
Abstract
The chronic ingestion of raw or undercooked kidney beans (Phaseolus vulgaris L.) causes functional and morphological derangement in various tissues. The major objectives of this study were to investigate the gavage effects of a raw Beldia bean variety that is widely consumed in Tunisia, on the small intestine morphology and jejunal absorption of water, electrolytes, and glucose in Wistar rats. Twenty young male rats were randomly divided into two groups of 10 rats. The first group served as the control and was gavaged with 300 mg of a rodent pellet flour suspension (RPFS), whereas the second experimental group was challenged with 300 mg of a Beldia bean flour suspension (BBFS) for 10 days. Histological studies were performed using light and electron microcopy. The intestinal transport of water, sodium, potassium, and glucose was studied by perfusing the jejunal loops of the small bowels in vivo. The feeding experiments indicated that BBFS did not affect weight gain. Histomorphometric analyses showed that the villus heights, crypt depths, and crypt/villus ratios in the jejunum and ileum were greater in the BBFS-fed rats than controls. Electron microscopy studies demonstrated that the rats exposed to RPFS exhibited intact intestinal tracts; however, the BBFS-treated rats demonstrated intestinal alterations characterized by abnormal microvillus architectures, with short and dense or long and slender features, in addition to the sparse presence of vesicles near the brush border membrane. BBFS administration did not significantly affect glucose absorption. However, significant decreases were observed in water and electrolyte absorption compared with the uptake of the controls. In conclusion, raw Beldia beans distorted jejunum morphology and disturbed hydroelectrolytic flux.
Introduction
M
Feeding growing animals diets containing raw beans as a main source of protein engenders numerous undesirable physiological and biochemical effects. These effects typically include growth inhibition, poor nitrogen balance, reduced intestinal absorption of sugars and amino acids, and impaired immune response. 8 –10 Ingested PHA, which is highly resistant to gut proteases, binds to the brush border cells of the intestine where it is rapidly endocytosed. When P. vulgaris lectin enters cells, it induces enhanced metabolic activity that eventually leads to hyperplasia and hypertrophy of the small intestine. 11 These processes might provoke potentially damaging reactions in the gastrointestinal track with powerful local or systemic effects. The toxic compound PHA is present in many common bean varieties, but it is especially concentrated in red and white kidney beans. 12
White kidney beans are an important Tunisian agricultural crop, particularly the Twila, Beldia, and Coco varieties (INRAT, Tunis, Tunisia). White beans are the most important source of energy, proteins, and vitamins for many Tunisian families. These seeds are also extremely versatile. They can be incorporated into soups and stews; or decorticated, ground into flour, and steamed or pied to make the bakery products; featured as traditional foods in Tunisia. They can also be found in fresh salads and purchased as dried, canned, or cooked. Despite the potential for beans to improve the diets of poor people across the world, one major hurdle exists regarding their optimal use as a food; they include lectins that cause low digestibility and abdominal upset.
Food poisoning arising from bean lectins, otherwise known as PHAs, has not been properly addressed in either Tunisia, or most parts of the developing world, that depend on/consume mostly vegetable-based diets. The abundance of these antinutritional factors and the toxic natures of the beans used to feed humans and animals certainly raise concerns. One might argue that because traditional Tunisian food preparation involves cooking, no cause for alarm exists; however, this case is not entirely correct because although the toxic effects of most of the lectins that are present in beans are generally eliminated by proper cooking, they might not be completely destroyed. For example, Korte 13 observed that in mixtures of ground beans and ground cereals prepared under the field conditions common in Africa, hemagglutinin was not always completely destroyed, and that cooked products produced diarrhea and other signs of toxicity.
The reduction in the boiling point of water in mountainous regions might also lead to the incomplete destruction of toxicity or the incomplete elimination of phytolectins. This problem is particularly serious with regard to seed lectins because they are resistant to cooking, and certain seeds; such as kidney beans, must be heated for several hours at temperatures above 80°C or boiled for 10–20 min to fully eliminate their lectin activity. 7,14 In addition, people do not patiently prepare or process beans to the recommended level to destroy the PHAs because of the high and prohibitive cost of energy sources, such as kerosene and gas, and the scarcity of firewood, especially during rainy seasons. 15,16 As a result, people are forced to eat improperly cooked nutritionally toxic beans. 17 The press has also reported instances of food poisoning leading to mortality in humans after the consumption of bean meal.
Tunisian public health authorities and other food regulatory bodies must be informed about the possible dangers related to PHAs. Thus, several coordinated pulse-improvement programs have been developed in Tunisia; including food safety programs geared toward ensuring the safety of beans for consumers.
The most recent work that was obtained, through a collaborative program between the Department of Animal Resources, Fisheries, and Food Technology, the National Institute of Agronomy of Tunisia (INAT), and the Intestinal Immunophysiology-Research Unit, Faculty of Medicine of Tunis (FMT), revealed that the white kidney bean variety Beldia contains a significant amount of lectins (∼9.20 g/kg) and has high hemagglutinating activity (HA ∼1/512). 18 Immunochemical analyses of seed proteins have indicated that abundant PHAs that cross-react with rabbit anti-PHA serum were often present. The biochemical characterization of this bean revealed that it consists of two similar glycoproteins, PHA-L and PHA-E, with molecular weights of 30 and 31 kDa, respectively. 18
Despite the fact that relevant research information is available on PHAs, the physiological functions of these glycoproteins remain unclear. Therefore, the current investigation was undertaken to clarify the mode of action of raw Beldia bean ingestion with respect to their lectins on the morphology and function of the small intestine.
Materials and Methods
Animal, housing, and diet
All experiments and treatments were conducted in compliance with the European Union regulations concerning the protection of experimental animals. A Local Ethics Committee approved the study protocol.
