
Abstract
Although kidney bean (Phaseolus vulgaris L.) lectin toxicity is widely known, its effects in the gastrointestinal tract require further study. This investigation aimed to identify and characterize phytohemagglutinins (PHAs) in the small intestine and sera of rats following oral challenge with ground white beans. Twenty young, adult male rats were divided randomly into two groups of 10 animals each. The control group underwent gavage with a suspension of 300 mg of rodent pellet flour. The experimental group was administered a 300 mg Beldia bean flour suspension (BBFS). After 10 days of daily treatment, jejunal rinse liquid (JRL) and ileum rinse liquid and secretions, as well as sera, were collected. All biological fluids were screened for lectin reactivity using competitive inhibition ELISA, Ouchterlony double immunodiffusion, and immunoelectrophoresis techniques. The results revealed the presence of immunogenic intraluminal PHAs 3–4 h after the oral intake of the BBFS in the JRLs as well as in the jejunal and ileal secretions; however, no PHA was detectable in the rat sera. Ingestion of raw Beldia beans may lead to interaction between PHAs and the mucosa of the small intestine, potentially resulting in an inflammatory response.
Introduction
P
P. vulgaris lectins, also referred to as phytohemagglutinins (PHAs), are carbohydrate-binding glycoproteins found in many species of beans. One of the striking biological features of PHAs is their ability to survive digestion in the alimentary canal of consumers. This feature allows PHAs to bind to the glycosylated membrane of the cells that line the digestive tract. 5 –7 This interaction triggers a cascade of detrimental local and systemic reactions, making this class of molecules antinutritional and/or toxic. Locally, they adversely influence the turnover and loss of gut epithelial cells, damage the epithelial lumen, and impede digestion and absorption processes. 8,9 Systemically, they disturb protein, carbohydrate, and lipid metabolism, 10 –12 promote enlargement and/or atrophy of key internal organs and tissues, and cause altered hormonal and immunological status. 13,14 At high intake amounts, PHAs can gravely threaten the growth and health of the animal or human who consumes them. 2 –4
The fact that PHAs are so widely distributed in food items that are commonly consumed by humans raises the important question of whether they pose any significant risk to human health. Fortunately, most PHAs are easily destroyed by the traditional methods of household cooking. Nevertheless, under special conditions, complete detoxification may not always be achieved, especially if ground seeds are used or industrial processes for quick cooking are applied. Because PHAs are resistant to inactivation by dry heat, 15 practices such as the use of raw bean flours in baked goods should be viewed with caution.
The presence of nutritionally significant amounts of active PHAs in fresh and processed foods and the lack of public knowledge concerning the deleterious effects of dietary lectins on the gut and general health have led to several cases of human intoxication.
For example, an outbreak of what appeared to be food poisoning occurred in England in 1976. A party of schoolboys on holiday ate kidney beans that had been soaked in water, but not cooked. All nine boys who ate the beans became acutely nauseated within 60–90 min after eating and experienced vomiting, which progressed to diarrhea. 16 Eating as few as four to five beans was sufficient to produce these reactions. Two of the boys were admitted to the hospital and required intravenous infusion; however, recovery was rapid in all cases. Noah et al. 2 reported seven incidents involving 43 persons in which the poisonings were attributed to toxins present in uncooked or partially cooked kidney beans (P. vulgaris). In 1981, a BBC television program cited 330 outbreaks of kidney bean food poisoning episodes involving 880 people. 17
In 1988, a hospital launched a healthy eating day in its staff canteen at lunchtime. One dish contained red kidney beans, and 31 portions were served. Between 3 and 7 pm, 11 customers experienced profuse vomiting, and some developed diarrhea. No pathogens were isolated from the food, but the beans contained an abnormally high concentration of PHA. 18 Illness after the consumption of raw beans as part of a salad or in dishes, such as stew, casserole, or chili con carne, cooked in a slow cooker is not uncommon. The times and temperatures involved in the slow cooking of kidney beans under household conditions may not be sufficient to destroy all lectin activity. Warnings often found on labels of dried kidney beans sold in the retail food markets in England and several other European countries recommend that the beans be boiled for at least 10 min before consumption. Recently, the importation of dry red kidney beans (a variety of the species P. vulgaris) for cultivation or consumption in South Africa was prohibited because of their potential toxicity to humans. 19
Bean poisoning is likely primarily initiated by the interaction of these lectins with the carbohydrates in the brush border membranes of the intestinal mucosa, but knowledge about their behavior after ingestion is lacking. It is, for instance, crucially important to test for the presence of lectins in the gastrointestinal tract, especially for beans consumed raw (soaked or sprouted, as for salads) or undercooked (prepared in dehydrators, slow cookers, or crock-pots). 20 –24 To help fill this gap, this study was conducted to identify and characterize the PHAs derived from raw white kidney beans of the Beldia variety in the rat small intestine. The findings emerging from this work have been the object of several previous investigations. 25,26
Materials and Methods
Plant material
White common kidney beans (P. vulgaris L.) of the Beldia variety were purchased from a local market in Ariana City, Tunis, Tunisia. The Beldia beans were authenticated at the Department of Botany, National Agricultural Research Institute of Tunisia (INRAT), Ariana, Tunis, Tunisia. Samples were free of dust, foreign material, and small broken kernels. The seeds were later ground in a mortar and pestle, packed in colored glass containers, and stored at 4°C until further use.
