
Abstract
Current experimental studies show that Ocimum tenuiflorum (commonly known as basil or Tulsi) possesses many health benefits. Ocimum is suggested to be antioxidative and anti-inflammatory. Eugenol, an orthomethoxyphenol, and ursolic acid have been identified as important components of basil. Myeloperoxidase (MPO), an oxidative enzyme, has been implicated in the pathogenesis of atherosclerosis. MPO-dependent oxidation of lipoproteins has been implicated in foam cell formation, dysfunctional HDL, and abnormalities in reverse cholesterol transport. Whole leaf extract of O. tenuiflorum and its major components, eugenol and ursolic acid, inhibit the oxidation of lipoproteins by myeloperoxidase/copper as measured by conjugated diene formation as well as by the thiobarbituric acid reactive substance (TBARS) assay. Whole basil leaf extract is able to attenuate the lipopolysaccharide-induced inflammation in RAW 264.7 cells compared with its components. In addition, whole basil leaf extract and eugenol inhibited MPO enzyme activity against synthetic substrates. Based on these results, we conclude that basil extract could act as an inhibitor of MPO and may serve as a nonpharmacological therapeutic agent for atherosclerosis.
Introduction
P
Eugenol (4-allyl-2-methoxyphenol) is a naturally occurring phenolic compound in basil, cinnamon, and nutmeg and is the major component of clove oil. It is widely used as a component of zinc oxide eugenol cement in dentistry and is applied to the oral environment. 13 In addition, eugenol is a flavoring agent in cosmetic and food products. 14 Eugenol has been shown to possess many medicinal properties, such as antispasmodic, 15 antipyretic, 16 anti-inflammatory, 17 and antibacterial activities. 18 Recently, it is reported that eugenol inhibits 5-lipoxygenase enzyme by a noncompetitive mechanism. 19
Ursolic acid (UA) is another component associated with basil and was identified as the major component in O. tenuiflorum. 20 It has been reported that ursolic acid has protective properties similar to that of basil. Several mechanisms have been proposed to explain its anti-inflammatory activity, including inhibition of secretory phospholipase (PLA2) enzymes 21 and inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2). 22
Existing evidence suggests that myeloperoxidase (MPO) plays a major role in the oxidative modification of both low-density lipoprotein (LDL) and high-density lipoprotein (HDL). MPO is a heme protein expressed at high levels in neutrophils, monocytes, and subpopulations of human macrophages. Catalytic activity of MPO is responsible for the generation of reactive intermediates that can promote further oxidation. 23,24 One feature that distinguishes MPO from other peroxidases is its ability to generate hypochlorous acid that may mediate formation of distinct chlorinated derivatives. Furthermore, its effects on lysine, methionine, and tyrosine residues within proteins may have additional effects on the tertiary structure and function of proteins. Therefore, MPO of inflammatory cells has considerable potential to cause tissue damage. It is also known that MPO converts LDL into a proatherogenic form within the artery wall. A role for this enzyme in the oxidation of apolipoprotein A1 (Apo A1) of HDL has also been recognized. The oxidation of critical amino acid residues of Apo A1 has been shown to reduce the cholesterol efflux function of HDL. 25 –27 Inflammation plays a major role in the progression of atherosclerosis, and oxidized (Ox)-LDL/HDL-triggered inflammation is considered to be one of the major causes of atherosclerosis.
In this study, we determined the antioxidant property of the basil leaf aqueous extract on lipoproteins and its anti-inflammatory property on lipopolysaccharide (LPS)-induced inflammation in vitro.
Materials and Methods
Chemicals
All chemicals were purchased from Sigma Chemicals (St. Louis, MO, USA); pimers and cell culture reagents were purchased from Invitrogen (Carlsbad, CA, USA).
Preparation of crude basil leaf extract
Basil extract (BE; aqueous) was prepared by macerating 10 g of dried basil leaf powder with 100 mL sterilized water and then filtered through Wattman filter paper. The filtrate was lyophilized and reconstituted with 10 mL of sterilized water. Approximately 2 μg of extract was used to run the spectrum between 200 and 300 nm. The spectra for eugenol and ursolic acid were also run at the same wavelength scan (data not shown).
