
Abstract
We previously demonstrated that Nostoc commune var. sphaeroids Kützing (NO), a blue-green alga (BGA), exerts a hypolipidemic effect in vivo and its lipid extract regulates the expression of genes involved in cholesterol and lipid metabolism in vitro. The objective of this study was to investigate whether the hypolipidemic effect of NO is attributed to an algal lipid or a delipidated fraction in vivo compared with Spirulina platensis (SP). Male C57BL/6J mice were fed an AIN-93M diet containing 2.5% or 5% of BGA (w/w) or a lipid extract equivalent to 5% of BGA for 4 weeks to measure plasma and liver lipids, hepatic gene expression, intestinal cholesterol absorption, and fecal sterol excretion. Plasma total cholesterol (TC) was significantly lower in 2.5% and 5% NO-fed groups, while plasma triglyceride (TG) levels were decreased in the 5% NO group compared with controls. However, neither NO organic extract (NOE) nor SP-fed groups altered plasma lipids. Hepatic mRNA levels of sterol regulatory element-binding protein 2, 3-hydroxy-3-methyl-glutaryl-CoA reductase (HMGR), carnitine palmitoyltransferase-1α, and acyl-CoA oxidase 1 were induced in 5% NO-fed mice, while there were no significant changes in hepatic lipogenic gene expression between groups. NO, but not NOE and SP groups, significantly decreased intestinal cholesterol absorption. When HepG2 cells and primary mouse hepatocytes were incubated with NOE and SP organic extract (SPE), there were marked decreases in protein levels of HMGR, low-density lipoprotein receptor, and fatty acid synthase. In conclusion, the nonlipid fraction of NO exerts TC and TG-lowering effects primarily by inhibiting intestinal cholesterol absorption and by increasing hepatic fatty acid oxidation, respectively.
Introduction
D
Blue-green algae (BGA) are photosynthetic microorganisms found in the aquatic ecosystem and have been consumed by humans as food and medicine for centuries. 11 Commonly consumed BGA species include Spirulina platensis (SP), Spirulina maxima, Aphanizomenon flos-aquae, and Nostoc commune var. sphaeroids Kützing (NO), of which SP is the most commonly commercialized and consumed species. 11 –15 BGA have been recognized as a natural product with significant pharmaceutical potential, such as anticancer, antibacterial, antiviral, antiallergic, antioxidant, anti-inflammation, and enzyme inhibiting activities. 16 –18
We previously reported that 5% NO supplementation significantly lowered plasma TC and TG levels in C57BL/6J mice with a concomitant increase in the expression of hepatic 3-hydroxy-3-methyl-glutaryl-CoA reductase (HMGR), the rate-limiting enzyme for cholesterol biosynthesis. 15 However, lipid extract of NO reduced the expression of sterol regulatory element-binding protein 2 (SREBP-2) and HMGR in HepG2 cells. 15 Therefore, the objectives of the present study were to determine if lipid extract or nonlipid fraction of NO is responsible for the lipid-lowering effect of NO and to gain mechanistic insight in vivo and in vitro compared with SP supplementation.
Materials and Methods
Animal care and diet
C57BL/6J male mice, at the age of 8 weeks, were purchased from Jackson Laboratory (Bar Harbor, ME, USA) and randomly assigned into one of seven groups. Mice were fed a modified AIN-93M control diet or diet supplemented with 2.5% or 5% of NO or SP (w/w), NO organic extract (NOE), or SP organic extract (SPE) ad libitum. NO (AlgaBerry™) and SP powder (Earthrise® Natural Spirulina) were kindly provided by Algaen Corporation (Winston-Salem, NC, USA) and Earthrise Nutritionals (Irvine, CA, USA), respectively. NOE and SPE contained the same amount of lipid extract present in 5% of NO or SP, respectively. Large-scale lipid extraction was conducted as previously described. 19 The algal extract was dissolved in soybean oil before being incorporated into the diet. The experimental diets were prepared by Dyets, Inc., (Bethlehem, PA, USA) according to our specifications and diet composition (Table 1). Mice (n=10 for all groups except n=9 for the SP 5% group) were housed in a controlled environment with 12-h light/12-h dark cycles. Body weight and food consumption were recorded weekly. After 4 weeks on the experimental diets, mice were fasted for 4 h and anesthetized by isoflurane inhalation. Blood samples were collected into a 2-mL BD vacutainer containing EDTA by cardiac puncture and centrifuged at 1500 g for 10 min at 4°C to remove red blood cells. Tissue samples were snap-frozen in liquid nitrogen and stored at −80°C until use. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Connecticut.