Protein extraction from raw Beldia beans
Protein extraction was conducted under the supervision of Prof. Mohamed Boussaïd at the Department of Chemical and Biological Engineering, National Institute of Applied Science and Technology (INSAT), Tunis, Tunisia. Briefly, the Beldia beans were soaked in distilled water at room temperature overnight to soften the seeds. Then, they were finely ground in a mortar and pestle at 4°C with Tris-NaCl buffer (50 mM Tris-HCl, 50 mM NaCl, pH 8). Subsequently, the mixture was centrifuged at 328 × g for 30 min at 4°C and filtered through Whatman No. 1 filter paper. Finally, the liquid supernatant (total crude extract) was precipitated using ammonium sulfate under different pH conditions (4.2, 5.5). Protein fractions were collected by centrifugation at 328 × g for 30 min, exhaustively dialyzed against Tris-NaCl buffer (pH 8) in 10-kDa molecular weight cutoff dialysis tubing, and subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Samples were stored at −20°C for further use.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
SDS-PAGE was performed under reducing conditions. As standard proteins, α-lactalbumin, trypsin inhibitor, carbonic anhydrase, ovalbumin, albumin, and phosphorylase with molecular weights of 14.00, 20.00, 30.10, 43.00, 67.00, and 94.00 kDa, respectively, were used. The standard proteins, purified PHA and phaseolin (Sigma Chemical Co., St. Louis, MO, USA), as well as total protein extracts of the Beldia beans extracted at pH 4.2 and 5.5, were treated for 5 min at 100°C in 0.0625 mM Tris-HCl buffer (pH 6.8) containing 10% (10 g/100 mL) SDS, 10% glycerol, and 0.1% bromophenol blue. Reduced protein samples were analyzed by electrophoresis in a 12% (12 g/100 mL) acrylamide gel containing water, 30% acrylamide, 1.5 M Tris-HCl (pH 8.8), 10% ammonium persulfate, and TEMED (NNNN-tetramethyl ethylene diamine).
A stacking gel (10 mL) containing 5.3 mL water, 2 mL 30% acrylamide mixture, 2.5 mL 0.5 M Tris-HCl (pH 6.8), 0.1 mL 10% SDS and 0.1 mL 10% ammonium persulfate, and 5 μL of TEMED (NNNN-tetramethyl ethylene diamine) was prepared and used. The gel was mounted in the electrophoresis apparatus (Apelex Laboratory, Lisses, France) in Tris-glycine electrophoresis buffer containing 25 mM Tris and 250 mM glycine (pH 8.3), and 0.1% SDS was added to the top and bottom reservoirs. The electrophoretic run was conducted at constant current of 30 mA for 5 h. The bands were visualized by 0.1% Coomassie brilliant blue (G-250) staining, followed by destaining in 10% acetic acid containing 25% methanol.
Hemagglutination assay
Agglutination of the red blood cells by the crude Beldia bean (P. vulgaris L.) extract was performed as described previously by Confalonieri et al. 27 Fresh human blood (group O+) generously provided by the Blood Banks, Hospital Charles Nicolle, Tunis, Tunisia, was used in this study. A 2% suspension of erythrocytes, obtained by centrifugation of the blood at 241 × g for 10 min, was prepared in 0.9% phosphate-buffered saline (PBS, pH 7). The activity of serial dilutions of the extract (1/1, 1/2, 1/4, 1/8, 1/16, 1/32, 1/64, 1/128, 1/256, 1/512, and 1/1024) was investigated against blood group O+. For this purpose, 1 mL each dilution was added to 1 mL erythrocytes (2%), and then incubated for 2 h at room temperature. The formation of a smooth button at the bottom indicated nonreaction, whereas a rough granular deposition indicated a positive reaction, the intensity of which was determined based on the extent of deposition. The hemagglutination titer (HT) was calculated as the reciprocal of the dilution that produced one hemagglutination unit, the minimal dilution needed to cause complete agglutination.
