
Abstract
We aimed in the present study to investigate the hepato- and nephroprotective effects of Lavandula stoechas essential oils (LSEO) against malathion-induced oxidative stress in young male mice as well as the possible mechanism implicated in such protection. Animals were divided into eight groups of 12 each: Control, malathion (200 mg/kg b.w.); Various doses of LSEO (10, 30, and 50 mg/kg b.w.), malathion+various doses of LSEO. Malathion and LSEO were daily per orally (p.o.) administered by intragastric gavage during 30 days. We initially found that malathion treatment induced body weight gain decrease as well as a clear nephro- and hepatotoxicity as assessed by significant relative liver and kidney weight increase and related hemodynamic parameters deregulation. Malathion exposure of mice also induced a considerable perturbation of metabolic parameters. On the other hand, we showed that malathion administration was accompanied by an oxidative stress status assessed by an increase of malondialdehyde (MDA) and hydrogen peroxide (H2O2) levels as well as a depletion of sulfhydril group content (-SH) and antioxidant enzyme activities such as catalase (CAT) and glutathione peroxidase (GPx), total superoxide dismutase (SOD), Cu/Zn-SOD, Mn-SOD, and Fe-SOD in the kidney and liver. More importantly, LSEO treatment abolished all malathion-induced body gain loss, liver and kidney relative weight increase, hemodynamic and metabolic disorders, as well as hepatic and renal oxidative stress. In conclusion, our data suggest that LSEO exerted potential hepato- and nephroprotective effects against malathion-induced oxidative stress in mice. The beneficial effect of LSEO might be related, in part, to its antioxidant properties.
Introduction
O
The cellular antioxidant status determines the susceptibility to oxidative damage and is usually altered in response to oxidative stress. The cellular antioxidant action is reinforced by the presence of dietary antioxidants. 13 Accordingly, interest has recently grown in the role and usage of natural antioxidants as strategy to prevent against oxidative damage in various health disorders, with oxidative stress as a factor in their pathophysiology. 13 Natural antioxidants from fruits and vegetables are reported to provide substantial protection that slows down the process of oxidative damage caused by ROS. 14 The beneficial effect of polyphenols is associated with a multitude of biological activities, including antioxidant and free radical-scavenging properties, antiplatelet aggregation, and inhibition of vascular smooth muscle cell proliferation. All these effects might interfere with atherosclerotic plaque development and stability. These observations might explain their cardiovascular protective properties. 15 On the other hand, it is now established that hyperlipidemia represents a major risk factor for the premature development of atherosclerosis and its cardiovascular complications. 16 A logical strategy to prevent against atherosclerosis and reduce the incidence of cardiovascular disease events is to target the hyperlipidemia and oxidative stress by diet and/or drug intervention.
Lavender (Lavandula stoechas L.) is a medicinal plant largely used and included for centuries in the pharmacopoeia of several countries, including Tunisia. The phytochemical screening of this plant revealed that it is rich in bioactive molecules, such as phenolic compounds present a high scavenging capacity. 17 Therefore, owing mainly to their antioxidant 18,19 and anti-inflammatory 20 properties, lavender extracts or its essential oils are known to exhibit antidiabetic, 21 neuroprotective, 22 antiallergic, 23 antimicrobial, 24 and anticancer 25 activities. Recently, we have shown that L. stoechas essential oils (LSEO) protect against reproductive damage and oxidative stress in alloxan-induced diabetic rats. 26
Hence, the present study aimed to investigate the hepato- and nephroprotective effects of LSEO against malathion-induced oxidative stress in young male mice as well as the possible mechanism implicated in such protection.
Materials and Methods
Chemicals
Malathion (96%), epinephrine, bovine catalase, 2-thio-barbituric acid (TBA), and butylated hydroxytoluene (BHT) were purchased from Sigma Chemical Co. (Sigma-Aldrich GmbH, Steinheim, Germany). All other chemicals used were of analytical reagent grade.
Plant collection
L. stoechas aerial parts were collected in March 2010 from the area of Ain-Draham (North-West of Tunisia) and identified by the laboratory of taxonomy in the Faculty of Sciences of Tunis (FST)-Tunisia. The voucher specimens (No. L101) have been deposited with the herbarium of the Higher Institute of Biotechnology of Béja and also in the Department of Biological Sciences, Faculty of Science of Tunis, Tunisia.
