
Abstract
Oxidative stress has a dual role in cancer; it is linked with tumorigenic events and host wasting, as well as senescence and apoptosis. Researchers have demonstrated the importance of coadjuvant therapies in cancer treatment, and Aloe vera and honey have immunomodulatory, anticancer, and antioxidant properties. The preventive and therapeutic effects of Aloe vera (L.) Burm. f. (Xanthorrhoeaceae) and honey in tumor progression and host wasting were analyzed in Walker 256 carcinoma-bearing rats. The animals were distributed into the following groups: C=control-untreated, W=tumor-untreated, WA=treated after tumor induction, A=control-treated, AW=treated before tumor induction, and AWA=treated before and after tumor induction. Proteolysis and oxidative stress were analyzed in the tumor, liver, muscle, and myocardial tissues. The results suggest that the Aloe vera and honey treatment affect the tumor and host by different mechanisms; the treatment-modulated host wasting and cachexia, whereas it promoted oxidative stress and damage in tumor tissues, particularly in a therapeutic context (WA).
Introduction
O
Cancer patients may show progressive weight loss, edema, muscle and adipose tissue atrophy, and fatigue as a result of host catabolic processes, involving oxidative stress and proteolysis, driven by the tumor. These events characterize a syndrome known as cachexia that affects approximately 30–80% of the patients depending on tumor type, jeopardizing quality of life and survival. 4
Over the last decades, researchers have demonstrated that the use of complementary and coadjuvant therapies can improve the prognosis and quality of life for cancer patients. 9,10 Aloe vera (L.) Burm. f. (Xanthorrhoeaceae) possesses some pharmacological activities, including immunomodulatory, antioxidant, and anticancer properties. 11 A. vera increases the hepatic levels of antioxidant and drug-metabolizing enzymes and decreases oxidative stress in mice. 12 In contrast, numerous A. vera compounds have been reported to generate oxidative stress and apoptosis in cancer cells in vitro. 13,14 Concurrently, studies have demonstrated that honey and its phenolic compounds may possess anticancer and antioxidant activities. Indeed, a sustained inverse relationship between honey consumption and cancer development has been reported. 15
Despite the fact that cancer patients commonly use the combination of A. vera and honey as a complementary therapy, few studies demonstrating real benefits of this association have been reported. 16,17 Therefore, this study aims to evaluate the effects of A. vera and honey treatment on oxidative stress and proteolysis in the host and tumor tissues of Walker 256 tumor-bearing rats.
Materials and Methods
Aloe vera and honey solution
The sample leaves of Aloe vera (L.) Burm. f. (Xanthorrhoeaceae) were collected in December 2008 by Dr R Tomasin in the Green House of Biology Institute of State University of Campinas—UNICAMP (local 22°49′10.3″S 47°04′15.5″W), São Paulo, Brazil. The specimen was identified and authenticated by Dr. João Semir, and the A. vera voucher specimen (bach number #183049) was deposited in the Herbarium of the Botanical Department/IB/UNICAMP.
The whole mature and healthy A. vera leaves (500 g) were collected, washed, and cleaned in running water; following, the leaves were homogenized in polytron (200 g) with wildflower honey (500 g; Lambertucci Ltd., Rio Claro, Brazil) in aseptic conditions, and the whole homogenized solution was maintained in dark vials at 4°C. 17
Experimental protocol
Adult male Wistar rats (21 days old, approximately 50 g body weight) were initially divided into two groups: (1) The control (C) (n=30) group received 670 μL/kg of 0.9% NaCl solution daily by gavage and (2) the A. vera- and honey-treated group (A) (n=30) was gavaged with a 670 μL/kg dose of the A. vera and honey solution daily. At the age of 100 days, the rats were redistributed according to the Walker 256 carcinoma subcutaneous implantation (2×106 viable tumor cells/rat in 0.5 mL inoculum) and treatment.