Solka-Floc cellulose.
AIN-93 mineral mix.
AIN-93 vitamin mix.
BGA, blue-green algae; NOE, NO organic extract; SPE, SP organic extract.
Plasma and liver lipid analysis
Lipid from liver samples was extracted into chloroform:methanol (2:1) and solubilized in Triton X-100 as previously described. 15 Plasma and liver TC levels were enzymatically determined using cholesterol reagents from Pointe Scientific (Canton, MI, USA), and TG levels were quantified by the L-type triglyceride M kit from Wako Chemical USA (Richmond, VA, USA). The liver free cholesterol (FC) level was measured by Free Cholesterol E reagent (Wako Chemicals).
Cholesterol absorption efficiency and fecal sterol analysis
Fractional intestinal cholesterol absorption was measured using the dual isotope method as we previously described. 15 Fecal neutral and plant sterols were determined by gas chromatography, and acidic sterols were determined by enzymatic analysis using the Wako Bile Acid Kit (Wako Chemicals) as described. 15
Gene expression analysis by quantitative real-time PCR
Total RNA was isolated from tissue samples using TRIzol reagent (Invitrogen, Grand Island, NY, USA), and quantitative real-time PCR (qRT-PCR) analysis for hepatic gene expression was conducted as previously described using the SYBR Green procedure and CFX96 real-time PCR detection system (BioRad, Hercules, CA, USA). 20,21 Primer sequences were designed according to GenBank database using the Beacon Designer software (Premier Biosoft, Palo Alto, CA, USA), and primer sequences are listed in Table 2.
SREBP-2, sterol regulatory element-binding protein 2; HMGR, 3-hydroxy-3-methyl-glutaryl-CoA reductase; LDLR, low-density lipoprotein receptor; SREBP-1c, sterol regulatory element-binding protein 1c; FAS, fatty acid synthase; ACC-1, acetyl-coA carboxylase 1; SCD-1, stearoyl CoA desaturase 1; CPT-1α, carnitine palmitoyltransferase 1α; ACOX-1, acyl-CoA oxidase 1; qRT-PCR, quantitative real-time PCR.
HepG2 cell culture and treatment
HepG2 cells (ATCC, Manasas, VA, USA) were incubated in low-glucose DMEM containing 10% fetal bovine serum (FBS), penicillin/streptomycin (P/S, penicillin 100 U/mL, streptomycin 100 μg/mL), 1×vitamins, and 2 mM
Primary hepatocyte isolation and culture
Primary hepatocytes were isolated from C57BL/6J mice on a regular chow diet. Mice were anesthetized by ketamine HCl/xylazine (120/6 mpk) (Henry Schein, Melville, NY, USA), and the abdominal cavity was opened to expose the portal vein. Subsequently, the ribcage was cut and a 20 g×1" catheter (Terumo, New Jersey, NJ, USA) was inserted into the inferior vena cava through the right atrium, after which the portal vein was cut. The liver was perfused with Hank's balanced salt solution (HBSS) (Gibco, Grand Island, NY, USA) containing 50 mM ethylene glycol tetraacetic acid, 1 M glucose, and P/S at a speed of 6–7 mL/min for 3 min, and then it was perfused with HBSS containing 1 M CaCl2, 1 M glucose, P/S, and 0.425 mg/mL of type I collagenase (Worthington, Lakewood, NJ, USA) at a rate of 6–7 mL/min for 6 min to digest tissue. The digested liver was then carefully removed and resuspended in prewarmed Williams' Media E (Sigma-Aldrich, St. Louis, MO, USA). Subsequently, the cells were filtered through a 100-μm nylon mesh strainer (Fisher, Waltham, MA, USA) and centrifuged at 50 g for 2 min thrice to remove nonparenchymal cells. Alive hepatocytes were then collected by laying the cells in Williams' Media E over 40% Percoll solution (Sigma-Aldrich) and subsequent centrifuging at 1100 g for 5 min with lowest acceleration and deceleration settings. Hepatocytes were counted and plated in Williams' E Media (Life Technologies, Carlsbad, CA, USA) containing 10% FBS, P/S, and 2 mM
Statistical analysis
One-way ANOVA and Newman–Keuls pair-wise comparison were performed to compare a mean difference between groups using GraphPad InStat 5 (GraphPad Software, La Jolla, CA, USA). An α-level of P<.05 was considered statistically significant and all data are expressed as mean±SEM.
