may explain the long known association between obesity and insulin resistance. 25,26 Increased plasma circulating levels of RBP4 have been associated in mice with impaired glucose uptake into the skeletal muscle and increased gluconeogenesis in the liver, while decreased RBP4 levels have been shown to improve insulin sensitivity and glucose tolerance. 25 In our study, RBP4 plasma concentrations were significantly higher in OZRs compared with LZRs. Following the WB diet, RBP4 circulating concentrations significantly decreased both in OZRs and in LZRs, compared with their respective controls. This effect mirrors the expression of the RBP4, which was measured in the AAT; RBP4 expression was significantly downregulated both in OZRs and LZRs following the WB diet. Conversely, no significant differences in RBP4 expression with WB consumption were observed in the liver. A similar pattern was observed by Sasaki et al. 27 in KK-Ay type 2 diabetic mice fed a 0.2% cyaniding-3-glucoside supplemented diet for 5 weeks; RBP4 expression was not affected in the liver, but it was downregulated in the AAT, while GLUT4 expression was upregulated, suggesting an antidiabetic effect that was confirmed by improved insulin sensitivity and reduced blood glucose levels. Thus, RBP4 appears to respond differently in different tissues. The reason why RBP4 expression in the liver tends to be more constant may be related to its primary role as a vitamin A transporter, whereas expression of RBP4 in the adipose tissue may be more dependent on the redox and inflammatory status. In mice, PPAR-γ agonist rosiglitazone, an insulin sensitizing medication, results in decreased serum RBP4 and downregulation of RBP4 expression in the adipose tissue, but not the liver. 28 It is very likely that the observed effect of WB consumption on RBP4 levels and expression is controlled by the transcription factor PPAR-γ. Indeed, in a previous study, 8 we have documented that 8% WB consumption upregulates PPAR-γ expression in the AAT, but not the liver, of both lean and OZRs. We also have documented a general anti-inflammatory effect of WB consumption in OZRs, as evidenced by decreased expression of nuclear factor kB both in the liver and AAT and decreased levels of proinflammatory cytokines, such as IL-6 and TNF-α. 7 Chronic low-grade inflammation and, in particular, the increased secretion of proinflammatory molecules by dysfunctional macrophage-rich adipocytes is likely the link between obesity and insulin resistance. 2
The insulin-sensitive glucose transporter GLUT4 is the rate-limiting step of glucose uptake into skeletal muscle cells and adipocytes. 29 Insulin resistance is associated with downregulation of GLUT4 in adipocytes, 25 resulting in impaired glucose uptake. This, in turn, promotes adipocyte secretion of RBP4 to stimulate gluconeogenesis in the liver, while liver RBP4 does not appear to be affected. In type 2 diabetic mice fed a bilberry extract (10 g/kg diet of ACN) for 5 weeks, GLUT4 expression was upregulated in white adipose tissue and skeletal muscle, while insulin sensitivity was markedly improved and blood glucose levels decreased. 30 GLUT4 expression was also upregulated in diabetic mice fed cyanidin-3-glucoside extract (0.2%) for 5 weeks. 27 In the present study, GLUT4 expression was unaffected by WB consumption in OZRs and LZRs. It is to be noted, however, that another important regulatory step of GLUT4 is its translocation to the cell membrane, 29 which was not evaluated in this study.
In conclusion, 8-week WB consumption significantly decreased plasma glycated hemoglobin, resistin, and RBP4 concentrations in the OZR model of metabolic syndrome and downregulated liver resistin expression and RBP4 expression in the AAT. Such changes can all be considered beneficial, suggesting that regular WB consumption could help normalize the dysregulated glucose metabolism associated with the metabolic syndrome. However, while WB consumption appears to affect some markers of glucose metabolism, further research is needed to clarify the extent of such response and pinpoint its exact effects on blood glucose and insulin concentrations.
Footnotes
Acknowledgments
The present study was supported by the Wild Blueberry Association of North America (WBANA). This work is based upon research supported in part by the Hatch Grant no. ME0-8553-09 from the United States Department of Agriculture (USDA), National Institute of Food and Agriculture and the Maine Agricultural and Forest Experiment Station publication No. 3355.
Author Disclosure Statement
No competing financial interests exist.