The control group was divided into the following three subsets: the control (C, n=10) and tumor-control (W, n=10) groups continued to receive 670 μL/kg of 0.9% NaCl solution daily, while the tumor-A. vera+honey group (WA, n=10) received 670 μL/kg of the A. vera and honey solution after the tumor induction. Concurrently, the A. vera and honey-treated group was also divided into the following three subsets: the control-A. vera+honey (A, n=10) and A. vera+honey-tumor-A. vera+honey (AWA, n=10) groups continued to receive 670 μL/kg of the A. vera and honey solution daily, while the A. vera+honey-tumor (AW, n=10) group received the 0.9% NaCl solution daily after the tumor implant. The groups are outlined in the scheme given below (Fig. 1).
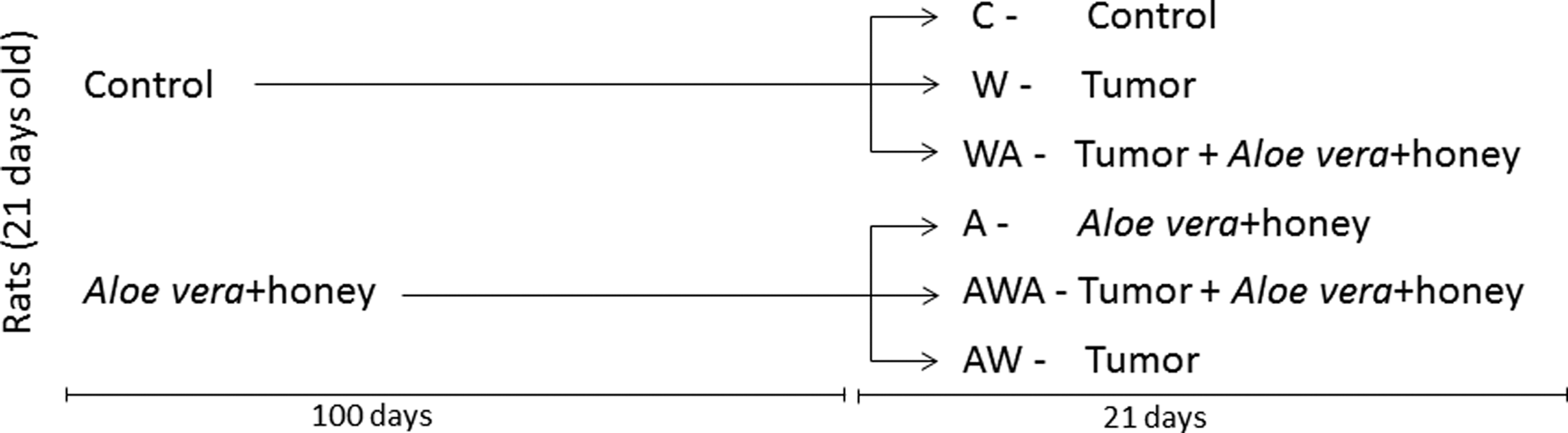
Diagram showing the organization of the experimental groups. C=control, W=tumor-control. WA=tumor-A. vera+honey, A=control-A. vera+honey, WA=A. vera+honey-tumor, and AWA=A. vera+honey-tumor-A. vera+honey.
During the entire experimental protocol, the animals were kept in an air-conditioned room with a 12-h light/12-h dark cycle, with free access to water and commercially available food. They were monitored for weight gain weekly and sacrificed 21 days after tumor induction. The tumors, livers, gastrocnemius muscles, hearts, and carcasses were collected for further analysis.
The general guidelines of the United Kingdom Coordinating Committee on Cancer Research for animal welfare were followed, 18 and the experimental protocols were approved by the Institutional Committee for Ethics in Animal Research (CEEA.IB/UNICAMP, protocol # 1400-1 and #2537-1).