Results
Effect of NO and SP on plasma TC and TG
To determine hypolipidemic effects of BGA and their lipid extracts, mice were fed a diet supplemented with BGA at 2.5% and 5% levels or their extract that contains lipid extract equivalent to 5% whole algae. Mice fed a diet containing 2.5% or 5% NO showed significantly lower plasma TC concentrations, while 5% NO supplementation significantly decreased plasma TG levels compared with the other groups (Fig. 1). However, NOE supplementation did not significantly lower either plasma TC or TG levels compared with the control group. There were no significant changes in plasma TC and TG concentrations by any SP supplementations.
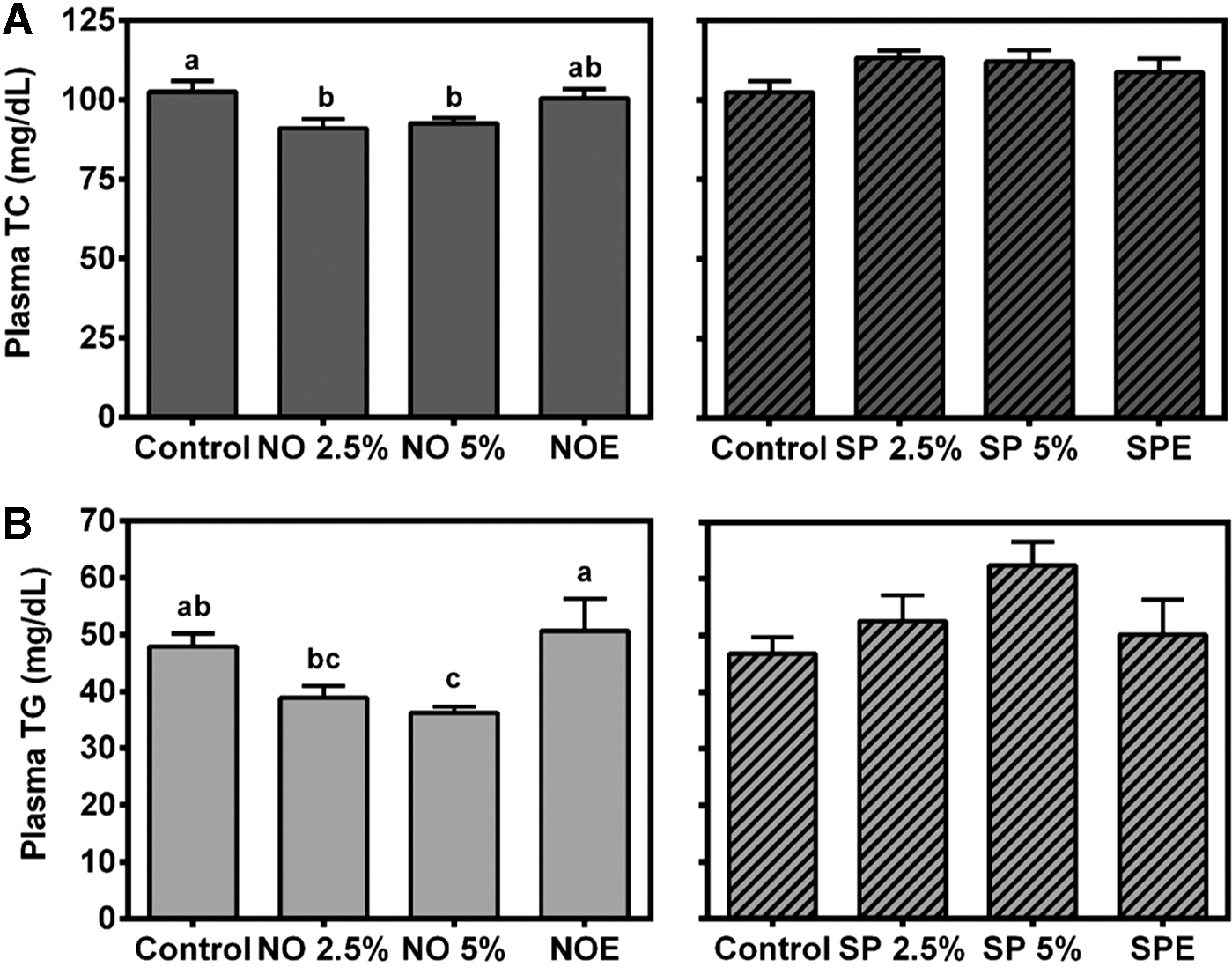
Plasma total cholesterol (TC) and triglyceride (TG) concentrations. Male C57BL/6J mice were fed an AIN-93M diet supplemented with 2.5% blue-green algae (BGA), 5% BGA, or lipid extract equivalent to 5% of BGA for 4 weeks. Plasma TC
Regulation of hepatic expression of genes related to cholesterol and lipid metabolism by BGA
As hepatic lipid levels largely affect plasma lipids, we next measured liver lipid contents. Body and liver weights of mice were not significantly different between groups (Table 3). Hepatic TC levels were not different between groups, but FC levels in SP 2.5%, SP 5%, SPE, and NO 2.5% groups were significantly higher