Twenty young adult male Wistar rats were purchased from the animal house of the Pasteur Institute, Tunis, Tunisia. Each rat weighted ∼60–80 g at the start of the study. The rats were individually housed in standard polypropylene cages with wood chip bedding. A single handler attended to their care and food hygiene daily. The animal facility was maintained under an inverted 12:12 h light-dark cycle, a constant temperature of 25°C ±1.4°C, and relative humidity of ∼50% ±5%.
After 1 week of conditioning the rats to the environment and feed, they were randomly distributed into two groups of 10 rats each: Group 1 (control) was provided with rodent pellet flour suspension (RPFS; lectin-free) from the Cereal Office of Tunis, and Group 2 (experimental) was provided with Beldia bean flour suspension (BBFS) authenticated by the National Agricultural Research Institute of Tunisia (INRAT), Ariana, Tunis. The same individual administered the feed each day via orogastric intubation using a stainless steel ball-tipped gavage needle attached to an appropriate syringe; the maximum dose was 300 mg of flour suspended in 3 mL of distilled water per animal. 19,20 Both groups were provided with water and consumed basic feed ad libitum throughout the 10-day experiment. Food intake, body-weight, behavior, and stool characteristics were recorded daily.
On the morning of the last (tenth) day of the experiment, one and half hours after their last 300 mg gavage of their corresponding diets, the rats were anesthetized with ethyl ether. Their body temperatures were maintained at 37°C with an overhead heat lamp connected to a temperature regulator. The Food Technology Service (STA) at the National Institute of Nutrition and Food Technology (INNTA), Tunis, Tunisia, analytically determined the protein, fat, sugar, and energy contents of their diets. Briefly, protein was determined via Dumas oxidation using a Carlo Erba nitrogen analyzer. Fat was measured via gravimetric determination following extraction with a chloroform/methanol mixture. 21 Sugars were measured via HPLC following extraction in 80% ethanol. The nutritional compositions of the control and experimental diets are shown in Table 1.
Data are shown as percentages of 100 g of dried product from the mean of two duplicate determinations.
BBFS, Beldia bean flour suspension; RPFS, rodent pellet flour suspension.
Histology of the small intestine
Tissue sampling and processing
For the histological analyses, five intestinal portions (1–2 cm) were removed from the following locations: proximal jejunum, mid-jejunum, jejuno-ileum, mid-ileum, and distal ileum. All tissue samples were longitudinally opened atop a piece of paper, and then rapidly and gently washed with water and immediately fixed overnight in Bouin's solution. Once fixed, the samples were dehydrated in ascending grades of ethanol as follows: one 70° ethanol bath for 50 min, two 90° ethanol baths for 60 min, one 90° ethanol bath for 75 min, one absolute ethanol bath for 70 min, and one absolute ethanol bath for 90 min. After dehydration, the samples were cleared in two toluene baths for 60 and 75 min. Afterward, the samples were embedded in two paraffin wax baths: the first one for 60 min and the second one for an entire night at 60°C under vacuum conditions.
On the following day, the tissues were submerged in paraffin wax and left to dry at room temperature. Following this protocol, the specimens were cut using a rotary microtome (Leitz, Wetzlar, Germany) into 4 to 6 μm sections and mounted onto glass slides. The sections were allowed to dry overnight in an oven. The tissue sections were deparaffinized three times in 95% ethanol and once in 75% ethanol for 4 min. The tissues were rehydrated in running tap water for 10 min and incubated in Ehrlich's hematoxylin for 10 min. The tissue sections were thoroughly washed in running tap water and differentiated in HCl-acidified 95% ethanol. The tissues were thoroughly washed in running tap water until they turned blue in lithium carbonate solution. After the desired blue staining of cellular infiltrates had been obtained, the tissue sections were dehydrated in 95% ethanol for ∼4 min and then stained with eosin Y (1% eosin Y in 95% ethanol) for ∼3 min. After staining with eosin Y, the tissue sections were dehydrated four times in absolute ethanol for 4 min and cleared three times in xylene for 3 min. The tissues sections were mounted in Canada Balsam (C-1795; Sigma-Aldrich Corporation, Missouri, USA) and dried overnight in an oven.
Histological morphometry
Five readings each of villus height (μm) and crypt depth (μm) were taken per specimen. These measurements were performed with a light microscope (Reichert Polyvar Met Optical Microscope) at 100× magnification using an eyepiece that contained a millimeter scale. Villus height was measured from the apical to the basal region, which corresponded to the superior portion of the crypts. The crypts were measured from the base to the region of transition between the crypt and the villus. The mitotic index (i.e., the crypt/villus ratio) was calculated by dividing crypt depth by villus height for each intestinal segment.
Transmission electron microcopy
For transmission electron microcopy, the intestinal tissues were treated as follows: Different fragments of proximal jejunum and mid-jejunum (0.5 cm) were immediately fixed in 4% glutaraldehyde in 0.2 M sodium cacodylate buffer, pH 7.2 (Agar Scientific Ltd., Stansted, United Kingdom) for 1 h at room temperature. The fixed samples were then washed three times with 0.2 M Na-Cacodylate buffer, pH 7.4, in 15-min intervals and postfixed in 2% osmium tetroxide OsO4 (Bio-Rad, Marnes-la-Coquette, France) for 2 h. Thereafter, the specimens were washed three more times with the same buffer. Dehydration was conducted with ascending concentrations of ethyl alcohols from 70° to 100°. When dehydration was complete, the jejunum tissues were infiltrated through two changes of 100% propylene oxide for 30 min followed by a 50:50 mixture of propylene oxide and embedding Epon 812 resin for 12 h. Finally, after incubation in a 50:100 mixture of propylene and embedding Epon 812 resin for 18 h, the tissues were transferred to fresh 100% embedding media for 12 h. Following 24–48 h of polymerization in an oven at 60°C, the blocks were then ready to section. The Epon blocks were sectioned with a Diatome diamond knife using an ultramicrotome Ultra-Cut Reichert. Vertical sections (taken perpendicular to the long axis of villi) were mounted on butvar-coated nickel grids and stained with 4% uranyl acetate for 20 min and 0.2% lead citrate for 10 min for contrast enhancement. All specimens were observed using a JEOL 1010 transmission electron microscope (JEOL Ltd., Tokyo, Japan).