Experimental animals and dietary treatments
This study was performed at the Intestinal Immunophysiology-Research Unit, Faculty of Medicine of Tunis (FMT), Tunis, Tunisia. Twenty young, adult male Wistar rats (Pasteur Institute, Tunis, Tunisia) weighing 60–80 g were acclimatized to the housing facilities for 7 days before being randomly assigned (10 animals/group) to the control or test group. Body weight and clinical observations were recorded daily before the start of the study. The test material, Beldia bean flour, was administered in a single intragastric dose of 300 mg made up to 3 mL with distilled water. 9,28,29 The control animals received an orogastric dose of 300 mg rodent pellet flour (lectin-free; Cereal Office, Tunis, Tunisia) suspended in 3 mL distilled water. Animals were housed individually in polypropylene cages under standard laboratory conditions in a pathogen-free environment with a temperature of 25 ± 1.4°C, a relative air humidity of 50 ± 5%, a 12-h light−12-h dark cycle, normal ventilation, and ad libitum access to water and food.
The duration of the experiment was 10 days. The diets were analyzed by the Food Technology Service (STA), National Institute of Nutrition and Food Technology (INNTA), Tunis, Tunisia. The nutrient analysis was performed as follows: nitrogen (crude protein = N × 6.25), fat, and sugar by AOAC, 30 and energy (kcal/100 g) by bomb calorimeter. Proximate analysis data are shown in Table 1. The study was approved by the Ethics Review Committee for Animal Experiments at the Faculty of Medicine of Tunis, and conducted according to European Community regulations concerning the protection of experimental animals. Groups were equalized with regard to body weight. Treatments and experimental conditions were implemented according to the recommendations of the Federation of the European Laboratory Animal Science Association (FELASA).
Data are shown as the percentage of 100 g of dried product from the mean of two duplicate determinations.
Collection of biological fluids from rats
On the 10th day of the experiment, in the morning, the animals were challenged by oral gavage with 300 mg rodent pellet flour suspension (control group) or with 300 mg Beldia bean flour suspension (BBFS) (experimental group). The rats were anesthetized with ether 2–3 h later. Each animal was placed on a heating pad and its rectal temperature was taken with a digital thermometer.
Upon verification of the loss of the pain reflex, a midline abdominal incision of ∼3–4 cm was made; the blood was obtained by cardiac puncture, and the small intestine was isolated by two ligatures (L1 and L2) located 7 and 18 cm from the pylorus. The segment was then placed on surgical gauze soaked in physiological saline, and the vascular supply of the intestine was kept intact. Then, 1.5 mL warm 140 mM Ringer's solution was infused into the segment using a calibrated syringe, and the intraluminal fluid was advanced through the segment by gently massaging the small intestine wall. Several seconds later, the content or the rinse liquids were recovered from the jejunal loop into a 5-mL test tube that had previously been chilled in ice water. A third ligature (L3) was placed near the first (L1) to isolate a new intact segment of the gut. The jejunal secretions (JSs) were collected by introducing 0.5 mL 300 mM Ringer hypertonic mannitol solution (570 mOsm/kg) into the segment delimited by ligatures, L2 and L3.
The intestine was returned to the abdominal cavity, and the abdomen was closed with wound clips. After 15 min, the loop was drained, and the content or the secretions were sampled. The jejunal rinse liquids (JRLs) and the JSs were cleared of insoluble material by centrifugation at 2683 × g for 10 min, and the supernatants were treated with sodium azide and stored at −20°C for further analysis. To prepare the ileal rinse liquids (IRLs) and ileal secretions (ISs), the ileum was subjected to the same procedure as the jejunum. Blood samples collected from the heart were left for 1 h at room temperature to aggregate the erythrocytes. Blood serum was purified by centrifugation at 241 × g for 10 min at 4°C and stored at −20°C.