Isolation and oxidation of lipoproteins
The study protocol was approved by the University of Central Florida Institutional Review Board. Blood was collected in heparinized tubes from consenting healthy donors and stored on ice. Blood was centrifuged at 450 g for 20 min and plasma was separated. Lipoproteins were isolated from normal plasma by sequential ultracentrifugation using a Beckman TL-100 tabletop ultracentrifuge (Beckman, Palo Alto, CA, USA). 28 The isolated lipoproteins were dialyzed against 0.3 mM EDTA in 1×phosphate-buffered saline (PBS) of pH 7.4 overnight and subsequently filter sterilized. The amount of protein was estimated using the Bio-Rad DC protein assay (Hercules, CA, USA). The LDL/HDL sample was subjected to oxidation immediately after dialysis. Oxidation of LDL/HDL was performed 29 with 5 μM copper or MPO (0.2 U) and 100 μM H2O2 to a 100 μg/mL of LDL in 1 mL of PBS at 37°C both in the presence and absence of different concentrations of either basil extract (2–50 μg) or its components, eugenol or ursolic acid (5–25 μM). The formation of conjugated dienes was monitored at an optical density of 234 nm for about 4–6 h using the SLM Aminco DB-3500 spectrophotometer (Urbana, IL, USA) equipped with a 12-chamber cuvette changer. The degree of LDL/HDL oxidation was assessed by determination of peroxide content using leucomethylene blue (LMB) assay 30 and thiobarbituric acid reactive substance (TBARS) assay. 31 Electrophoretic mobility of Ox-LDL was determined by using agarose gel electrophoresis.
Preparation of HPODE and incubation with basil extract, eugenol, and ursolic acid
13-Hydroperoxylinoleic acid (13-HPODE) 32 of 200 nmoles/mL was prepared and used to determine the effect of the whole basil extract and its components, eugenol and ursolic acid, on free fatty acid peroxides (FFAOOHs). HPODE was incubated with increasing concentrations of basil extract (2–20 μg), eugenol (1–10 μM), and ursolic acid (5–25 μM) for 1 h at room temperature. Lipid peroxide generated in the reaction system was analyzed by LMB assay. 30
Cell culture
RAW 264.7 cells were obtained from ATCC. Cells were grown as a monolayer in flasks and dishes and maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum (Sigma Chemicals), 2 mM
WST-1 for cell viability assay
Cells (2.5×103 cells/100 μL) were seeded in a 96-well plate and incubated overnight at 37°C under a 5% CO2 atmosphere. The medium in the wells was then replaced with fresh medium containing whole basil leaf extract (5–100 μg) and its components, eugenol and ursolic acid (1–50 μM), and incubated for 24 h. The effect of the basil leaf extract and its components on cell viability was determined by WST-1 assay (Roche Applied Science, Indianapolis, IN, USA). Briefly, 10 μL of WST-1 reagent, which is processed to formazan by cellular enzymes, was added to each well in 100 μL of cell culture medium, and the plates were further incubated for 4 h at 37°C. Cell viability was calculated by measuring absorbance at 450 and 630 nm as reference wavelengths using a 96-well plate reader (Bio-Rad).
Incubation of RAW 264.7 cells with LPS in the presence and absence of eugenol/BE/UA
To monitor the changes in gene expression of tumor necrosis factor-α (TNF-α), interleukin (IL)-1α, IL-1β, and IL-6, RAW 264.7 cells (2×105 cells/well) were incubated in serum-free RPMI 1640 for 4 h. Ten millimolar concentrations of eugenol and ursolic acid were prepared in PBS and used. Cells were pretreated with 5 and 25 μM of eugenol and ursolic acid, 10 and 50 μg of BE for 2 h, followed by addition of 10 ng/mL LPS. At the end of the 24-h incubation, cells were harvested in Trizol™ for RNA isolation.