In vivo intestinal perfusion
Male Wistar rats were maintained ad libitum on a diet of commercial rodent pellets. Approximately 15 h before experimentation, food was withdrawn, but the animals were allowed free access to water. On the morning of the study, one and a half hours after gavage (i.e., the time required for PHAs to reach and exert their effects in the jejunal lumen), 22 the rats were anesthetized under ether inhalation. The rats were then placed under a heat lamp to keep their core temperature at 37°C ±0.5°C. Breathing was maintained spontaneously, and the normal color of the ears and feet indicated acceptable oxygenation throughout the experiment. The abdominal wall was opened through a midline incision, and a ligature was placed around the jejunum 5 cm immediately below the pylorus.
The jejunum was then gently flushed with warm Ringer's solution (140 mM NaCl, pH 7.4, 37°C; containing Na+, 140; K+, 5.2; Cl−, 120; HCO3 −, 25; HPO4 −, 2.4; H2PO4 −, 0.4; Ca2+, 1.2; and Mg2+, 1.2, mEq/L) introduced through a hypodermic needle inserted immediately below the ligature. During this washing-out process, the jejunum was repeatedly milked downward to force out the solution. After washing, a 10–15 cm loop of the emptied jejunum was isolated in situ between ligatures, thereby preserving the nerves and vascular pedicle. Tow cannulas were then inserted into both distal and proximal extremities of the jejunal loop, one for perfusion and the other for fluid drainage. The abdominal wall was then closed to prevent tissue dehydration. Both cannulas were exteriorized through the extremities of the abdominal suture.
Ringer's mannitol solution (30 mM mannitol, pH 7.4, 37°C) was maintained in a bottle connected to the catheter infusion pump, which was kept at 37°C in a water bath. The Ringer's mannitol solution (buffered by HCO3 −), was perfused at a rate of 0.5 mL/min for 15 min to obtain basal absorption/secretion in the jejunal lumen. After rinsing the isolated segment of the jejunum with Ringer's solution (140 mM NaCl, pH 7.4, 37°C) for 10 min, the animals received Ringer's glucose solution (30 mM glucose, pH 7.4, 37°C) under the same conditions described above. The effluents were collected separately in test tubes and maintained on ice, then stored in a freezer at −20°C for subsequent biochemical analysis. To determine the jejunal surface area (A), at the end of the experiment, the length and the width of the perfused intestinal segment were measured without stretching.
Biochemical determinations
This study was conducted at the Clinical Biochemistry Laboratory, Institute of Carcinology Salah Azaiez, Tunis, Tunisia. Briefly, the glucose concentration (μmol/μL) of the effluent was determined using an enzymatic method based on glucose oxidase that was estimated from a standard glucose concentration curve. Effluent potassium and sodium ion concentrations were measured using flame photometry. The results are expressed as the difference between influx and efflux.
Data calculation
The fluxes of glucose and electrolytes (Jg
and Je
, respectively, in μmol/min/cm2 and μEq/min/cm2) and the flux of water (Jw
in μL/min/cm2) were calculated using the following formulas:
where C(g;e)in represents either the glucose or electrolyte concentrations of the infusate (μmol/μL of glucose and μEq/μL of electrolyte); C(g;e)out represents either the glucose or electrolyte concentrations in the outflowing perfusate; Vin and Vout are the respective volumes of inflowing and outflowing perfusates (μL) for Δt intervals (min); A is the jejunal surface area (cm2); and e is the electrolyte (Na+ and K+).
Preliminary experiments indicated that when 140 mM NaCl and 30 mM mannitol solution was injected; neither electrolytes nor water moved transmurally into the jejunum. These concentrations were then incorporated as the basis of a solution containing Ringer's mannitol solution. [ 3 H]-PEG (polyethylene glycol 4000) and [ 3 H]-D-glucose were included as markers for water flux and active transport, respectively. [ 14 C]-Mannitol was used as an extracellular space marker. Osmolality was ∼300 mOsm/kg. Comparisons between the control and experimental groups were based on the means derived from 10 loops.
Statistical analyses
All values were expressed as the mean ± standard error of the mean. Tests of significance were performed using Student's t-tests (paired and unpaired); the level of significance was set at 5% (P < .05).
Results
Animal status
No mortalities or clinical signs were observed based on the daily ingestion of raw BBFS at a dose of 300 mg for 10 days.
Body weight
The effect of raw BBFS on body weight is presented in Figure 1. This figure shows the growth curves of the control and experimental groups during the 10-day experiment. The oral administration of BBFS at a dose 300 mg did not significantly affect the weights of rats; compared with the control group receiving RPFS; P > .05.

Growth curves of rats gavaged daily with 300 mg RPFS (control group) or 300 mg BBFS (experimental group) for 10 days (mean ± standard error of the mean of 10 rats). BBFS, Beldia bean flour suspension; RPFS, rodent pellet flour suspension.
Small intestine morphology
Light microscopic findings
The light microscopic examination of the proximal jejunum and the mid-jejunum of the experimental group at 10 days, after BBFS gavage, revealed conspicuous villus elongation (Fig. 2B, D). The goblet cell number increased in the villous and crypt epithelium. However, the appearance of the epithelial enterocytes was comparable with the group treated with RPFS (data not shown).