Plant extracts preparation
The dried aerial parts were submitted to hydrodistillation for 3 h using a Clevenger-type apparatus. Briefly, the plant was immersed in water and heated to boiling, after which the essential oils were evaporated together with water vapour and finally collected in a condenser. The distillate was isolated and dried over anhydrous sodium sulfate.
Animals and treatment
Healthy male Swiss Albino mice (Mus musculus) (weighing 10±0.3 g, 3 weeks old, and housed six per cage) were purchased from SIPHAT (Tunis, Tunisia) and used in accordance with the Local Ethics Committee of Tunis University for use and care of animals in conformity with the NIH recommendations. The animals were housed in standard cages (40×28×16 cm) under controlled conditions: 12-h light–12-h dark, 20–22°C, food (BADR, Utique, Tunisia) and water are ad libitum. Mice were divided into eight groups of 12 animals each: control, malathion (200 mg/kg b.w.); various doses of LSEO (10, 30, and 50 mg/kg b.w.), malathion+various doses of LSEO. The animals were daily treated by gavage during 30 days with vehicle (control, NaCl 0.9%) or with various doses of LSEO (injection volume was 1 mL/kg b.w.). After 1 hour, mice also received malathion in corn oil (200 mg/kg b.w.) by gavage during 30 days.
Group I: served as a negative control and received NaCl (0.9%) and corn oil by gavage during 30 days.
Group II: served as a positive control and received NaCl (0.9%) and malathion (200 mg/kg b.w.) by gavage during 30 days.
Groups III–V: received various doses of LSEO (10, 30, and 50 mg/kg b.w.) and corn oil during 30 days.
Groups VI–VIII: received various doses of LSEO (10, 30, and 50 mg/kg b.w.) and malathion (200 mg/kg b.w.) during 30 days.
Twenty-four hours after the last injection, animals were sacrificed and blood was collected in heparinized tubes. After centrifugation at 3000 g for 15 min, plasma was processed for biochemical parameter determinations. The liver and kidney were rapidly excised and homogenized in phosphate-buffered saline (PBS). After centrifugation at 10,000 g for 10 min at 4°C, supernatants were used for biochemical determination.
Body and reproductive organ weights
Initial (weight at start point) and final (weight at end point) body weights were recorded. After decapitation, laparotomy was performed. The liver and kidney were stripped from fatty tissues and blood vessels, blotted, and their absolute weights were determined using precision balance Denver S-603 series. To normalize the data for statistical analysis, organ weights were expressed per 100 g body weight.
Functional and metabolic parameters
Assessment of liver function
Plasma albumin, bilirubin, aspartate aminotransferase (AST), alanine aminotransferase (ALT), phosphatase alcaline (PAL), acid phosphatase (ACP), lactate dehydrogenase (LDH), and gamma-glutamyl transferase (γ-GT) were measured using commercially available diagnostic kits (Biomaghreb, Ariana, Tunisia).
Assessment of renal function
Plasma urea, creatinine, and uric acid analyses were performed using commercially available diagnostic kits (Biomaghreb).
Metabolic parameters
Plasma total lipids (TL), total cholesterol (TC), low-density lipoprotein-cholesterol (LDL), high-density lipoprotein-cholesterol (HDL), and triglyceride (TG) concentrations were measured using commercially available diagnostic kits supplied by Randox Laboratories (Ardmore, Northern Ireland, United Kingdom).
Lipid peroxidation
Lipid peroxidation was detected by the determination of malondialdehyde (MDA) production by the method of Buege and Aust. 27 Briefly, homogenates of the liver and kidney were centrifuged at 1000 g for 10 min at 4°C to sediment cell debris. Supernatants were suspended in PBS, pH=7.4, mixed with BHT-TCA solution (1‰ BHT dissolved in 0.2 g/mL TCA), centrifuged at 1000 g for 35 min, and finally, mixed with 0.5 N HCl and 120 mM TBA (thiobarbituric acid) in 26 mM Tris and heated in a water bath at 80°C for 10 min. After cooling, the absorbance of the resulting chromophore was measured at 532 nm. MDA levels were determined by using an extinction coefficient for the MDA-TBA complex of 1.56×105 M−1cm−1. The results were expressed as nmol of malondialdehyde per milligram of protein (nmol MDA/mg protein).