Thin-layer chromatography and SDS-PAGE for aqueous extract
An amount of 5 μL of basil extract (2 mg/mL), eugenol, and ursolic acid (5 mg/mL in chloroform) were loaded onto silica G thin-layer chromatography (TLC) plates and fluorescent plates using chloroform/ethylacetate/acetic acid (10:2.5:0.05 v/v) as the solvent system. Visualization of the components was carried out with a brief exposure to iodine vapors and with FeCl3 for visualization of fluorescent spots from unsaturated compounds.
Twelve percent sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was used to determine the protein content in basil aqueous extract by both coomassie staining and silver staining.
cDNA synthesis and real time-polymerase chain reaction
Total RNA from cells was isolated by using Trizol reagent. One microgram of RNA was reverse transcribed into cDNA using the Superscript™ III First-Strand Synthesis system (Invitrogen). cDNA (50 ng) sample was used to perform quantitative real-time PCR reaction by the iQTM5 iCycler Multicolor Real-Time PCR Detection system (Bio-Rad) with SYBR Green (Invitrogen). Polymerase chain reaction (PCR) was carried out with IL-1α, IL-1β, IL-6, and TNF-α-specific primers for mouse (Table 1), resulting in 200- and 400-bp fragments. As a reference gene, we used GAPDH primers, resulting in a 200-bp fragment. PCR was performed with an initial step of denaturation at 50°C for 2 min, 95°C for 10 min, followed by 40 cycles at 95°C for 20 sec, and 60°C for 20 sec. Melt curves were established for the reactions. Normalized fold expression was calculated by using the 2−ΔΔCt method. The entire reaction product was electrophoresed on 2% agarose gels. Experiments were repeated more than three times.
IL, interleukin; TNF-α, tumor necrosis factor-α.
Inhibition of peroxidase activity of MPO by basil extract and eugenol
MPO activity was measured colorimetrically by oxidation of substrate guaiacol and monitored by absorbance at 470 nm. 33 A reaction volume of 200 μL contained 25–100 ng of basil extract, 0.25–10 μM eugenol in phosphate buffer (pH 7.4), 0.5 μmoles guaiacol, 10 nmoles H2O2, and 0.1 U of MPO. The change in optical density at 470 nm was measured over 5 min using a 96-well plate reader (Bio-Rad). Relative MPO activity was calculated and used.
MPO activity was also measured colorimetrically by oxidation of substrate TMB (3,3′,5,5′-tetramethylbenzidine) and monitored by absorbance at 650 nm. 34 A reaction volume of 200 μL contained 25–100 ng of basil extract, 0.25–10 μM eugenol and ursolic acid, 50 mM sodium acetate buffer (pH 5.6), 25 nmoles TMB, 400 nmoles H2O2, and 0.02 mU of MPO. The change in optical density at 650 nm was measured every 10 min using a 96-well plate reader (Bio-Rad). Relative MPO activity was calculated and used.
Statistical analyses
Values are presented as mean±standard deviation, and statistical analyses were performed by using Student's t-test at significance of P<.05.
Results
Basil extract and its components, eugenol and ursolic acid, inhibit the oxidation of lipoproteins (LDL and HDL)
To test the effect of BE and its components on the oxidation of LDL, the lipoprotein was incubated with 5 μM copper or 0.2 U MPO, and oxidation was performed in the presence and absence of different concentrations of BE, eugenol, and ursolic acid. The formation of conjugated dienes was measured by following the increase in absorption at 234 nm.