Body composition analysis
The carcass (body without the tumor and gastrointestinal tract) was weighed and dried at 55°C until a constant weight was obtained. The carcass water was determined as the difference between the dry and wet weights. The carcass was homogenized, and a 10 g sample was used for fat extraction by petroleum ether; the total body fat was calculated as the difference between the initial sample weight (10 g) and the final sample weight (without fat) in proportion to the whole body weight. In addition, aliquots of the carcass after water and fat extraction were analyzed for total and collagen nitrogen content using a colorimetric micro-Kjeldahl method. 19 The noncollagen nitrogen was calculated by subtracting the collagen nitrogen from the total nitrogen content.
Oxidative stress and proteolysis evaluation
Tissue samples from the liver, heart, gastrocnemius muscle, and tumor were weighed, homogenized in PBS, and centrifuged for 15 min at 11,180 g. The supernatant was collected for biochemical analysis. The samples were maintained on ice and assayed in duplicate.
The total protein content of the samples, used to normalize the enzymatic activities, was determined by the Bradford method 20 using bovine serum albumin as a standard.
Superoxide dismutase (SOD) activity was assessed by measuring the degradation of horseradish peroxide and dimethylaniline in the presence of 4-aminofenazon and riboflavin following the method described by Tang et al., 21 and the data are expressed as arbitrary units*μg protein−1. Catalase activity was measured in the presence of hydrogen peroxide (final concentration 6 mM) as described by Cohen et al., 22 and the results are expressed in ηmol*min−1*μg protein−1. Glutathione-S-transferase (GST) activity (ηmol*min−1*μg protein−1) was determined following the conjugation of 1-chloro-2,4-dinitrobenzene (CDNB) with glutathione. 23 The malondialdehyde (MDA) content (ηmol*μg protein−1) was determined using the substrate n-methyl-2-phenylindole (MPO). 24 Once the GST activity and MDA content were determined, the MDA/GST ratio was calculated.
The chymotrypsin-like activity was determined using the fluorogenic substrate succinyl-Leu-Leu-Val-Try-7-amino-4-methylcoumarin (Suc LLVY-AMC; Calbiochem, Boston, MA, USA) in Tris-HCl, pH 7.4). The fluorescence (excitation: 360 ηm, emission: 460 ηm) was measured, and the results are expressed as arbitrary units*μg protein−1. 25
Calpain activity was measured by incubating the samples in a 50 mM imidazole-HCl buffer (pH 7.5) containing 10 mM β-mercaptoethanol, 1.0 mM NaN3, and 4 mg/mL casein. Subsequently, 5 mM CaCl2 was added to activate calpain to initiate the hydrolysis reaction and the absorbance (540ηm) was measured. 26 The results are expressed as arbitrary units*μg protein−1.
Alkaline phosphatase activity was measured using 37 mM 4-nitrophenyl disodium phosphate (p-NPP) 27 and expressed as ηmol*min−1*μg protein−1.
Statistical analysis
The results are expressed as the mean±SEM (standard error of mean). The comparisons among all groups were performed using one-way ANOVA followed by the Bonferroni test. When necessary, further analyses between selected groups were performed using an unpaired t-test. The P-values<.05 were considered statistically significant. Statistical analysis was performed using the GraphPad Prism software (v3.00 for Windows 98, Bethesda, MD, USA). 28
Results
The body parameter data are presented in Table 1. Positive weight gain was observed in the healthy rats (groups C and A), while the Walker 256 tumor-bearing groups displayed some degree of weight loss during tumor development (21 days). Remarkably, only rats from the W and AWA groups experienced significant weight reductions. In addition, significantly reduced tumor sizes were exclusively observed in the WA group.
P<.05 in comparison with the C group (one-way ANOVA and Bonferroni test).
Relative tumor weight (%) was calculated as [tumor weight (g)/final body weight (g)] *100.
Weight gain (%) was calculated as {[(final body weight (g)–tumor weight (g)]–body weight before tumor injection (g)/body weight before tumor injection (g)}*100.
Carcass relative weight (%) was calculated as [fresh carcass weight (body without the gastrointestinal tract and tumor) (g)/final body weight (g)]*100.