Thiol groups measurement
Total concentration of thiol groups (-SH) was performed according to the Ellman's method. 28 Briefly, aliquots from the liver and kidney were mixed with 100 μL of 10% SDS and 800 μL of 10 mM phosphate buffer (pH 8) and the absorbance was measured at 412 nm (A0). Then, 100 μL of DTNB were added and incubated at 37°C during 60 min. After incubation, the absorbance of the sample was measured at 412 nm (A1). The concentration of thiol groups was calculated from A1−A0 subtraction using a molar extinction coefficient of 13.6×103 M−1cm−1. Results were expressed as nmol of thiol groups per mg of protein.
Antioxidant enzyme activity assays
Liver and kidney SOD activities were determined by using a modified epinephrine assay of Misra and Fridovich. 29 At alkaline pH, superoxide anion O2 − causes the autoxidation of epinephrine to adenochrome; while competing with this reaction, SOD decreased the adenochrome formation. One unit of SOD is defined as the amount of the extract that inhibits the rate of adenochrome formation by 50%. The enzyme extract was added in a 2 mL reaction mixture containing 10 μL of bovine catalase (0.4 U/μL), 20 μL of epinephrine (5 mg/mL), and 62.5 mM sodium carbonate/bicarbonate buffer pH 10.2. Changes in absorbance were recorded at 480 nm. Characterization of SOD isoforms was performed using KCN (2 mM), which inhibits Cu/Zn-SOD or H2O2 (5 mM) and affecting both Cu/Zn-SOD and Fe-SOD, whereas Mn-SOD was insensitive to both inhibitors. 30
Liver and kidney CAT activities were assayed by measuring the initial rate of H2O2 disappearance at 240 nm. 31 The reaction mixture contained 33 mM H2O2 in 50 mM phosphate buffer pH 7.0, and the CAT activity was calculated using the extinction coefficient of 40 mM−1cm−1 for H2O2.
The GPx activity was measured by the procedure of Flohé and Günzler. 32 Briefly, 1 mL of reaction mixture containing 0.2 mL of sample supernatant (liver or kidney), 0.2 mL of phosphate buffer 0.1 M pH 7.4, 0.2 mL of GSH (4 mM), and 0.4 mL of H2O2 (5 mM) was incubated at 37°C for 1 min and the reaction stopped by addition of 0.5 mL TCA (0.05 g/mL). After centrifugation at 1500 g for 5 min, an aliquot (0.2 mL) from the supernatant was mixed with 0.5 mL of phosphate buffer 0.1 M pH 7.4 and 0.5 mL DTNB (10 mM) and absorbance recorded at 412 nm. The GPx activity was expressed as nmol of GSH consumed/min/mg protein.
H2O2 determination
The liver and kidney H2O2 levels were performed according to Dingeon et al. 33 Briefly, the hydrogen peroxide reacts with p-hydroxybenzoic acid and 4-aminoantipyrine in the presence of peroxidase leading to the formation of quinoneimine, which has a pink color detected at 505 nm.
Protein determination
The protein concentration was determined according to the Bradford method. 34 Serum albumin was used as a standard.
Statistical analysis
The data were analyzed by one-way analysis of variance (ANOVA) and expressed as mean±standard error of the mean (SEM). The data are representative of 12 independent experiments. All statistical tests were two-tailed, and a P value <0.05 was considered significant.
Results
Mortality and macroscopic symptoms of toxicity
No mortality was noted until the end of the experiment in control and treated groups. Concerning the liver and kidney, no macroscopic alterations were found. However, cholinergic signs of adverse toxicological effects were observed such as sluggishness, muscular tremors, irregular movements, and abdominal tremble in the malathion-treated group. LSEO administration reversed all adverse cholinergic effects induced by malathion intoxication.
Body and relative organ weights
Data from Table 1 showed that malathion-treated mice showed a significant decrease of body weight gain. Malathion intoxication also increased the relative weights of the liver and kidney by 19% and 27%, respectively. LSEO administration significantly and dose dependently protected against malathion-induced body weight decrease as well as liver and kidney relative weights increase.