35
As shown in Figure 1, control incubation generated an oxidized LDL that reached maximum increase in absorption at about 200 min. The lag phase, which represents the time at which antioxidants are consumed, was 60–80 min. As shown in Figures 1 and 2, in the presence of increasing amounts of either basil extract or eugenol/UA, there was an increase in lag time, suggesting that even low concentrations are able to delay the oxidation rate. Reduced thiobarbituric reactive substances were observed in the presence of BE and eugenol/UA compared with Ox-LDL, as shown in Supplementary Figures S1 and S2 (Supplementary Data are available online at
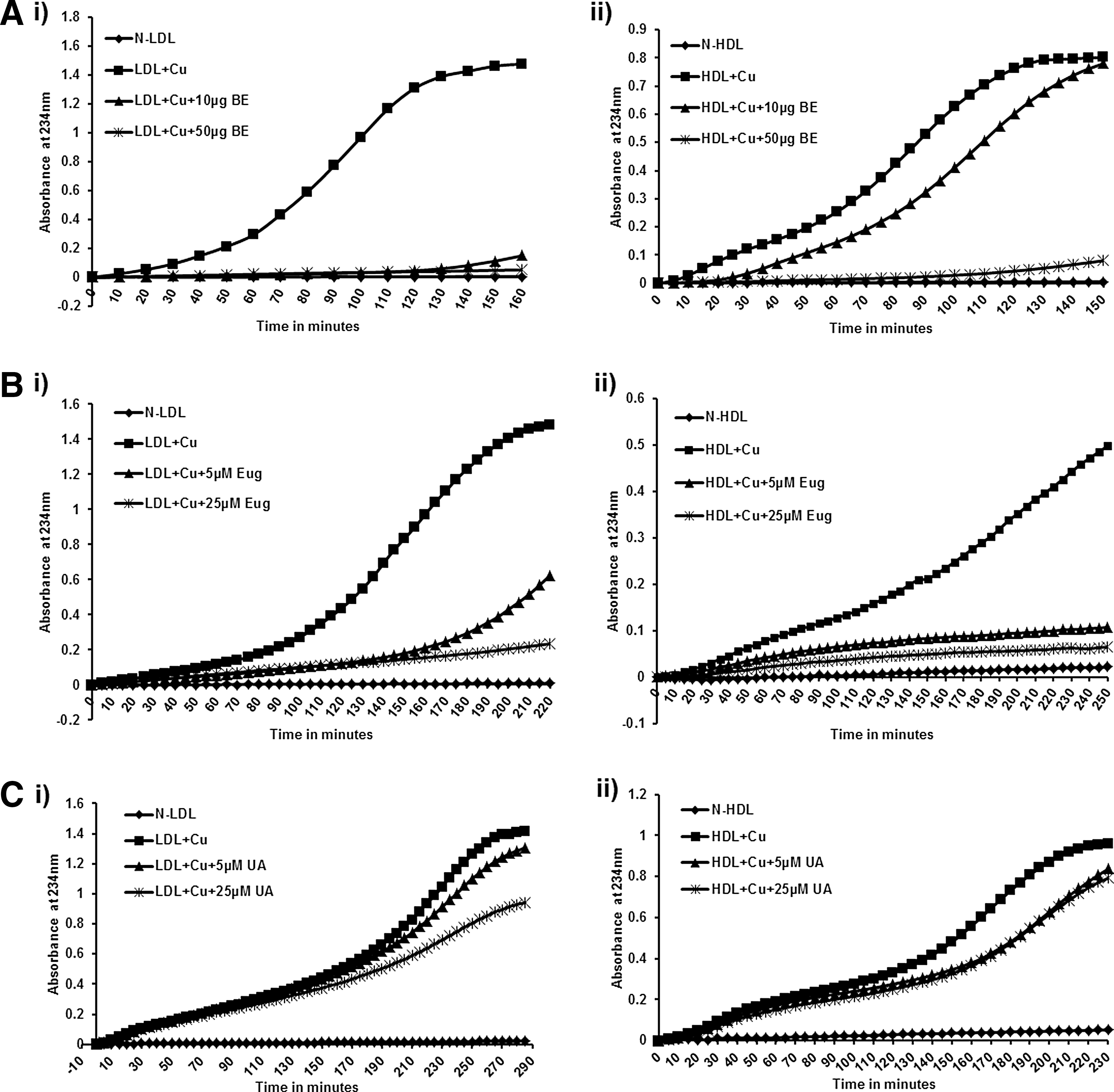
Basil extract (BE) and its components inhibit the oxidation of lipoproteins by copper. Lipoproteins were isolated from the plasma of consenting subjects and used for oxidation with copper. One hundred micrograms of low- or high-density lipoprotein (LDL/HDL) was pretreated with BE (10 and 50 μg) and its components, eugenol and ursolic acid (UA), at different concentrations (5 and 25 μM) for 1 h at room temperature, and then oxidation was performed with 5 μM copper in 1 mL phosphate-buffered saline (PBS); OD was measured at 234 nm. As concentrations of BE

Basil extract and its components inhibit the oxidation of lipoproteins by myeloperoxidase (MPO). Lipoproteins were isolated from the plasma of consenting subjects and used for oxidation with copper. One hundred micrograms of LDL/HDL was pretreated with BE (10 and 50 μg), and its components, eugenol and ursolic acid, at different concentrations (5 and 25 μM), were incubated for 1 h at room temperature, and then oxidation was performed with 0.2 U MPO in 1 mL PBS; OD was measured at 234 nm. As concentration/volume of BE
Basil extract and its components, eugenol/UA, were also tested on HDL oxidation in the presence and absence of 5 μM copper (Fig. 1) or 0.2 U MPO, as shown in Figure 2. Similar results as seen with Ox-LDL were observed.