than control, NO 5%, and NOE-fed groups (data not shown). Interestingly, TG levels in the livers of the SPE group were significantly higher than the NOE group. To gain insight into the hypolipidemic effect of NO, hepatic expression of genes involved in cholesterol and lipid metabolism was measured. Although both NO 2.5% and 5% groups showed significantly lower plasma TC concentrations than control, HMGR and SREBP-2 mRNA levels were significantly higher only in the NO 5% group (Fig. 2). We also examined the effect of BGA on the expression of lipogenic genes, that is, sterol regulatory element-binding protein 1c (SREBP-1c), fatty acid synthase (FAS), acetyl-coA carboxylase 1 (ACC-1), and stearoyl CoA desaturase 1 (SCD-1) in the liver. Hepatic expression of genes related to fatty acid oxidation, including carnitine palmitoyltransferase 1α (CPT-1α) and acyl-CoA oxidase 1 (ACOX-1), was also measured. Hepatic mRNA abundance of FAS, ACC-1, and SCD-1 was similar between groups, while SREBP-1c mRNA was significantly lower in the SP 2.5% group than controls (Table 4). In contrast, ACOX-1, the rate-limiting enzyme for peroxisomal fatty acid β-oxidation, was significantly increased by 5% NO supplementation. There also was an increasing trend in the expression of CPT-1α, the rate-limiting enzyme for mitochondrial fatty acid β-oxidation, in 5% NO-fed mice.