Animals were treated during 30 days with LSEO (10, 30, and 50 mg/kg b.w., p.o.). One hour after essential oils administration, mice were daily exposed to malathion (200 mg/kg b.w., p.o.) or corn oil for the same period. Assays were carried out in triplicate (n=12).
P<.05 compared to control group (NaCl+corn oil).
P<.05 compared to malathion group (NaCl+malathion).
LSEO, Lavandula stoechas essential oils.
Effects of malathion and LSEO on liver and kidney functions
To test the in vivo effect of LSEO on malathion-induced liver injury, mice were treated with various doses of LSEO and malathion during 30 days. As shown in Table 2, malathion-induced hepatotoxicity was shown by a decrease of plasma albumin content as well as a significant increase of plasma bilirubin, AST, ALT, ALP, ACP, LDH, and γ-GT levels. Interestingly, when animals were coadministrated with LSEO, the levels of these parameters were practically restored in a dose-dependent manner.
Animals were treated during 30 days with LSEO (10, 30, and 50 mg/kg b.w., p.o.). One hour after essential oils administration, mice were daily exposed to malathion (200 mg/kg b.w., p.o.) or corn oil for the same period. Assays were carried out in triplicate (n=12).
P<.05 compared to control group (NaCl+corn oil).
P<.05 compared to malathion group (NaCl+malathion).
AST, aspartate aminotransferase; ALT, alanine aminotransferase; PAL, phosphatase alcaline; ACP, acid phosphatase; LDH, lactate dehydrogenase; γ-GT, gamma-glutamyl transferase.
We also studied in the present work, the effect of LSEO on malathion-induced renal dysfunction (Table 3). Malathion per se increased plasma creatinine and urea levels when the uric acid content decreased significantly. However, the LSEO treatment significantly and dose dependently protects against malathion-induced renal hemodynamic parameters deregulation.
Animals were treated during 30 days with LSEO (10, 30, and 50 mg/kg b.w., p.o.). One hour after essential oils administration, mice were daily exposed to malathion (200 mg/kg b.w., p.o.) or corn oil for the same period. Assays were carried out in triplicate (n=12).
P<.05 compared to control group (NaCl+corn oil).
P<.05 compared to malathion group (NaCl+malathion).
Effects of malathion and LSEO on lipid metabolic parameters
Table 4 showed that malathion exposure induced a considerable metabolic disorder, as assessed by a significant increase of TL, TG, TC, and LDL plasma contents as well as a decreased plasma HDL level. Importantly, we showed that LSEO supplementation significantly protects against the disturbance of lipid metabolic parameters induced by malathion intoxication.
Animals were treated during 30 days with LSEO (10, 30, and 50 mg/kg b.w., p.o.). One hour after essential oils administration, mice were daily exposed to malathion (200 mg/kg b.w., p.o.) or corn oil for the same period. Assays were carried out in triplicate (n=12).
P<.05 compared to control group (NaCl+corn oil).
P<.05 compared to malathion group (NaCl+malathion).
TL, total lipids; TC, total cholesterol; LDL, low-density lipoprotein-cholesterol; HDL, high-density lipoprotein-cholesterol; TG, triglyceride.
Effects of malathion and LSEO on lipid peroxidation and thiol groups content
We further looked at the effect of malathion and essential oils on oxidative stress conditions. We initially showed that oxidative stress induced by malathion treatment is accompanied by a significant increase of liver and kidney MDA, hydrogen peroxide, as well as a decrease of thiol groups content (Fig. 1). Treatment with LSEO protected against lipoperoxidation, H2O2 increase, as well as (-SH) groups decrease induced by malathion intoxication.

Subchronic effects of Lavandula stoechas essential oils (LSEO) and malathion on malondialdehyde (MDA)
Effects of malathion and LSEO on antioxidant enzyme activities
We also studied the hepatic and renal antioxidant enzyme activities (Figs. 2 and 3). As expected, malathion per se significantly decreased CAT, GPx, total SOD, Cu/Zn-SOD, and Mn-SOD activities. The subchronic administration of LSEO protected against the depletion of hepatic and renal antioxidant enzyme activities induced by malathion intoxication in a dose-dependent manner. ROEO alone had no antioxidant enzyme activities.