Reduction of peroxides in the presence of BE/eugenol/UA
FFAOOHs were reduced into hydroxides in the presence of increasing concentrations of basil extract and eugenol. As shown in Figure 3, both components showed a significant reduction in the peroxide content of FFAOOHs with increasing concentration, whereas ursolic acid did not show any effect on peroxide decomposition (Fig. 3).
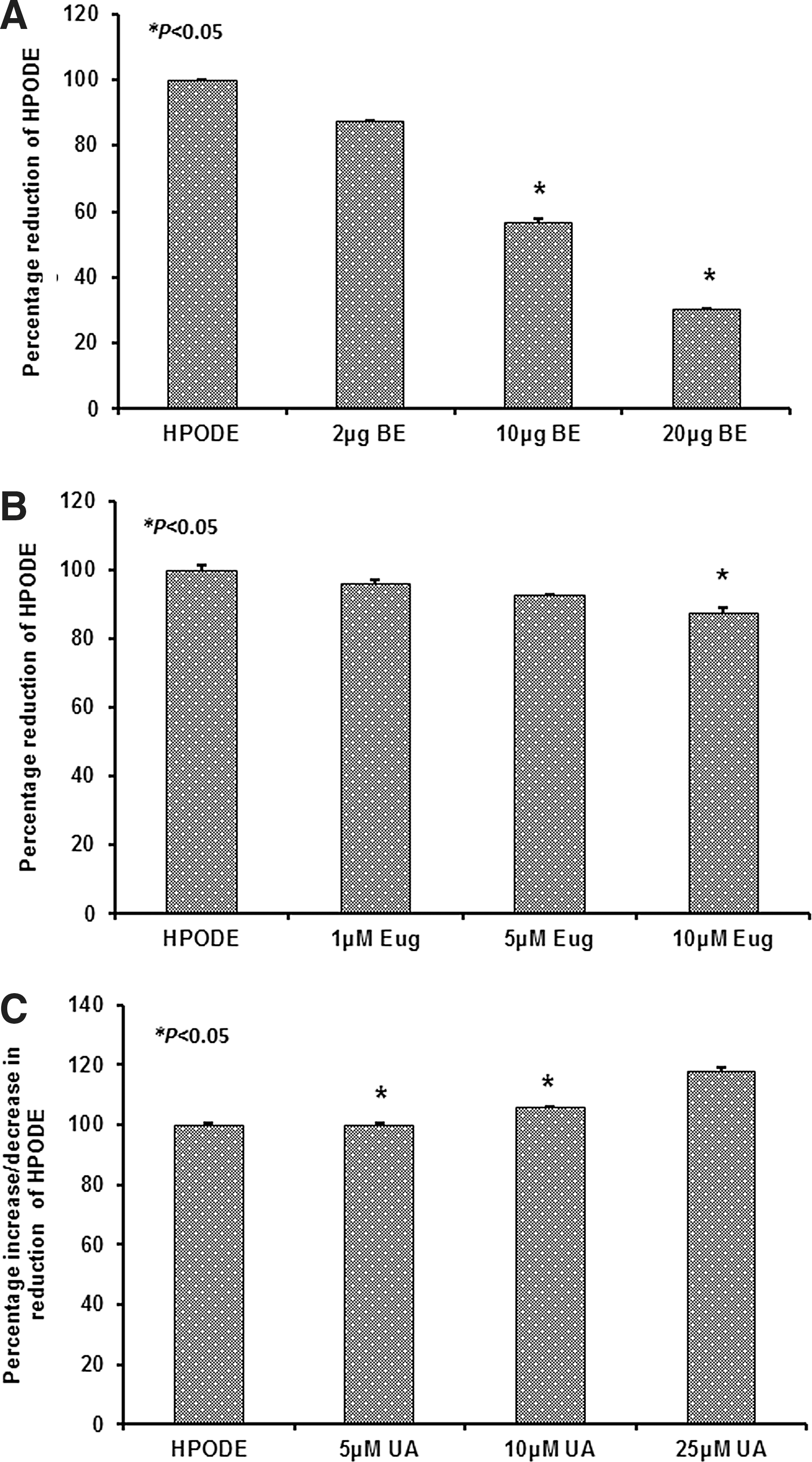
Decomposition of HPODE in the presence of BE and its components. Two hundred nmoles per mL of HPODE was prepared by oxidizing linoleic acid with soybean lipoxygenase and used to determine the effect of BE and its components on free fatty acid peroxides (FFAOOHs). HPODE was incubated with increasing volume/concentrations of
TLC and SDS-PAGE for BE component identification and protein determination
In BE, spots corresponding to eugenol were observed with iodine staining, whereas the ursolic acid spot was not observed. Similarly, with FeCl3 staining for unsaturated compounds, only the eugenol spot was observed, whereas the others were not seen. The aqueous extract components are less polar than the corresponding components as shown in Figure 4.

Thin-layer chromatography for BE and its components. Dotted circle represents the corresponding spot of eugenol.
Basil extract was further analyzed by SDS-PAGE gel to determine the protein components. After confirming the absence of considerable protein content (Supplementary Fig. S3), basil extract was used for in vitro cell culture studies.
Cell viability assay