AH, A. vera and honey solution. C, control rats; W, tumor-bearing rats; WA, tumor-bearing rats administered AH after tumor injection; A, control treated with AH; AW, tumor-bearing rats administered AH before tumor injection; AWA, tumor-bearing rats administered AH before and after tumor injection.
In the assessment of body composition parameters (Table 1), the carcass water content increased in all of the Walker 256 tumor-bearing groups, whereas the fat content significantly decreased exclusively in the groups not treated with the A. vera and honey solution during tumor development (W and AW). In addition, the total body nitrogen content also tended to be decreased exclusively in untreated rats (W, P=.1344). The plasma albumin levels decreased in all tumor-bearing groups, but the reduction is only significant in the W and AWA groups (Table 1).
The analyses of the proteolytic enzymes calpain and chymotrypsin in the tumor and host tissues (gastrocnemius muscle, liver, and heart) are presented in Figure 2. Chymotrypsin activity significantly increased in the liver and tended to increase in the myocardial tissues of the untreated rats (W), whereas chymotrypsin activity is maintained in the groups treated with the A. vera and honey solution during tumor development (WA and AWA) (Fig. 2A–D). Moreover, calpain activity tended to be increased in the liver (P=.0735), gastrocnemius muscle (P=.0780), and tumor tissues (P=.0897) only in the untreated group (W) (Fig. 2E–H). In addition, alkaline phosphatase activity decreased in the cardiac tissues of the W, A, AW, and AWA rats, as well as the tumor tissues of the WA group (Fig. 2I–L).