Production of rabbit anti-PHA and anti-crude Beldia bean extract sera
Three rabbits kindly provided by the Central Animal House, the Faculty of Mathematical, Physical, and Natural Sciences (FST), Tunis, Tunisia, were used in this study. Two rabbits, R1 and R2, were immunized four times on days 1, 8, 14, and 29, with 100 μL of purified PHA (1 mg/mL; 100 μg PHA; Sigma Chemical Co.) emulsified with 500 μL complete Freund's adjuvant (CFA) and 400 μL distilled water. One rabbit, R3, was injected with 500 μL of crude Beldia bean extract (CBBE) mixed with 500 μL CFA on days 1, 6, 21, and 48 (Table 2). All of the rabbits were injected subcutaneously at several points on their back. Serum was separated from blood samples obtained by ear vein puncture, inactivated by incubation at 56°C for 30 min, and stored at −20°C until use. The rabbits were checked daily for signs of discomfort or local immunologic reactions. The immune responses of all of the animals were monitored by routine serological tests such as electrophoresis and immunoelectrophoresis (Table 3).
The immunization of rabbit R3 was shifted by 1 week from those of rabbits, R1 and R2.
CBBE, crude Beldia bean extract; PHA, phytohemagglutinin; R1, R2, and R3, the rabbits used for the immunization.
+, immunoserum subjected to analysis; −, immunoserum not subjected to analysis; R1, R2, and R3 denote the rabbits used for the immunization; Rn-IS day x, rabbit immunoserum sampled on day x.
Serological tests
Electrophoresis cellogel
Electrophoresis was performed with cellulose acetate membranes (Cellogel strips; SEBIA, Paris, France), which were incubated for 15 min in sodium veronal buffer (pH 9.2) and pre-electrophoresed for 10 min at 250 V before the rabbit serum was applied in a sample volume of 1.5 μL. The origin was 2 cm from the cathode. Subsequently, the strips were electrophoresed at 250 V for 35 min, stained sample side up for 5 min in amido black mixture, and destained for 10–15 min in a 50:10:40 mixture of methanol, acetic acid, and water. The membranes were dried for 30 sec in methanol and made transparent by immersion in an 87:12 mixture of methanol and acetic acid. The strips were then dried in an incubator at 80°C for 3 min on a clean glass plate. Densitometric tracing was performed with an implicit error of 0.18%. The implicit error of the electrophoresis was 0.2%.
Immunoelectrophoresis
For immunoelectrophoresis, glass microscope slides (7.5 cm long, 2.5 cm wide) were coated with 2 mL melted 1.5% agarose in veronal buffer (pH 8.6) (ionic force 0.05). The troughs were 1.3 mm wide, and the wells were punched with an 18-gauge needle attached to suction. After a prerun of 30 min, the wells were charged with the purified PHA (Sigma Chemical Co.) or the CBBE (0.5–1.0 μL/well) and subjected to electrophoresis at 400 V constant current for 50 min (panel meter), ∼1 mA per slide at room temperature in a Buchler immunoelectrophoresis chamber, with Whatman 3MM paper wicks. Troughs were cut out and filled with rabbit immunosera or the rat biological fluids (JSs, ISs, and sera). The slides were incubated for 20 h in a humid chamber at 36°C. The precipitation arc was observed using a down-up highlight source. The troughs were washed with 0.9% PBS for 20 h and with distilled water for 3 h, then dried in the incubator at 37°C, and stained with 0.1% (0.1 g/100 mL) amido black (in 45% methanol and 10% acetic acid solution).
Ouchterlony double immunodiffusion
Glass slides suitable for agarose gel diffusion studies were prepared using 15 mL 1.5% agarose (30 mg/mL) in 5 mL distilled water, buffered with 10 mL agar (pH 7.2), and heated to 100°C. Sodium azide (1%) was added as a preservative. A solution of warm agarose mixture (∼1 mL) was poured onto the slides to form a thin layer and allowed to gel. Wells with a capacity of ∼50 μL were formed using stainless steel cylinders, and the templates were withdrawn with forceps. For the diffusion experiments, the purified PHA preparation (1 mg/mL, PHA; Sigma Chemical Co.) or the CBBE (prepared from 300 mg bean flour diluted in 3 mL distilled water) was added to the center wells, and the test fluids (rabbit immunosera, rat intestinal secretions) were placed in the peripheral wells (other combinations are possible).