A WST-1 assay was used to evaluate the cell viability of RAW 264.7 macrophages in the presence and absence of BE and its components, eugenol and UA. Cells were incubated with BE (5–100 μg) and its components, eugenol and UA (1–50 μM), for 24 h and used for the assay as described in the Materials and Methods section. Percentage of cell viability was calculated using control as 100% viability. As shown in the Supplementary Figure S4, BE and eugenol have no obvious toxic effects on macrophages, whereas UA at higher concentrations has a slight, but insignificant, toxic effect.
Basil extract and its components attenuate LPS-induced IL-1α, IL-1β, IL-6, and TNF-α mRNA expression
As seen in Figure 5, BE and its components attenuated LPS (10 ng/mL)-induced proinflammatory markers (Fig. 5A: IL-1α, Fig. 5B: IL-1β, Fig. 5C: IL-6; Fig. 5D: TNF-α) in RAW cells to varying degrees. Significant attenuation was observed with BE, whereas eugenol and UA were less effective at the concentration used. As shown in Figure 5E, PCR products also corroborated the gene studies.

Basil extract and its components attenuate lipopolysaccharide (LPS)-induced proinflammatory gene expressions in RAW 264.7 cells. RAW 264.7 cells were incubated with BE (50 μg) and eugenol/UA (25 μM) for 2 h, followed by LPS (10 ng/mL) for 24 h. RNA was isolated and Real Time-PCR analysis was performed for
MPO activity
The effect of BE and eugenol on MPO activity was studied. As shown in Figure 6, MPO activity was completely inhibited in the presence of BE and eugenol in a dose-dependent manner with both substrates, guaiacol (6Ai and ii) and TMB (6Bi and Bii).