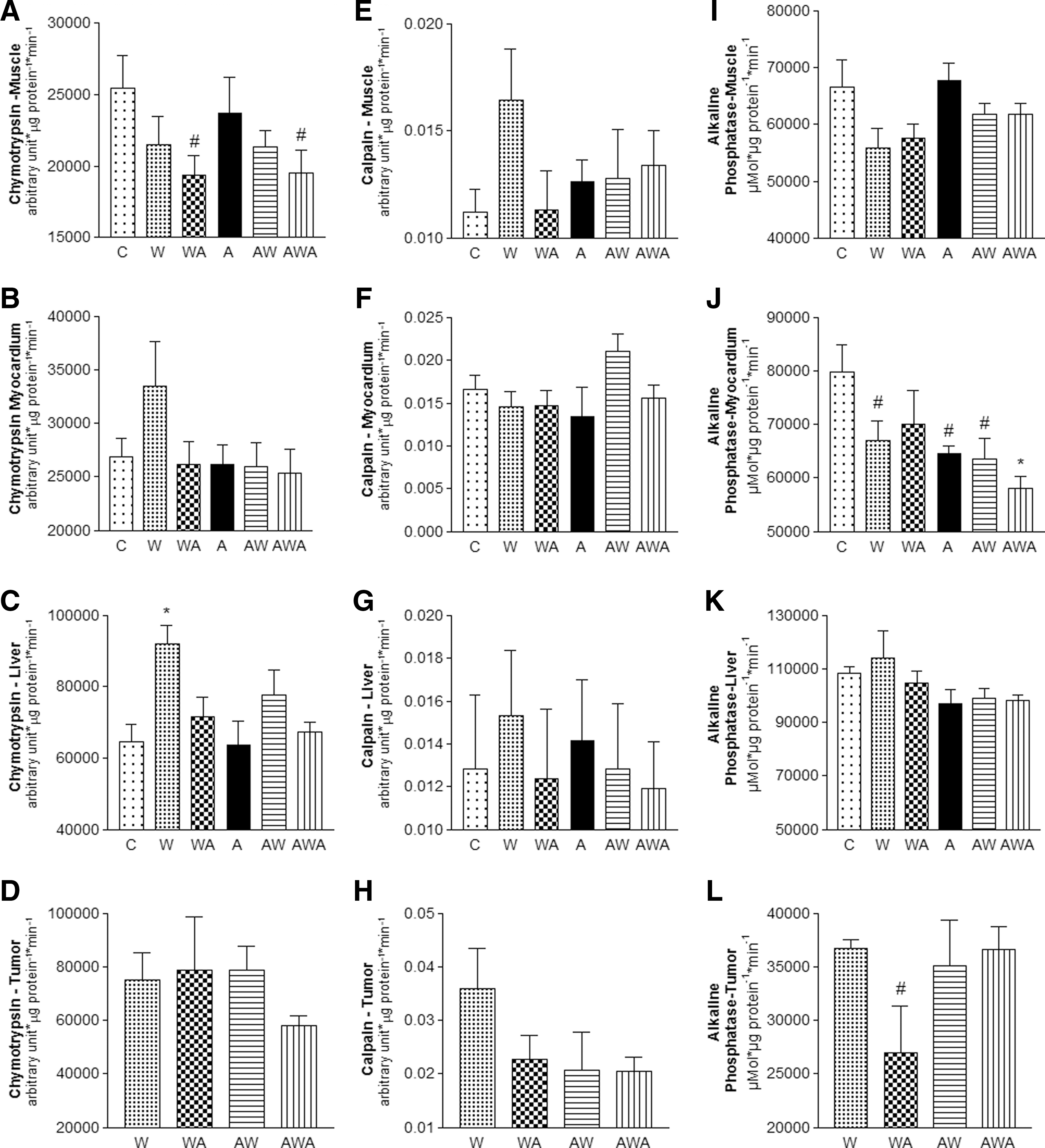
Chymotrypsin-like
The activities of the antioxidant enzymes SOD and catalase (Cat) as well as the MDA/GST ratios are presented in Figure 3. SOD activity decreased in liver tissues of the W, AW, and AWA groups as well as the muscles of the AWA group. Increased SOD activity was observed in the WA tumors (Fig. 3A–D). Catalase activity decreased in the myocardial tissues of the W and AWA rats and tended to progressively decrease in the tumors through the groups treated with the A. vera and honey solution (WA, AW, and AWA; respectively; Fig. 3E–H).
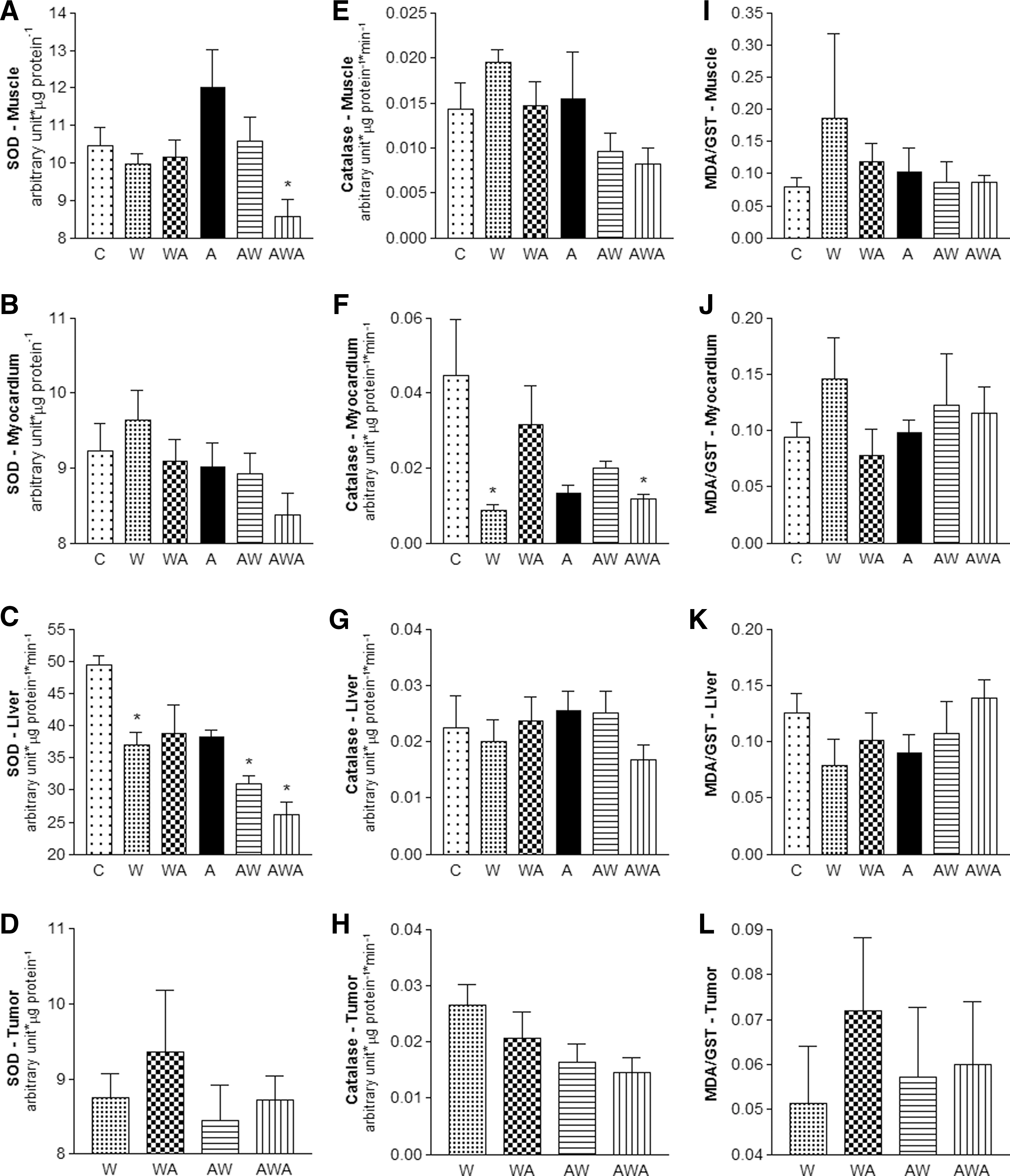
Superoxide dismutase
Finally, the MDA/GST ratio, an indicator of oxidative stress, tended to increase in the muscles (P=.0591) and hearts (P=.0752) of the untreated rats (W). In addition, this ratio tended to increase in the tumors of rats treated with the A. vera and honey solution after tumor induction (WA; P=.0698; Fig. 3I–L).
Discussion
One of the greatest challenges in cancer therapy is inducing tumor cell death without inflicting harm to the host. Complementary therapy studies are now being conducted, and this work shows that phytotherapies, such as A. vera and honey, have the potential to affect tumor development while minimizing cachectic processes in the host.
The Walker 256 carcinoma is a widely used cancer-cachexia model that mimics the human syndrome. According to data presented here, indeed tumor development leads to a significant decrease in weight gain, fat content, and carcass mass. These decreases were attenuated by the A. vera and honey solution, particularly when the solution was administered exclusively after tumor induction (WA group). During cancer-induced cachexia, albumin production is decreased and its degradation is enhanced. In fact, in this work, the albumin content decreased in all tumor-bearing groups, but this depletion was significant only in the W and AWA groups. In parallel, the water content also tended to be increased in all tumor-bearing groups, accompanying the decreased plasma albumin content, which is consistent with the fact that albumin maintains the colloid osmotic pressure.
Cachexia is also known to induce additional protein catabolism processes that first target the extracellular compartment and then extend to skeletal muscle cells, promoting atrophy and apoptosis. 4,29 Evidence for extracellular protein mobilization is observed in the tendency of decreased body collagen nitrogen content in the untreated Walker-256-bearing rats (W), not in the other treatment groups. This finding suggests that the A. vera and honey solution might be beneficial to the host.