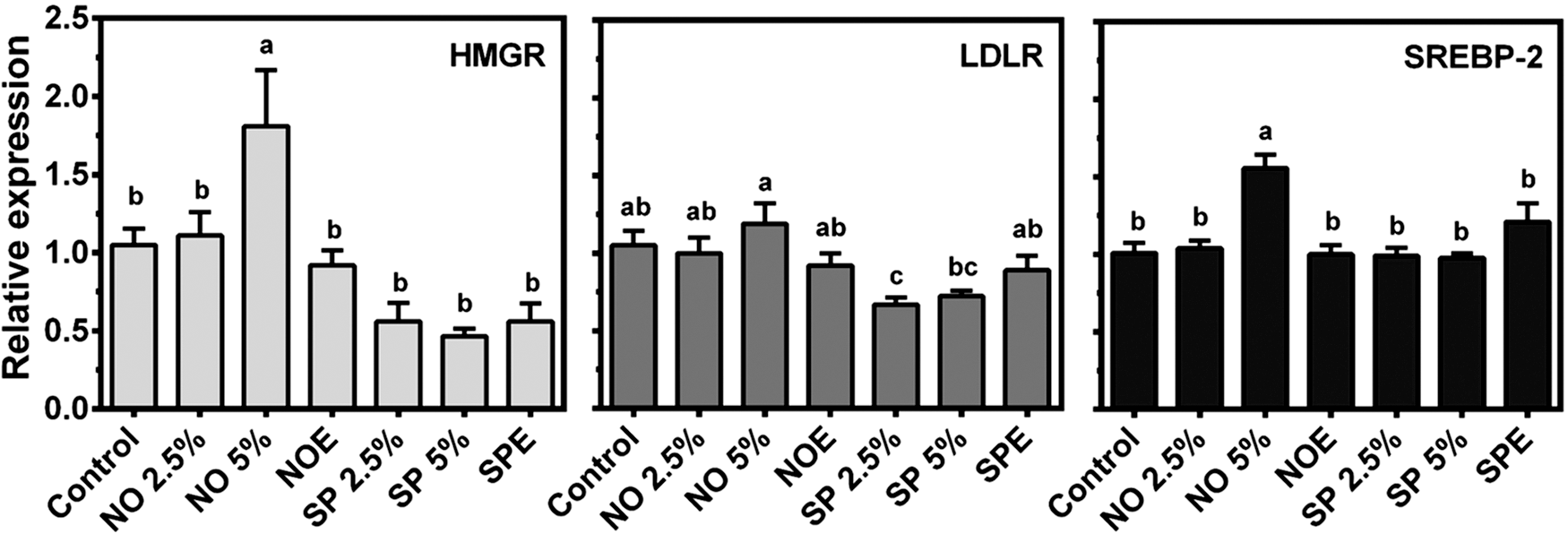
Hepatic expression of genes for cholesterol biosynthesis and uptake by quantitative real-time PCR (qRT-PCR). Total RNA was extracted from the livers of male C57BL/6J mice fed an AIN-93M diet supplemented with 2.5% BGA, 5% BGA, or lipid extract equivalent to 5% of BGA for 4 weeks for gene analysis by qRT-PCR. Data are expressed as relative expression to control, and bars with a different letter are significantly different (P<.05). Values are mean±SEM, n=10.
Values represent mean±SEM; n=10, except SP 5% (n=9).
Data with a different superscript are significantly different (P<.05).
TC, total cholesterol; TG, triglyceride.
Values represent mean±SEM; n=10, except SP 5% (n=9).
Data with a different superscript are significantly different (P<.05).
Effects of BGA supplementation on intestinal cholesterol absorption and fecal sterol excretion
We examined if the TC-lowering effect of 2.5% and 5% NO supplementation is attributed to decreased intestinal cholesterol absorption. Fractional cholesterol absorption in the intestine was significantly lower only in NO 2.5% and 5% groups than control (Table 5). However, SP and SPE supplementation did not significantly alter intestinal cholesterol absorption compared with control. Fecal neutral sterols were significantly higher in the SP 5% group, while the NO 2.5%-fed group showed a lower value compared with control. Fecal plant sterol excretion was significantly higher in NO 2.5% and 5%, as well as SP 5%, groups than control mice (Table 5).
Values represent mean±SEM; n=10, except SP 5% (n=9).
Data with a different superscript are significantly different (P<.05).
Effect of NOE and SPE on the expression of genes related to cholesterol and lipid metabolism in vitro
When HepG2 cells were treated with NOE or SPE, mRNA of HMGR and SREBP-2 was decreased in a dose-dependent manner (Fig. 3A). In addition, both NOE and SPE significantly decreased the expression of HMGR, SREBP-1c, FAS, and SCD-1, while only NOE repressed LDLR expression in HepG2 cells (Fig. 3B). However, the algal extracts markedly decreased protein levels of LDLR, HMGR, FAS, and mature SREBP-1 (mSREBP-1) (Fig. 3C). In primary mouse hepatocytes, the repressive effects of NOE on the expression of SREBP-2, LDLR, HMGR, SREBP-1c, and FAS, which was seen in HepG2 cells, were not observed (Fig. 3D). However, SPE was able to decrease HMGR, SREBP-1c, and FAS mRNA, while both algal extracts inhibited the expression of SCD-1 in primary hepatocytes. Both NOE and SPE markedly decreased protein levels of LDLR, HMGR, and FAS (Fig. 3E).