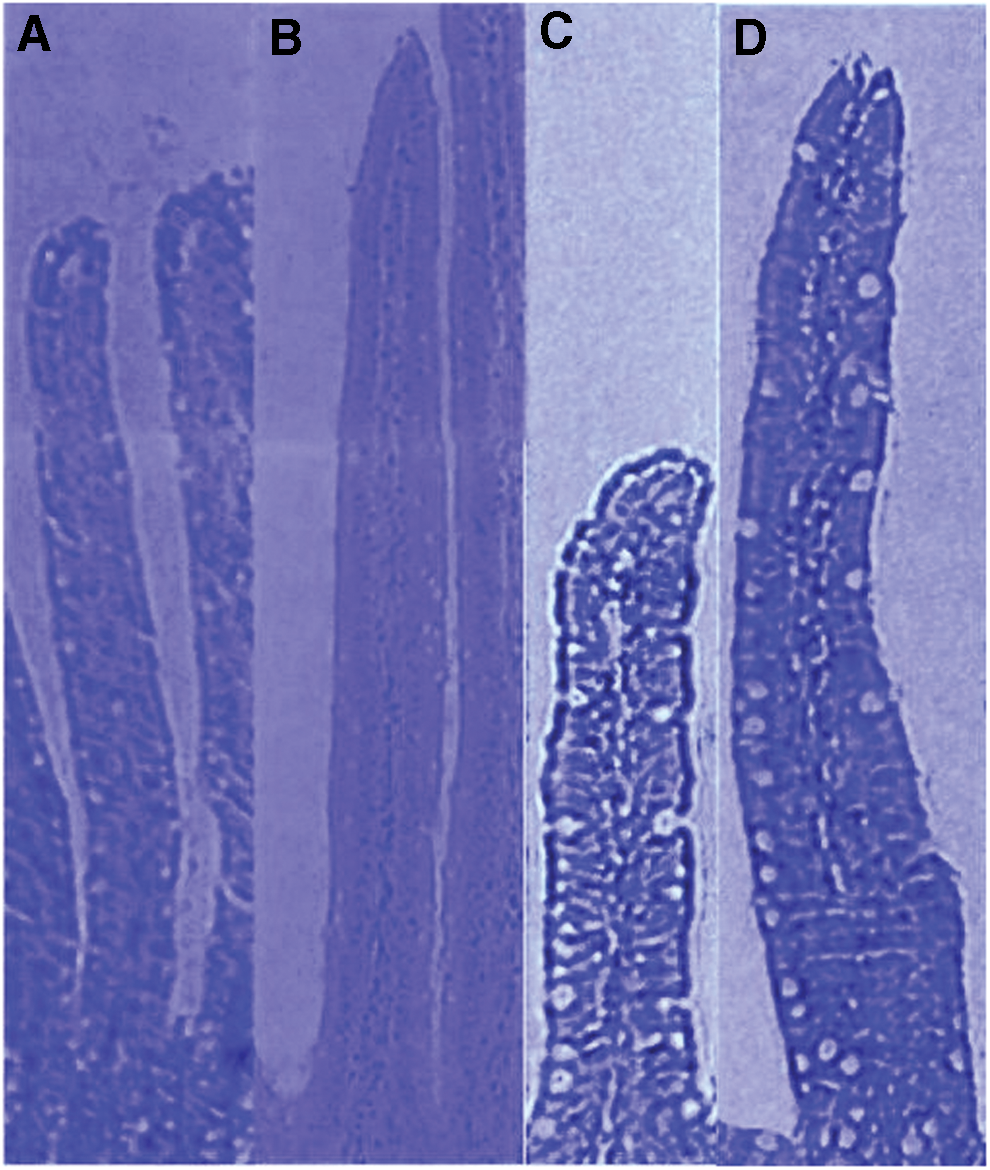
Both villous height and crypt depth are important indicators of rat digestive health and directly related to the absorptive capacity of the mucous membranes. Theoretically, villous height reflects the balance between the mitotic activity of the crypt enteric cells 23 and the desquamation produced principally by external aggressors. 24 The morphology of the small intestine at day 10 is highlighted in Table 2. The histological study showed that the oral administration of BBFS significantly increased the villus height, crypt depth, and crypt depth/villus height ratio in rats' jejunums and ileums compared with control values (P < .05). The changes were of greater magnitude in the jejunum than the ileum.
Mean value ± SEM.
Significantly different from corresponding control value, P < .05.
Significantly different from corresponding control value, P < .02.
Significantly different from corresponding control value, P < .01.
Significantly different from corresponding control value, P < .001.
SEM, standard error of the mean.
Electron microscopy findings
The morphology picture of epithelial mucosa varied among the treatment groups, from normal in the animals gavaged with the RPFS to abnormal in the animals orogastrically fed BBFS.
Determining whether the BBFS caused a structural change in the microvilli is difficult based solely on the longitudinally sectioned images (Fig. 3A, B) of jejunal enterocytes. The electron scanning micrographs (Fig. 3D, E) clearly show that the microvilli of the rats receiving BBFS were altered.

The electron micrographs of the jejunum of the control animals exhibited well-arranged microvilli that were uniform in size, shape, and density. No electron microscopy evidence suggested cellular degeneration or necrosis. The epithelial lining of the mucosa was intact, and no vesicles were observed (Fig. 3C). The cellular organelles; such as the endoplasmic reticulum, Golgi bodies, and mitochondria appeared normal, and no obvious signs of damage were detected. However, the jejunums of the animals gavaged with BBFS showed evidence of marked ultrastructural changes in the enterocyte microvilli. Clusters of small vesicles, tied off from the microvilli, were found in higher number when compared with control jejunums. The terminal web was slightly disorganized just beneath the brush border. The degree of abnormality of the microvilli varied from short and dense (Fig. 3E) to long and slender (Fig. 3D).
Absorption activity
Table 3 displays the effects of the daily gavage of 300 mg of raw BBFS over 10 days, on the jejunal water, electrolytes, and glucose movements. Under basal conditions (i.e., perfusion with Ringer's mannitol solution), the water, sodium, and potassium uptakes were significantly decreased in rats challenged with BBFS compared with the control group (P < .02, P < .05, and P < .05, respectively). The glucose absorption throughout the isolated loop was not significantly reduced in the group treated with BBFS (0.30 ± 0.16 μmol/min/cm2) compared with the controls (0.44 ± 0.28 μmol/min/cm2).