The host wasting process observed during cancer-induced cachexia is complexly regulated by tumor cell release of proteolysis and lipolysis factors and specific immune system activation that results in chronic inflammation and increased reactive oxygen species in host tissues. 1,4 This process, mediated by tumor factors and cytokines, is capable of regulating not only the proteolysis/lipolysis machinery but also the cellular antioxidant system. 1,4
A. vera has been reported to contain various immunomodulatory compounds. 11,16 Therefore, we hypothesize that A. vera and honey therapy reduces chronic inflammation, oxidative stress, and catabolism through modulation of the activities of antioxidant enzymes, such as SOD and catalase. SOD characterizes the first defense against oxidative stress and catalyzes the dismutation of the superoxide anion (O2 −) into oxygen and hydrogen peroxide (H2O2), which is subsequently converted into water and oxygen by catalase. 30
In this work, numerous observations suggest that the A. vera and honey treatment may attenuate cancer-induced cachexia associated with skeletal muscle wasting. One example is the increase in calpain activity and MDA/GST levels, which were particularly apparent in untreated tumor-bearing rats (W), but not in A. vera- and honey-treated rats (especially those of the WA group). Given that the cysteine protease calpain integrates the calcium-dependent protein degradation pathway, which is known to be activated in muscles during cancer-induced cachexia 4,31 and involved in cell death, 32 the A. vera and honey solution potentially decreased host wasting processes through modulation of this pathway. In addition, the rats treated with A. vera and honey after tumor induction (WA and AWA) displayed a significant decrease in the chymotrypsin-like activity in the gastrocnemius muscle. The chymotrypsin-like activity corresponds to the catalytic core of the 20S subunit of the ubiquitin–proteasome pathway, which is considered the most important pathway for intracellular protein degradation under catabolic conditions. 33 Although we observed no increase in chymotrypsin-like activities in the muscle tissues of untreated tumor-bearing rats (W), these animals had alterations in other parameters consistent with a cachectic status, such as decreased body weight, decreased body fat and nitrogen content, increased chymotrypsin-like activity in the myocardial and liver tissues, and increased MDA/GST ratio in the gastrocnemius muscles.
MDA is a natural product from membrane phospholipid peroxidation. In high levels, MDA can bind to nucleic acids to form adducts, resulting in DNA mutations. 34 In contrast, GST is a phase II detoxification enzyme that is responsible for the conjugation of glutathione to MDA, promoting MDA removal; thus, a high MDA/GST ratio is an indicative of oxidative stress. 35 Our data suggest that A. vera and honey might modulate muscle wasting by reducing the chymotrypsin-like activity in the WA and AWA groups as well as simultaneously modulating antioxidant enzymes to lower the MDA/GST ratio, as observed in the WA, AW, and AWA groups relative to the untreated group (W).
Recently, cancer-induced cachexia has been reported to cause cardiac alterations and atrophy in mouse models. 36,37 Our experiments in rats also suggest that heart damage occurs, as we observed a tendency toward an increase in chymotrypsin-like activity and oxidative stress in the untreated Walker-256-bearing rats (W group). The observed tendency of increase in the MDA/GST levels, which was particularly apparent in the W group, could result from an imbalance in SOD and catalase activities. The combination of increased SOD activity and significantly decreased catalase activity may result in hydrogen peroxide accumulation and MDA production. The SOD and catalase activities in cardiac tissues of the rats treated with the A. vera and honey solution after tumor induction (WA) were maintained, leading to a balanced MDA/GST ratio.