Slides were maintained in a wet room at ambient laboratory temperature (20–25°C) and observed twice daily. Precipitation bands, which usually began forming between 12 and 24 h, were readily visible. Amido Black, methanol, and acetic acid were used as the staining solution for further identification of the precipitation bands and for the permanent preservation of the slides. Staining was performed as follows: excess reactants were removed by washing with saline (containing 0.01 M phosphate buffer, pH 7.2) for ∼20 h and with distilled water for 3 h. The agarose was allowed to dry in an incubator overnight at 37°C. The dried plates were immersed in a solution of the dye (duration, 5–10 min), and excess dye was washed off with a solution of methanol, acetic acid, and water (5:1:4, v/v/v).
Counter-immunoelectrophoresis
Glass slides (2.5 × 7.5 cm) were covered with 3 mL 1.5% agarose prepared in veronal buffer (pH 8.6; ionic strength, 0.05). Cathodic wells were filled with protein antigens (purified PHA or CBBE), and the anodic wells were filled with rabbit immunosera. Slides were placed in an electrophoretic chamber containing veronal buffer (pH 8.6; ionic strength, 0.05) and run at 5 V/cm for 60 min at room temperature. The slides were examined by incident light for precipitation lines immediately after electrophoresis and again after overnight storage in a humid atmosphere at 4°C.
Competitive inhibition enzyme-linked immunosorbent assays
To detect the presence of PHAs in raw Beldia beans as well as in the rat biological fluids (intestinal rinse liquids, intestinal secretions, and sera), a competitive inhibition ELISA (CI-ELISA) was run according to Hajós et al. 31 with minor modifications.
ELISAs were performed in microtitration polystyrene plates (MaxiSorp, Nunc, Denmark). Purified PHA (Sigma Chemical Co.) was bound to each well by incubating 100 μL (1 mg/mL protein concentration) of 0.01 M PBS (pH 7.2) overnight at 4°C. The plates were washed four times with 0.5% Tween 20 in PBS (PBS-T). Unoccupied binding sites were blocked by incubating the plates for 2 h at 37°C with 100 μL of bovine serum albumin diluted at 3% with PBS and washed with PBS-T. Subsequently, 50 μL of rabbit anti-PHA serum diluted at 1:1500 and 50 μL of the different inhibitors (BBFS, intestinal rinse liquids, intestinal secretions, and sera) were added and incubated for 1 h at 37°C.
After six washes with PBS-T, 100 μL of peroxidase-conjugated goat anti-rabbit IgG (Sigma Chemical Co.) (1:500) was added and incubated for 1 h at 37°C. The plates were washed with PBS-T, and the reaction was revealed for 10 min in the dark at room temperature by the addition of 100 μL of o-phenylenediamine (Sigma Chemical Co.) dissolved to a concentration of 0.4 mg/mL in 0.1 M citric acid phosphate buffer (pH 5.0) containing 0.0120% H2O2. The reaction was stopped by adding 50 μL 2 N H2SO4, and samples were read at 492 nm in an ELISA reader (Axia Microreader, Biomérieux, France). All assays were performed in triplicate, and the results represent the mean of three determinations. The inhibition percentage in competitive ELISA was calculated using the following formula:
ODinhibitor (i = x), absorbance measured at μg/mL inhibitor concentration.
ODinhibitor (i = 0), absorbance measured at zero inhibitor concentration.
ODblank represents the wells showing nonspecific antibody binding, that is, binding observed in the absence of specific antibodies.
Statistical analysis
Statistical analysis was performed using the Mann–Whitney U-test for nonparametric data. All statistical calculations were performed using a computer-based, commercially available statistics package (Minitab data analysis software). P < .05 was considered significant.
Results
Characterization of PHA in raw Beldia beans
In a preliminary work, the presence of PHAs in raw Beldia beans was checked using a hemagglutination assay (HA), SDS-PAGE, immunoprecipitation, and CI-ELISA. The serological characterization of this lectin was performed using rabbit immunosera raised specifically against the purified PHA (Sigma Chemical Co.) and the CBBE. Thus, three rabbits were immunized: two with purified PHA and one with CBBE.
Hemagglutination assay
An HA is the best semiquantitative method to determine the presence of lectins in legumes. 32 As mentioned previously, the HT was defined as the reciprocal of the highest dilution that produced agglutination. The highest dilution of CBBE that produced agglutination was 1/512 for an HT of 512. These results demonstrate that bioactive PHAs are present in significant amounts in raw Beldia beans.