Mean value ± SEM of 10 loops.
Significantly different from corresponding control value, P < .02.
Significantly different from corresponding control value, P < .05.
Not significantly different from corresponding control value, P > .05.
Table 4 summarizes the jejunum response to glucose in the experimental and control groups (i.e., perfusion with Ringer's glucose solution). Interestingly, the absorption of water and ions was favorably enhanced in the presence of glucose. However, no significant differences were recorded with regard to the uptake of these molecules between the two groups of animals (Table 5; P > .05).
Mean value ± SEM of 10 loops.
Not significantly different from corresponding control value, P > .05
Mean value ± SEM of 10 loops.
Not significantly different from corresponding control value, P > .05.
ΔJ, Difference between the flux in the presence of glucose and the basal flux.
Discussion
The most striking finding of the present investigation is that a daily gavage of 300 mg raw BBFS for 10 consecutive days provoked profound morphologic and functional effects on the small intestines of rats. With regard to previous nutritional studies, the most promising strength of the experimental protocol is that the morphometric and functional measurements were taken together from the same subjects. However, several reports in the literature have separately examined the effects of kidney beans (P. vulgaris L.) or their PHAs on the structure of the small intestine mucosa and their function.
In addition, this study was much more concerned with the chronic effects of white kidney beans rather than the acute effects. This difference might explain the divergence that was found between the current data and those reported in the literature. Changes in permeability that obviously occurred under basal conditions, in water and electrolyte movement, cannot be ascribed neither to the age nor to the nutritional status of animal. In fact, clinical and experimental malnutrition are well known to modify the mechanisms of intestinal absorption and secretion. These effects are chiefly associated with profound histological and functional alterations, and are more pronounced in developing animals than adults, accounting the importance of energy and nutritional needs for organism development. 25
The feeding experiments clearly showed that the changes in body-weight and daily weight gain among the BBFS-treated rats were approximately similar to those of RPFS-treated rats. The former animals did not reduce their food intake (data not shown) and no symptoms of gastroenteritis (e.g., anorexia, vomiting, or diarrhea) were recorded.
The inclusion of raw kidney beans (P. vulgaris L.) in diets can reduce feed consumption, induce failure to thrive, and deteriorate the nutritional status of animals, thereby leading to its eventual death. 26 However, the findings of this study disagree with several earlier lines of experimental evidence demonstrating the capacity of P. vulgaris L. to effectively reduce the weight of animals.
For example, Honavar et al. 27 demonstrated that diets containing raw beans or orally administered bean agglutinins caused weight loss and death in rats. Jaffé, 28 reported that rats fed with a diet containing ∼40% ground raw beans (P. vulgaris) rapidly lost weight and died within 10 days. Pusztai et al. 29 showed that the inclusion of different levels of raw kidney beans (P. vulgaris) with high lectin contents (27 g/kg meal) depressed the growth of both obese and lean rats. Carai et al. 30 obtained similar results when they tested P. vulgaris dry extract, showing that it resulted in a decrease of daily food intake and body weight in obese Zucker rats. Bardocz et al. 31 also reported an impairment of growth among animals fed diets containing red kidney beans.
The primary effect of PHA, as far as growth was concerned, appeared to be a suppression of appetite. Liener, 32 also reported that pepsin inactivated the hemagglutinating activity. Since these earlier studies were conducted, several researchers have noted a marked reduction in protein digestibility and a growth inhibitory effect caused by appetite suppression when purified lectins are added to diets. With regard to mode of action, Jaffé, 33 postulated that the toxic effects of lectins, when ingested orally, are due to their ability to bind to specific receptor sites on the surfaces of intestinal epithelial cells, thereby causing nonspecific interference via the absorption of nutrients.
On the other hand, several investigators have suggested that certain bean cultivars are completely devoid of poisonous effects. Pereira et al. 34 stated that white bean flour (P. vulgaris) administered at 1 mg/g body weight over 21 days produced a normal rate of growth in rats. 31 Rigas and Osgood 35 did not observe poisonous properties in a bean agglutinin that they had prepared. Nciri, 36 created a crude preparation from white bean (P. vulgaris var. Twila) that failed to exhibit any deleterious effects on the growth of rats fed at a concentration of 100 mg/mL, at which kidney beans are definitely toxic.
One potential source for these discrepancies in the literature is that considerable variation exists in lectin concentration, nature, and type in P. vulgaris bean cultivars. 37 –39
High-PHA bean varieties are usually responsible for stunted growth in animals, whereas low-PHA bean varieties have no appreciable effect on body-weight gain and are nontoxic in rats. In an experimental study with rats, cultivars of kidney bean (P. vulgaris) provided half of the protein (10% level) of the diet, with the remainder being in the form of casein. Protein use was depressed in relation to the amounts of lectin present in each cultivar; the cultivar “Pinto III”, which was almost free of lectin, had no appreciable nutritional toxicity; however, the high-lectin cultivar “Processor” was much more deleterious. When fed to rats as the sole source of protein, diets containing the latter cultivar produced greater weight loss than that recorded for animals receiving protein-free diets. 22
Honavar et al. 27 fed lectins from kidney beans and black beans to rats and found that the former was more toxic than the latter. Jaffé and his research group observed that when the extracts of different bean types were tested by injection in mice 40 or by feeding ground seeds to rats, 41 only the saxa and porillo bean varieties were toxic, whereas the cubagua and mountaineer half runner bean varieties were of low toxicity. Pusztai et al. 42 found that 11 of 13 cultivars of kidney beans were highly toxic to rats in their raw state. On the other hand, two low-lectin bean varieties, Pinto III and great northern, were essentially nontoxic to rats.