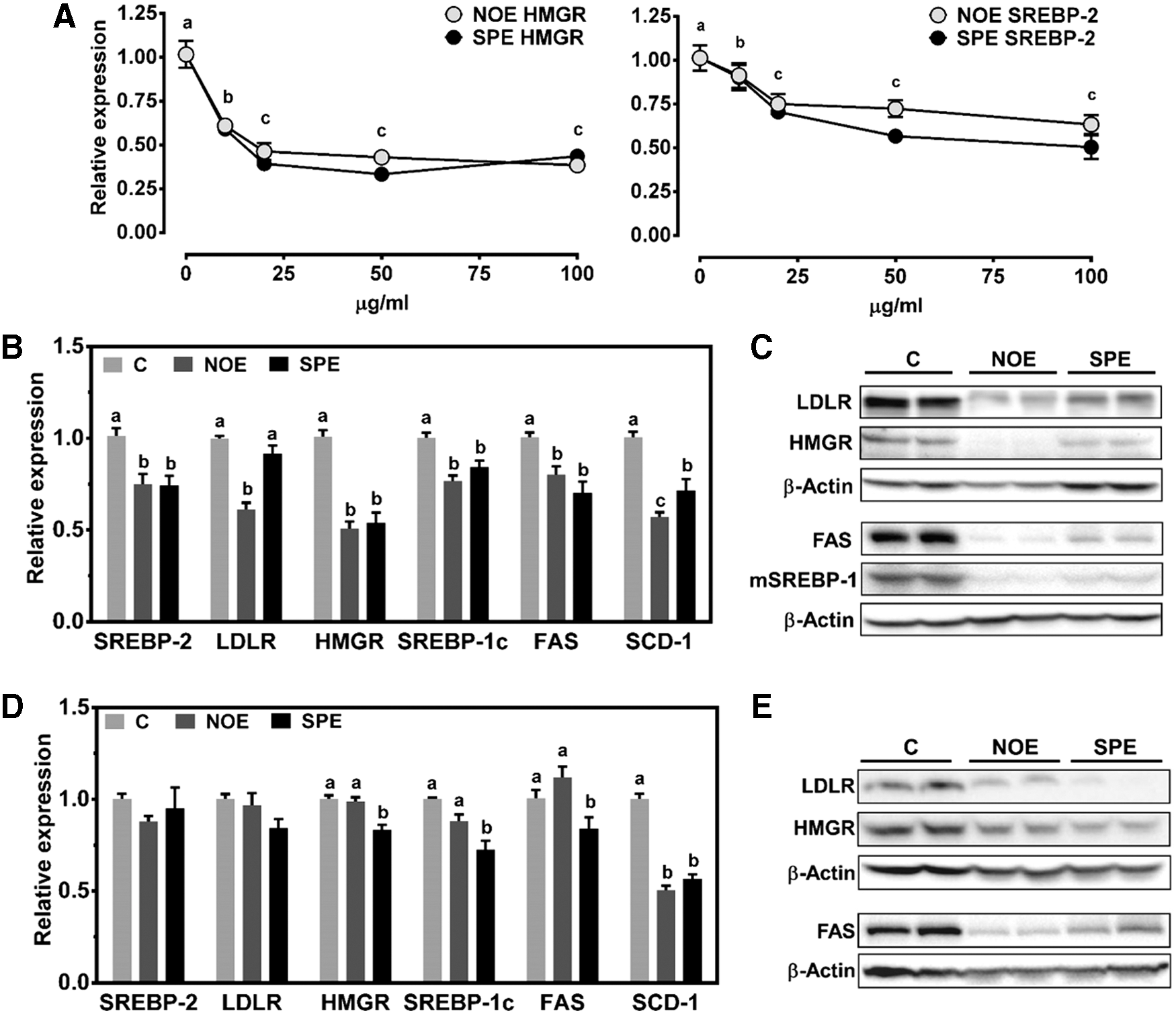
Effects of NO and SP lipid extract incubation on hepatic expression of genes for cholesterol biosynthesis and fatty acid metabolism. HepG2 cells
Discussion
Progression of atherosclerosis is accelerated by dyslipidemia. 22,23 There has been an increasing public demand for using safe and effective natural products as preventive and/or therapeutic agents against CVD. BGA have been shown to exert health-promoting effects. 11,12,24 –26 We previously observed that NOE significantly repressed HMGR and LDLR expression through downregulation of the SREBP-2 pathway in HepG2 cells. 15 NOE also repressed the lipogenic gene expression by inhibiting the activation of SREBP-1. In addition, in male C57BL/6J mice fed a regular chow diet supplemented with 5% whole dry NO for 4 weeks, plasma TC and TG levels were decreased and the cholesterol-lowering effect was attributed to a decrease in intestinal cholesterol absorption. 15 Interestingly, hepatic mRNA levels of HMGR, LDLR, and SREBP-2 were induced. The goals of this present study were to explain this apparent discrepancy between our in vivo and in vitro findings and to compare lipid-lowering effects of NO with SP. We demonstrated in the present study that the nonlipid fraction of NO, but not NOE, exerts plasma lipid-lowering effects by increasing intestinal cholesterol absorption, while SP did not lower the plasma lipids.