Subchronic effects of LSEO and malathion on CAT
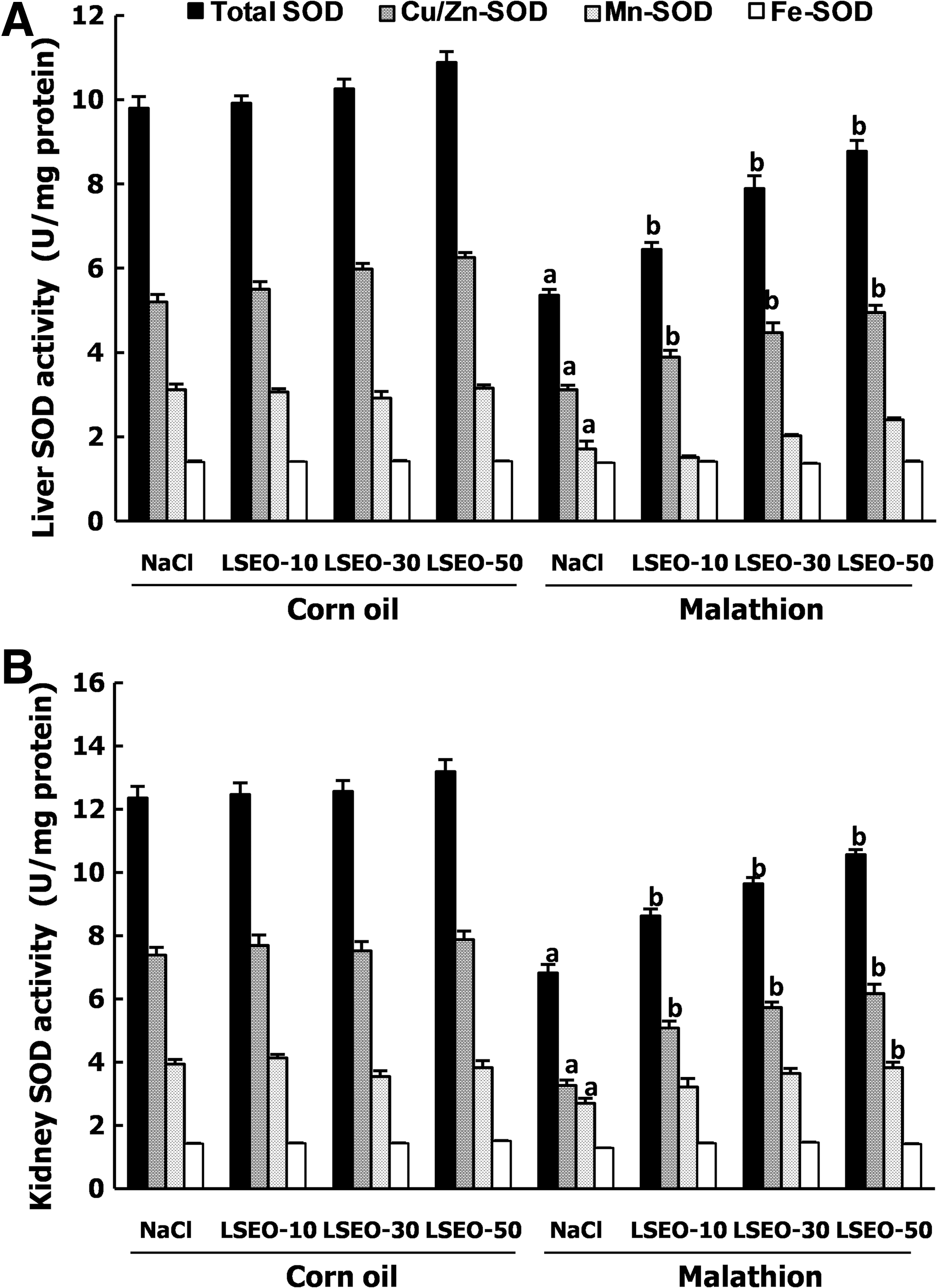
Subchronic effects of LSEO and malathion on total-SOD, Cu/Zn-SOD, Mn-SOD, and Fe-SOD activities in the liver (
Effects of malathion and LSEO on antioxidant enzyme activities
The effects of malathion and LSE on hydrogen peroxide level are reported in Figure 4. We showed that malathion intoxication significantly increased the liver and kidney H2O2 levels. LSEO administration significantly protected against malathion-induced hydrogen peroxide increase.