Cultivars of P. vulgaris display marked differences in toxicity, and, for reasons that are not fully understood, only those that have lectins that specifically agglutinate trypsinized cow cells are toxic. 40 –44 The nontoxic cultivar “Pinto III” was found to possess a lectin that had two subunits instead of the four subunits that are characteristic of the lectins found in most of the toxic cultivars. 45 This lectin did not inflict the usual damage to the intestinal mucosa that is associated with the ingestion of toxic beans. 22 The lack of toxicity in “Pinto III” beans is likely a consequence of their lower valency (i.e., two subunits instead of four), which might weaken their affinity for cell-surface receptors.
Evidently, differences in formulation, animal species, strain, duration and frequency of treatment, dose levels and ranges, nutritional conditions, and individual susceptibility might also be responsible for different toxic effects.
For instance, Honavar et al. 27 observed that kidney bean agglutinin markedly inhibited growth and caused death within 2 weeks in rats fed at a 0.5% level in the diet. In a similar experiment with chicks, Wagh et al. 46 noted that this growth decrease was much less than that in rats, and no lethal action was detected. The susceptibility of mice of different strains to injected black bean agglutinin varies. 47 However, Grant et al. 48 concluded that the toxicity of the P. vulgaris lectins is independent of the age or maturity of experimental animals after pairing feeding rats aged between 30 and 123 days. No significant age group differences were detected with regard to protein use, food intake, disruption of intestinal lining, or circulating anti-lectin antibodies. These results definitely confirm the observations of Pusztai et al. 49 who showed that the inclusion of raw kidney beans in the diets of young and mature rats resulted in rapid weight loss and eventual death.
In a preliminary attempt, this study fed rats a diet containing fractions of raw ground beans var. Beldia. Accordingly, these animals refused to eat and started to lose weight compared with controls. In light of this behavior, the administration of BBFS via a stomach tube was chosen, leaving the animals to eat conventional commercial feed. The nutritional status of the rats then remained satisfactory throughout the experimental period. Thus, a malnutrition factor did not bias the observations and measurements taken from the small intestines.
Histomorphometric analyses indicated changes in mucosal architecture in terms of increased villus height, crypt depth, and the mitotic index of jejunum and ileum in raw BBFS-fed animals, compared with the corresponding controls.
The results from this study match those obtained by Hawn Roe et al. 50 who emphasized the marked increase in villous height and crypt depth in a crude red kidney bean-fed group compared with a control group. These trends were observed in the proximal, middle, and distal segments of rat small intestines. Zucoloto et al. 51 demonstrated similar effects in rats. They indicated that the length of the ileal villi in rats fed a diet containing 4% active PHA was significantly higher than that in a group supplemented with a diet composed of inactive PHA. Despite its general toxicity, PHA is clearly a potent growth factor for the gut. Although this lectin binds to the upper half of absorptive villi, its most dramatic effects occur in the proliferative compartment. Thus, the villi and the crypts of Lieberkuhn in the jejunum and the ileum increased in length after 10 days of BBFS treatment, indicating initially high metabolic and mitotic activities.
Scanning electron microscope studies have revealed that the jejunum preserved its normal structure after placebo (i.e., RPFS) administration for 10 days. Under normal circumstances, intestinal epithelial tissues maintain structural integrity via continuous cell proliferation; this action is associated with the normal development of the endoplasmic reticulum, golgi bodies, and mitochondria. However, after the exposure of the rat intestinal epithelium to BBFS, the jejunum demonstrated anomalous effects; such as the disruption of brush borders, abnormally short and thick or long and thin microvilli, and the presence of scattered vesicles near the brush border membrane. The presence of these vesicles might reflect an increased turnover of microvillus membrane.
Several investigators have reported that dietary kidney bean lectins are usually associated with abnormal intestinal histology and serious alterations in the microvillus border.
For instance, Rossi et al. 52 offered a similar interpretation and postulated that when rats were maintained for 5 days on a diet containing purified lectins extracted from a Brazilian variety of white kidney beans (i.e., P. vulgaris, var. “Jalo”), they developed marked ultrastructural changes in the epithelium of the proximal jejunum, whereas both pair-fed and ad libitum-fed controls did not.
In an article entitled “The nutritional toxicity of P. vulgaris lectins”, Pusztai et al. 53 used electron microscopy to observe irregularly arranged, stunted, elongated, fragmented, or vesiculated microvilli were presented on the apical surfaces of the duodenal and jejunal enterocytes in rats fed diets containing high-lectin “Processor” beans or purified bean lectins. However, they also noticed that the brush borders of rats fed diets containing low-lectin “Pinto III” beans or control diets were not damaged.
Three years later, the same group of researchers 54 demonstrated that, when rats were fed for 10 days with a diet containing 10% cooked “Processor” bean protein, the appearance of jejunal microvilli was normal; however, when they were fed a diet containing 10% raw “Processor” bean protein for only 3 days, extensive disruptions in the jejunal microvilli were recorded. Many of these microvilli were longer and thinner than those of the control group. Likewise, vesiculated membranous debris and flocculent and filamentous electron-dense material were often associated with the microvilli. Zucoloto et al. 51 obtained similar findings as those reported in this study. Rats that ingested active PHA showed marked ultrastructural changes in their villus enterocytes, consisting mainly of abnormalities in the microvilli, which were shorter, slightly thicker, irregular, and more sparse.
Using transmission electron microscopy, Kik et al. 55 demonstrated that the microvilli were shorter and irregularly positioned when pig jejunal mucosa was cultured for 5 h with P. vulgaris isolectins PHA-E4 and PHA-L4. The high phytolectin content of Beldia seeds (∼9.20 g PHA/kg bean flour) 18 was most likely responsible for the abnormal damage that occurred in rat small intestines.