Consistent with our previous observation, 15 5% NO supplementation significantly lowered plasma TC and TG levels than control mice. While 2.5% NO supplementation decreased plasma TC to a similar degree as 5% supplementation, a significant hypotriglyceridemic effect of NO required 5% of the supplementation level. Of importance is that NOE supplementation, which contains lipid extract equivalent to 5% NO, did not lower plasma TC and TG, suggesting that a nonlipid fraction of NO is likely to be responsible for the hypolipidemic effect of NO. In contrast, SP at both 2.5% and 5% did not decrease plasma TC and TG. Studies have shown that SP supplementation improved plasma lipid profiles in hyperlipidemic rats and mice. 27,28 In humans, SP significantly lowered plasma TG levels in patients with type 2 diabetes 25,29 and with hyperlipidemic nephrotic syndrome. 30 Reasons for the discrepancy between the present study and aforementioned studies by others remain unclear. One possibility is that our study was conducted without dietary cholesterol and fat challenges, whereas the above-mentioned studies were conducted in the setting of hyperlipidemia. Dietary or metabolic challenges may be required to elicit the potential lipid-lowering effect of SP.
Studies suggested that inhibition of intestinal cholesterol absorption decreases plasma TC concentrations. 31 –34 It should be noted that NO supplementation at 2.5% and 5% significantly decreased intestinal cholesterol absorption, which was consistent with our previous study. As neither plasma TC nor intestinal cholesterol absorption were altered by NOE, compounds in the nonlipid fraction are presumed to play a critical role in the hypocholesterolemic effect of NO. Studies have shown that soluble fibers 35,36 and plant sterols 37,38 inhibit intestinal cholesterol absorption and consequently decrease plasma TC concentrations. NO contains ∼9% fiber, while SP has ∼6.8%. Additionally, our previous gas chromatograph analysis indicated that NO contains 3.25 μg/g of sitosterol with no detectable amount of cholesterol. Importantly, both 2.5% and 5% NO-fed mice showed significantly higher fecal plant sterol excretion, while only 5% SP displayed the same effect. Therefore, it can be speculated that the lack of lipid-lowering effects of SP compared with NO may be due to its less fiber content, although we cannot rule out the possibility that the two BGA may contain a different composition of fiber, that is, soluble and insoluble, which demonstrate different hypolipidemic potency. The critical role of fibers in lowering plasma cholesterol was also seen in rats fed 5% NO. 12 In addition to fiber and plant sterols, C-phycocyanin, a phycobiliprotein highly present in BGA, 39 is known to possess hypocholesterolemic properties by lowering solubility of cholesterol in micelle, consequently decreasing cholesterol absorption and increasing fecal neutral and acidic sterol output. 40
SREBPs are transcription factors that play a critical role in cholesterol and fatty acid metabolism. 41,42 SREBP-2 increases the transcription of genes involved in the cholesterol biosynthetic pathway and uptake, including HMGR, LDLR, HMG-CoA synthase, and squalene synthase, in response to cellular cholesterol depletion. 43 –45 Although mRNA expression of HMGR and SREBP-2 was significantly higher in 5% NO-fed mice than the other groups, protein levels of HMGR, mature SREBP-2, and LDLR were significantly different between groups (data not shown). We previously observed that NO markedly induced the expression of cholesterol 7α hydroxylase, the rate-limiting enzyme for bile acid synthesis, in the liver, but fecal acidic sterol output was not altered. 15 Therefore, the TC-lowering effect of NO may not be mediated through LDLR-mediated LDL uptake and conversion of cholesterol to bile acids in the liver. Mechanisms underlying the hypocholesterolemic effect of NO are currently elusive. Whether NO alters cholesterol metabolism in the intestine to lower plasma cholesterol is another possibility to be tested. In contrast to SREBP-2, SREBP-1c primarily regulates the expression of lipogenic genes, such as FAS, ACC-1, and SCD-1. 