In addition to muscular, adipose, and cardiac tissues, cachexia also severely affects the liver by disrupting of the homeostatic balance between anabolism and catabolism to promote persistent catabolic status corroborating to the cachectic process and/or by promoting the same alterations observed in other host tissues (e.g., oxidative stress, catabolism, and cell death). 38 In the present study, chymotrypsin-like activity increased and calpain activity tended to increase in the livers of the untreated tumor-bearing rats (W); however, these changes were not present or significant in the A. vera- and honey-treated rats (WA, AW, and AWA). Altogether, these results suggest a protective role for A. vera and honey treatment in hepatic tissues, particularly in the WA group. Indeed, A. vera has been reported to have hepatoprotective activity. 39
Several studies have demonstrated that both A. vera and honey possess antioxidant properties, and synergism of some of their compounds cannot be discounted. 11 A. vera specifically activates SOD, catalase, and phase II enzymes (including GST) in organs such as the liver, lungs, kidney, and forestomach. 12 Honey flavonoids and phenolic compounds increase the serum antioxidant capacity in healthy human subjects 40 and increase antioxidant enzyme activities in mice. 41
Although several data suggest that the A. vera and honey solution is beneficial to the host, the observed results are likely indirect. The majority of the benefits were observed in the WA group, and this group presented the smallest tumors. The production of tumor factors that promote development of the cancer cachexia syndrome would be compromised in tumors of reduced volume, thereby resulting in a reduction in the general host wasting process.
In fact, analyzing the tumor tissue itself, we noticed a tendency of decrease in calpain activity in the tumors of the A. vera- and honey-treated groups (WA, AW, and AWA) compared with the untreated group (W), suggesting that the A. vera and honey solution modulates calpain's protumorigenic activities. Calpain expression is often increased in aggressive tumors and linked to cell migration and survival. 42 In addition, reduced alkaline phosphatase activity is observed in the WA tumors, suggesting decreased cell activity. In this case, reduced cell activity could be indicative of cell death or senescence, both of which were previously reported to occur in response to A. vera or honey compounds. 17,43 Importantly, the WA tumors, which were the smallest on average among the groups, displayed a tendency of increase in SOD activity and no alterations in catalase activity. The subsequent increase in oxidative stress (MDA/GST ratio) is consistent with the notion that the A. vera and honey treatment leads to oxidative stress and cell death in cancer cells. A previous study reported that an increase in SOD activity without a concomitant increase in catalase activity in cancer cells (as observed in the WA tumors) results in oxidative stress through hydrogen peroxide accumulation. 5 These observed effects are in agreement with a study that treated lymphoma-bearing mice with emodin, an active compound found in A. vera. 44
Indeed, the effects observed in the WA-treated tumors are similar to those proposed by the oxidative therapy, which comprises various strategies to increase oxidative stress to lethal levels in tumor cells. 11,45,46 These strategies include the inhibition of antioxidant enzymes, such as SOD, catalase, and GST, to promote the accumulation of reactive oxygen species, which leads to cell damage and cell death. 47 Such approaches appear to be very promising in cancer therapy and their application has recently increased. 46
Thus, the achievements of this work provide evidence that the A. vera and honey solution decreased host wasting and also increased oxidative stress in tumor cells, particularly when administered therapeutically (immediately after tumor induction, group WA), by differentially modulating antioxidant and proteolysis enzymes in the host and tumor tissues. The mechanisms responsible for these effects might include the deleterious actions of various A. vera and honey compounds (aloin, aloe-emodin, acemannan, and flavonoids) on tumor cells, which might result in indirect effects on the host tissues. These mechanisms may also include immune system modulation as well as altering the chronic proinflammatory status of cancer-induced cachexia. Certainly, several compounds in A. vera and honey synergized to generate the observed effects in this model, and our group is still working to elucidate both the compounds and the mechanisms involved in A. vera- and honey-mediated modulation of the cancer cachexia syndrome and tumor growth.
Footnotes
Acknowledgments
The authors are grateful for the financial support of FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo, #2010/00209-9; #2013/16115-1) and CNPq (Conselho Nacional de Desenvolvimento Cientifico e Tecnológico; #304604/2010-0; #302863/2013-3). R. Tomasin, MSc, was supported by a scholarship sponsored by FAPESP (Grant #2007/05788-4 and #2010/00714-5). The article was edited for correct English language by native English speaking editors at American Journal Experts (Certificate number 8EF3-A3CE-B828-1C26-82F8).
Author Disclosure Statement
The authors declare that there are no conflicts of interest.