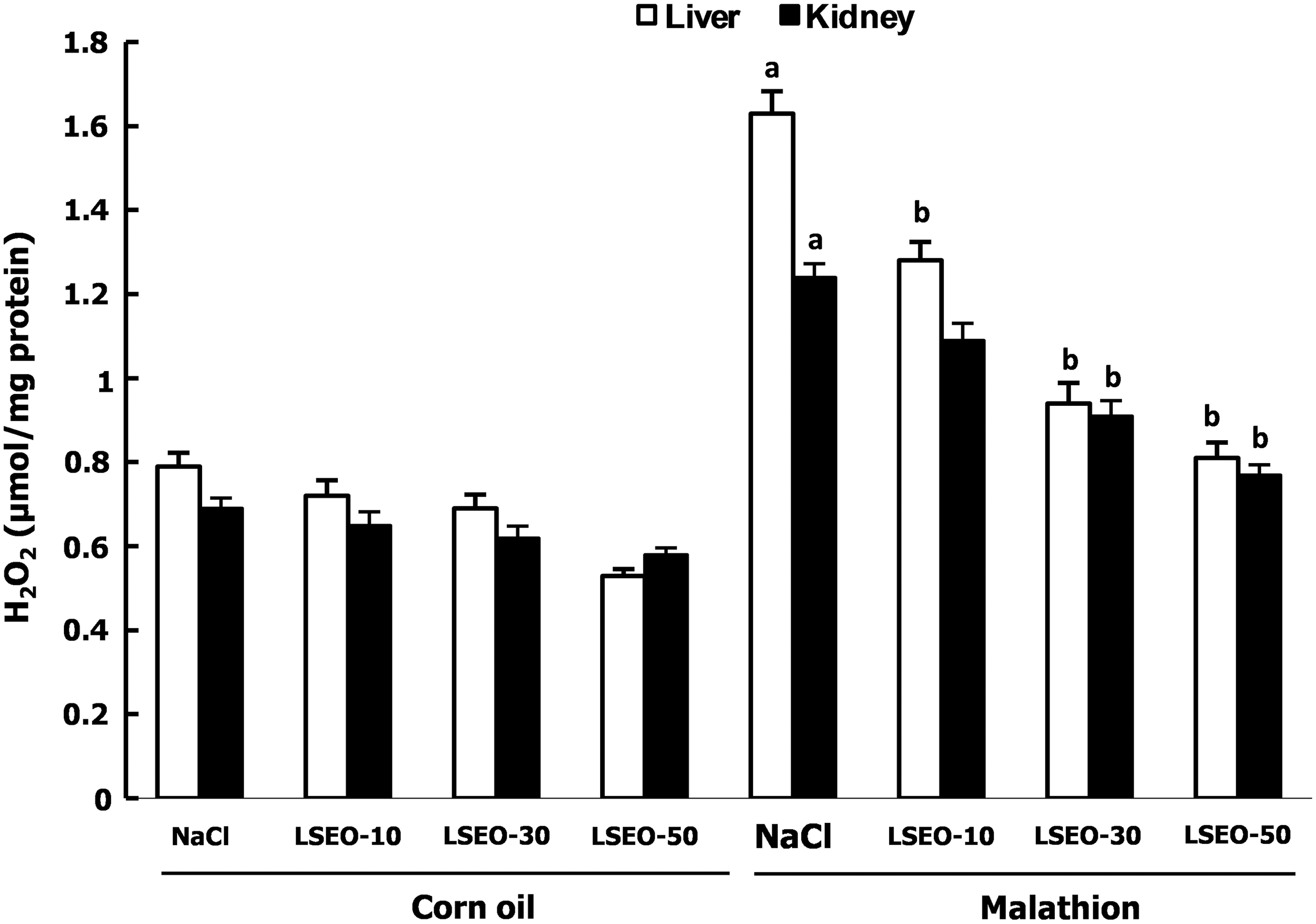
Subchronic effects of LSEO and malathion on hydrogen peroxide level in the liver and kidney of young male mice. Animals were treated during 30 days with LSEO (10, 30, and 50 mg/kg b.w., p.o.). One hour after essential oils administration, mice were daily exposed to malathion (200 mg/kg b.w., p.o.) or corn oil for the same period. Assays were carried out in triplicate (n=12). a P<.05 compared to control group (NaCl+corn oil) and b P<.05 compared to malathion group (NaCl+malathion).