Light and electron microscope analyses demonstrated an increase in goblet cell numbers and a substantial secretion of mucins in the small intestine epithelium of BBFS-treated rats. These observations, coupled with the stimulation of secretion, might translate into a nonspecific defense response of the intestine toward aggravations of the intraluminal origin. This effect can be observed in the presence of bacterial toxins; such as cholera toxin and other invasive, and noninvasive species. 25,56
In general, goblet cells contain mucins, trefoil proteins, and other factors that help protect intestinal mucosa from injury and facilitate tissue repair. 57 This mucus gel layer is an integral structural component of the intestine and acts as a medium for protection, lubrication, and transport between luminal contents and epithelial cell lining. 58 This reaction presumably occurred after the prolonged exposure of the epithelium to irritating agents derived from Beldia bean flour, such as PHAs and other bioactive constituents.
Studies addressing the effect of kidney beans (P. vulgaris L.) and their lectins on intestinal absorption are sparse. Thus, an attempt was made to characterize the effect of raw Beldia beans on the water, electrolytes, and carbohydrate absorption in the small intestines of rats. In vivo intestinal loop experiments have revealed that the basal fluxes of water and electrolytes were significantly reduced in BBFS-treated rats compared with controls.
The sodium ion results match the data published by Dobbins et al. 59 who investigated the effect of PHA on the ion transport in the rabbit ileum in vitro. These authors noted that, compared with control tissues, the net Na+ absorption decreased when PHA was added to a mucosal solution. Kunzelmann et al. 60 also obtained similar results when they investigated the effect of dietary lectins on the ion transport in the mouse epithelia. They observed that erythroagglutinin (PHA-E), leuagglutinin (PHA-L), and concanavalin A (Con-A) but not wheat germ agglutinin and soy bean agglutinin, inhibited electrogenic Na+ absorption in a dose-dependent manner in both the colon and the trachea.
These researchers believe that the inhibition of Na+ absorption essentially occurs because the lectins fix to the proteoglycan receptors in the epithelium. They added that the inhibition of electrogenic Na+ absorption by Con-A or P. vulgaris lectins will shift the net transport of colonic epithelium from NaCl absorption to secretion. This sequence is likely associated with the diarrhea-inducing effects of these lectins.
In the current physiological research, measurements were taken in vivo and the obtained fluxes represent net fluxes that refer to the differences between influxes (i.e., the movement of particles from the intestinal lumen into the bloodstream) and effluxes (i.e., the opposite direction of an influx). In general, the reduction of absorption could be attributed either to a decrease in influx or to an increase in secretion. 61 Therefore, it is logical to assume that the decrease in water and ion uptake in the animals treated with BBFS might have arisen from stimulated secretion. The morphometric studies that have revealed the considerable elongation of the crypt of Lieberkuhn depth in BBFS-gavaged rats compared with controls strengthens this hypothesis.
Deeper crypts indicate faster tissue turnover, which permits the renewal of the villus as needed. 62 Crypt cells are well known as highly specialized in the process of intestinal secretion, 56 whereas villus cells are more involved in water, electrolyte, and nutrient absorption. 63 The current results somewhat agree with earlier reports; 2,22,53,64 describing many cases of experimental and clinical diarrhea in animals or humans following the ingestion of either raw or improperly prepared kidney beans.
The literature shows that the oral intake of PHA causes intestinal maldigestion and malabsorption. 65,66 This malabsorption might be caused by damage to the enterocytes and villus atrophy. 67 –69 PHA might alter the absorption of nutrients via its effects on transport across the intestinal membrane. 70 Therefore, it is reasonable to conclude that the secretory effect of BBFS is functional and not related to mucosal alterations because no apparent organic alterations were recorded in the intestinal mucosa of this study.
PHAs are glycoproteins of nonimmunological origin that specifically recognize sugar molecules; therefore, they are capable of binding to glycosylated membrane components. This interaction might not only alter the absorption process, but also the cellular internalization along different pathways (lysosomal and nonlysosomal), depending on the biological signal transmitted to the epithelial cells upon the binding of specific bioadhesive ligands. 53
No significant difference was observed with regard to the rate of glucose uptake between the control and experimental groups. This finding considerably differs from several published physiological studies. Donatucci et al. 71 examined the effect of the binding of navy bean (P. vulgaris, “Sanilac” variety) lectins to the intestinal cells of rats regarding the absorption of glucose. These authors found that the rate of absorption of [ 14 C] glucose in the intestines of rats fed raw navy bean flour or casein plus lectin was approximately one-half of that of their counterparts fed autoclaved flour or casein alone. Gislason, 72 noted that the absorption of glucose from ligated intestinal loops in anesthetized rats, previously fed a bean diet or administered black bean agglutinin via a stomach tube, greatly decreased. Freed 73 stated that lectins reduce intestinal glucose uptake by 50%.
In fact, glucose absorption and its stimulating action on the transport of water and electrolytes, in the jejunal loops of rats administered BBFS orogastrically, were comparable with controls. This finding conflicts with the morphometric data, showing an increase in villus height, highlighting the greater transport potential of BBFS-fed rats. This discrepancy can be easily explained by analyzing the crypt depth/villus height ratio, which was significantly greater in rats administered BBFS via orogastric intubation than in the control group, providing evidence of an efflux (secretion) of water and electrolytes that reduced the net absorptive fluxes of H2O, Na+, and K+. However, it cannot be explained for glucose.
Normally, glucose flux should drastically increase in animals orally administered BBFS, whose villous height was significantly higher than controls. The following hypothesis might clarify this situation: The stimulation of cell division in the crypts of Lieberkuhn, as substantiated by the prominent increase in the mitotic index of BBFS-fed rats, leads to a rapid increase in the proportion of immature cells in the jejunal villi. This result would influence the physiological roles of enterocytes and alter the crypt/villus ratios. Enterocytes acquire their abilities of transport (transporters), digestion (disaccharidases, peptidases), and their cytoskeletons during the migration of crypts to the tip of villi. This change would result in an excess of secretory cells and glucose imbalance. Additional autoradiographic analyses are required to confirm this hypothesis, which is based on functional data only.