MPO activity. MPO activity was performed for BE
Discussion
Oxidative stress plays a significant role in the pathogenesis of several cardiovascular diseases. Current experimental and clinical studies suggest that O. tenuiflorum (commonly known as basil or Tulsi) possesses many health benefits. Oxidation of lipids is assumed to be implicated in the pathophysiology of atherosclerosis. It has been suggested that scavenging of lipid peroxyl radicals contributes to the antiatherosclerotic effects of naturally occurring compounds such as polyphenols. These compounds are capable of inhibiting the lipoprotein oxidation in vitro and suppressing the formation of plasma lipid oxidation products in vivo. 32 Therefore, inhibition of LDL oxidation might be an important step in preventing atherosclerosis. Humans are protected from reactive oxygen species, in part, by absorption of dietary antioxidants. 36
The ability of BE and its components to inhibit the oxidation of LDL by copper or MPO in an in vitro system suggests that they could prevent the formation of Ox-LDL in vivo. While this is interesting, numerous other antioxidants may also act more effectively and more inexpensively. This might be due to the ability of basil extract and its components to decrease the rate of initiation of oxidation by reacting with oxygen radicals. They might also act as chain terminators and prevent propagation of oxidation. Pretreatment of LDL with basil extract and its components renders LDL resistant to oxidation, suggesting that these compounds might be able to remove 13(S)-HPODE and 15(S)-HPETE from LDL, which are the major enhancers of the nonenzymatic oxidation of both 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphocholine and cholesterol linoleate. 37,38 It is assumed that MPO secreted in the extracellular fluid is responsible for the generation of Ox-LDL, and whether peroxidized lipids formed on the cell membranes during ischemia/reperfusion injury may also be responsible could only be speculated.
HDL plays a major role in reverse cholesterol transport (RCT) and may carry not only cholesterol but also phospholipids and oxidized lipids from foam cells. HDL has several functions beyond its role as an agent of RCT. It interferes with the oxidative modification of LDL (although it is also readily oxidized) and has enzymes such as paraoxonase 1 (PON1) associated with it. It is also reported to bind and reduce peroxides. 39 These putatively beneficial properties and the association of proinflammatory serum amyloid A and MPO has given rise to the concept of functionality of HDL. MPO-Ox-HDL, as MPO has been identified as an APOA 1 modifier under oxidative stress conditions, can lead to loss of anti-inflammatory and antioxidant properties of HDL and change to be proinflammatory in nature in vivo. Prevention of HDL oxidation by these components will retain all its native functions in addition to RCT, such as the anti-inflammatory, antithrombotic, cytoprotective, anti-infectious, and vasodilatory natures.
The ability of BE and its components to reduce FFAOOHs is exciting and could have a major impact in preventing propagation of oxidation as well as in reducing the cellular effects of lipid peroxides. Eugenol is a compound containing methoxyphenol, which is a major source of many of the drugs of dentistry. Evidence suggests that the compounds associated with this group have the capacity to act as MPO inhibitors. We also tested the effect of guaiacol, which is a precursor for eugenol, and vanillin, also considered as an oxidizer, has shown similar effect on peroxide formation at a higher concentration (data not shown). This might be due to the presence of a methoxy group, which acts as an MPO inhibitor also inhibiting the soybean lipoxidase action. There is a reasonable homology of soybean lipoxidase with human 15 lipoxygenase and myeloperoxidase showing ∼19.9% and 11% similarity, respectively.
LPS is an endotoxin that is an integral structural component of the outer membrane of gram-negative bacteria. It is released during cell division, cell death, or during antibiotic treatment against bacterial infection and leads to activation of macrophages and enhanced production of proinflammatory cytokines. 40 Although generation of proinflammatory cytokines is essential for the development of the local inflammatory response, an unbalanced and sustained overproduction of such cytokines may lead to septic shock characterized by endothelial damage, loss of vascular tone, coagulopathy, and multiple system organ failure, often resulting in death. 41 It has been identified that eugenol and ursolic acid prevent LPS-induced cytokine induction in macrophages at a higher concentration, particularly above 50 μM. 22,42
Oxidative stress and inflammation are closely associated in atherosclerosis. At what stage of the disease inflammation plays a pivotal role is unknown. The results of our study suggest that basil extract and its components are able to inhibit the oxidation of LDL, and BE is able to attenuate LPS-induced inflammation more than its individual components. Basil leaves can be simple nonpharmacological agents, which could prove invaluable as an adjunct therapy in the treatment of cardiovascular diseases. This study will form the basis for many future studies not only in atherosclerosis but also in other oxidative stress-related diseases, including diabetes, cancer, obesity, and hypertension.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.