Discussion
In the present work, we studied the protective role of LSEO against malathion-induced injury in the liver and kidney of young male mice as well as the implication of oxidative stress in such protection.
We initially revealed that malathion exposure (200 mg/kg b.w.) of young male mice during 30 induced a significant decrease of body weight as well as an increase of liver and kidney relative weights. The increased relative weights of liver and kidney were also observed with several cases of OP poisoning, in mice, such as chlorfenvinfos, fenthion, and dimethoate. 35 This loss of corporal mass has consequently numerous toxic effects in male mice, such as their feeding ability, and therefore, their reproductive performances. 36 LSEO coadministration protects in a dose-dependent manner against malathion-induced body and relative organ weights deregulation. We also showed in the present work that malathion treatment resulted in clear hepatotoxicity as evidenced by a decrease of plasma albumin content as well as a significant increase of plasma bilirubin, AST, ALT, ALP, ACP, LDH, and γ-GT levels, used as indexes of liver injury. On the other hand, malathion administration also induced renal dysfunction as assessed by an increase of plasma creatinine and urea levels when uric acid content decreased significantly. However, LSEO treatment significantly and dose dependently protects against malathion-induced liver injury and renal hemodynamic parameters deregulation. The high level of ALT and AST in the plasma compartment is a central indication of the degree of damage to the liver caused by OP administration. 37,38 We suggest that LSEO prevent against Kupffer cell activation by decreasing the formation of inflammatory and fibrogenic mediators. The same mechanism was, in part, proposed by Zhong et al. 39 to explain the protective effect of polyphenols extracted from Camellia sinensis against cholestasis-induced liver fibrosis in rats.
Our results also showed that malathion exposure induced an increase in serum HDL as well as a decrease of TL, TC, TG, and LDL levels when compared to the control group. LSEO coadministration protected against all malathion-induced metabolic lipid parameters deregulation. HDL may hasten the removal of cholesterol from the peripheral tissue to the liver for catabolism and excretion. Also, the high level of HDL may compete with LDL receptor sites on arterial smooth muscle cells and thus partially inhibit the uptake and degradation of LDL. Also, HDL could protect LDL against oxidation in vivo, because the lipids in HDL are preferentially oxidized before those in LDL. 40
Malathion-induced hepato- and nephrotoxicity was also assessed by an increase in lipoperoxidation, a decrease in SH- group levels, as well as depletion of antioxidant enzyme activities such as CAT, GPx, total SOD, Cu/Zn-SOD, Mn-SOD, and Fe-SOD. Malathion-induced tissue oxidative stress was widely documented in many organ systems such as the liver, 3 kidney, 3 heart, 41 brain, 9 and erythrocytes. 9 However, OP exposure can lead to oxidative stress through the generation of ROS 42 produced by cellular metabolic activities leading to the reduction of the cellular antioxidant defence capacity. 6 Hydroxyl radical (OH•) reacts with all biological substances and the most susceptible ones are polyunsaturated fatty acids. Reaction with these cell membrane constituents leads to lipid peroxidation. 43 Increased LPO impairs membrane function by decreasing membrane fluidity and changing the activity of membrane-bound enzymes and receptors. 43
More importantly, LSEO treatment protected against malathion-induced hepatic and renal oxidative stress. These data fully corroborated all previously reported in vivo and in vitro antioxidant and anti-inflammatory properties of L. stoechas. 21 Malathion-induced oxidative stress as well as liver and kidney dysfunctions have been shown to be attenuated by N-acetylcysteine, 44 vitamin C, and vitamin E. 45 However, to the best our knowledge, our report is the first one to deal with the LSEO protective effect on malathion-induced oxidative stress in the liver and kidney of young mice.
The chemical composition of LSEO using the GC-MS technique has been done in our previous report, which allowed the identification of 22 compounds.
21
The principal compounds detected are as follows:
Conclusion
Our data clearly demonstrate that LSEO exert protective effects against malathion-induced injury in the rat liver and kidney owing, in part, to their antioxidant properties.
Footnotes
Acknowledgment
The financial support of the Tunisian Ministry of Higher Education and Scientific Research is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.