Under normal circumstances, glucose enhances the mucosal-to-serosal transport of sodium and water in the small intestines of humans 74 and animals. 75 This enhancement remains intact in animals exposed to choleragen 76 or E. coli enterotoxins. 77 –79 Similar to this study, the intraluminal glucose-enhancing effect was comparable between BBFS-treated and control animals. Because actively transported organic metabolites usually stimulate fluid and electrolyte absorption, this effect might indicate that the proximal jejunums in both of the groups of growing rats possessed relatively similar capacities in their transport systems for glucose. This glucose-mediated effect might be effectively used in oral PHA oral therapy among humans by employing oral electrolyte solutions with glucose.
According to Banwell et al. 80 bacteria also play an important role in the pathogenesis of kidney bean lectins in rats. They demonstrated that exposure to purified PHA in the diet changed the microbial ecology of the small intestine. These changes in the microbial flora contributed to the malabsorption of nutrients in the small intestines of PHA-fed animals, a finding that requires additional investigation. Importantly, no correlation was found between morphometry and function variables in rat jejunum.
At present, the most critical issue consists of determining the major agent (s) in raw Beldia beans, responsible for all of the aforementioned effects. Beans are a good source of protein, complex carbohydrates, dietary fiber, several vitamins, and minerals. In addition to these nutritional components, common beans are rich in a variety of several phytochemicals with potential health-harmful compounds; such as polyphenolic compounds, lectins, trypsin inhibitors, and phytates, among others. 81 Some of these bioactive components impair the absorption of nutrients and minerals both in living animals and semi-isolated intestinal preparations. 8,82,83
In this study, it is most likely that Beldia bean lectins bound to specific monosaccharides on brush border cells and severely disrupt the normal morphology of the gut epithelium, leading to absorption modifications without impairing animal growth or feed consumption. Muzquiz et al. 84 conducted chemical studies and noted that both environmental and genetic factors clearly influenced the content of PHAs in certain dried beans. The direct effects of PHA are highly comparable with those generated by the common bean P. vulgaris. 52 However, PHA concentrations appear to be decisive in the development of bean toxicity because it has a dose–response relationship. 19,85
In a recent investigation, native PHA was characterized in a Beldia bean flour, administered to rats via gavage, using different immunobiochemical techniques, such as hemagglutination tests, competitive ELISA, SDS-PAGE, and Ouchterlony double immunodiffusion. The combination of these techniques convincingly showed that Beldia variety seeds effectively contain a considerable amount of PHAs (∼9.20 g/kg beans flour) in biologically active form. 18 Moreover, other in vivo serological tests conducted at the Intestinal Immunophysiology-Research Unit at the Faculty of Medicine of Tunis have enabled the detection of immunologically intact PHAs in the rat jejunal lumen, as early as 3 h and 30 min after a final challenge with 300 mg of BBFS, revealing their higher resistance to gut proteolytic enzymes. 86
This finding might account for their oral toxicity. Simultaneously, Hagen et al. 20 showed that PHA bound to the upper third of the absorptive villi, 2 h after the oral administration of a 300 mg suspension of pulverized raw kidney beans (Montcalm variety) to rats.
The possibility of the transfer of PHAs, in their native forms from the intestinal lumen into systemic circulation and all of their functional consequences has not been excluded. Numerous studies have postulated that dietary proteins of animal and vegetal origins (β-lactoglobulin, ovalbumin, lactoferrin, gliadine, and so on.) remain intact when crossing the intestinal epithelium. 87,88 The amount of protein absorbed did not play a nutritional role in an energetic and plastic sense, rather, it had a functional regulatory role driven by a complex system, coordinated by the different cells present in the chorion (immunocompetent cells, nerve cells, and endocrine-paracrine [APUD] cells). 89 –91
In fact, this transfer depends broadly on the concentration of protein that comes into contact with the apical membranes of epithelial cells. In turn, this concentration is intimately linked to gastric (i.e., the secretion of HCl and pepsin) and pancreatic (i.e., the secretion of pancreatic proteases) activities, which are highly dependent on the nutritional status of the subject in question. Usually, malnutrition cases are accompanied by gastric and pancreatic insufficiencies. The specific inhibiting action of PHA on gastric secretion should be added to these effects. 92,93 Under these conditions, the intraluminal native PHA content might readily increase, thereby facilitating its access to intestinal cells.
In conclusion, the results of the present investigation are of the upmost nutritional, immunological, and physiological importance.
From a nutritional standpoint, the excessive consumption of raw (e.g., soaked or sprouted in salads) or insufficiently cooked (e.g., prepared in slow cookers) common beans might induce organic and functional implications, following the interaction of P. vulgaris L. lectins with the intestinal mucosa. Furthermore, this interaction might elicit local (gut-associated lymphoid tissue) and systemic (mucosa-associated lymphoid tissue) immune responses. The information gained from the study of these phenomena might lead to a better understanding of the mechanisms of tolerance and the disorders associated with dietary proteins.
Physiologically, studying the interaction between the intraluminal content (which is rich in proteins) and epithelium and chorion cells enables a more rational approach toward identifying the physiopathological mechanisms of certain intestinal pathologies; such as celiac disease, chronic inflammatory bowel disease, and food allergies. In addition to the lectins, the effect of other protein components of the seeds (e.g., phaseolin [Glycoprotein II]) and their breakdown products (i.e., peptides and oligopeptides) on absorption and digestion should be reconsidered. These mechanisms should be investigated by placing greater emphasis on the mediatory, messenger, and regulatory roles played by these macromolecules at the mucosa level.
Footnotes
Acknowledgments
A grant from the Intestinal Immunophysiology-Research Unit (02/RU/09-02), Faculty of Medicine of Tunis, University of Tunis El Manar, Tunis, Tunisia, supported this investigation. We are greatly indebted to Prof. Mohamed Habib Jaafoura from the Laboratory of Histology-Embryology, Faculty of Medicine of Tunis for his excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.