46 –48 Although 5% NO supplementation decreased plasma TG levels, hepatic expression of SREBP-1c and its target genes for lipogenesis was not altered in this group of mice. In contrast, ACOX-1 mRNA levels were significantly increased and CPT-1α showed an increasing trend in the NO 5% group compared with control. CPT-1α and ACOX-1 are involved in mitochondrial and peroxisomal fatty acid β-oxidation, respectively. 49,50 These genes are under the transcriptional regulation by peroxisome proliferator-activated receptor α (PPARα). Fibrates, PPARα agonists, have been shown to lower plasma TG concentrations. 51 –53 Whether NO contains potential PPARα agonists is an interesting question, and identification of bioactive compounds needs to be conducted in the future.
Although both NOE and SPE did not alter the expression of genes related to cholesterol and fatty acid metabolism in vivo, there were potent in vitro effects of the algal extracts on the gene expression. In HepG2 cells, the expression of SREBP-2 and its target genes, that is, LDLR and HMGR, was inhibited, except LDLR mRNA in SPE-treated cells. Furthermore, SREBP-1c, FAS, and SCD-1 mRNA levels were significantly decreased. Stronger inhibition of their protein levels than mRNA was displayed in NOE or SPE-treated cells. The similar repression of the cholesterogenic and lipogenic genes was also observed in primary mouse hepatocytes. There are several possibilities for the absence of these potent inhibitory effects of NOE and SPE in mice. We speculated that given the very low level of lipid fraction in NO and SP, the potential bioactive compounds in the lipid fraction might not be sufficient to elicit their effect in vivo. Alternatively, putative bioactive compounds that modulate the expression of genes mentioned above may be metabolized, losing their effect. Nonetheless, our in vitro observations indicate that the nonlipid fraction of NO is likely responsible for its lipid-lowering effect.
In conclusion, NO, but not NOE, supplementation exerted plasma TC and TG-lowering effects. This supports our hypothesis that the nonlipid fraction of NO is likely to contain bioactive compounds responsible for the hypolipidemic effect, such as fiber and plant sterols. SP, the most commonly consumed BGA by humans, did not lower plasma TC and TG. However, in apolipoprotein E knockout mice, a mouse model of atherosclerosis, SP inhibited atherosclerotic lesion formation as effectively as did NO, although it did not lower plasma lipids (manuscript in preparation). Therefore, SP can provide an atheroprotective effect by mechanisms other than lowering plasma lipids. We previously demonstrated that NO and SP used in this study are free of microcystins, major algal toxins, and 2.5% and 5% of NO and SP supplementation for 6 months did not have any adverse side effects on growth, plasma transaminase levels, and liver histology in mice. 54 Therefore, NO and SP are likely to be safe natural products that can be used to prevent CVD risk. Well-designed human clinical trials are necessary to determine the efficacy of NO and SP in preventing CVD.
Footnotes
Acknowledgments
C.S.K. played a major role in conducting experiments and writing the manuscript; B.K., T.X.P, Y.Y., and Y.P. contributed to the experiments; C.L.W. extracted the lipid from BGA to formulate the experimental diet; T.P.C. conducted and analyzed fecal sterols; J.L. designed the experiments and contributed to data analysis and manuscript preparation. This work was supported by a National Institutes of Health grant, R21AT005152, and a fund from the College of Agriculture and Natural Resources at the University of Connecticut to J.L.
Author Disclosure Statement
No competing financial interests exist.