Electrophoretic analysis (SDS-PAGE)
The main goal of SDS-PAGE was to visualize a band with molecular weight of 33 kDa that corresponded to PHAs in the crude extract of the white Beldia beans. The purified PHA and phaseolin (Sigma Chemical Co.) were used as supplemental standard markers for migration (Fig. 1/lane 5). Their electrophoretic patterns revealed one major band at ∼33 kDa and two thin minor bands at ∼45 kDa. These subunits at ∼45 kDa are ascribed to phaseolin, an oligomeric protein consisting of different polypeptide subunits with molecular weights (Mw) between 43.10 and 51.50 kDa. 33 As shown in Figure 1/lane 1, the migration profile of the protein isolates from the Beldia bean extract displayed a number of polypeptide subunits of Mw between 92.40 and 14.40 kDa, with one migrant at 33 kDa, which might be assigned to PHA. This band is strong in the supernatant proteins extracted from the beans at pH 4.2 (Fig. 1/lane 2) and pH 5.5 (Fig. 1/lane 3). These results are consistent with the molecular weight of PHA reported in the literature. 34

Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) of crude Beldia bean (Phaseolus vulgaris L.) extract. Lane 1, total protein extracts from white Beldia beans. Lane 2, proteins extracted from beans at pH 4.2. Lane 3, proteins extracted from beans at pH 5.5. Lane 4, molecular mass markers: α-lactalbumin, 14.00 kDa; trypsin inhibitor, 20.00 kDa; carbonic anhydrase, 30.10 kDa; ovalbumin, 43.00 kDa; albumin, 67.00 kDa; and phosphorylase, 94.00 kDa. Lane 5, purified phytohemagglutinin (PHA), purified phaseolin.
Detection of PHA in raw Beldia beans by CI-ELISA
A CI-ELISA against the rabbit anti-PHA serum (R2-IS day 79) was performed using homologous PHA (control antigen) and a BBFS (test antigen). The inhibition curves illustrated in Figures 2 and 3 exhibit a total inhibition (100%) with homologous antigen (PHA) as well as with BBFS. These data clearly demonstrate the presence of PHA in the white bean seeds.
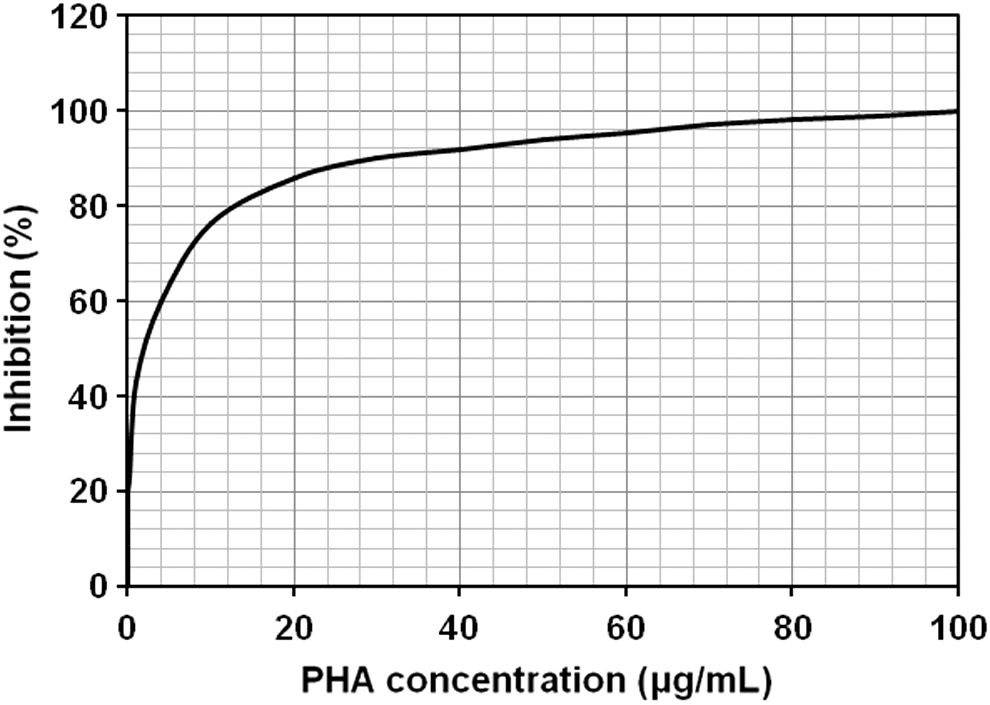
Competitive inhibition ELISA (CI-ELISA) against rabbit anti-PHA serum (R2-IS day 79) with homologous PHA concentration ranging from 0 to 100 μg/mL. The rabbit immunoserum was diluted at 1/1500. Data are presented as mean ± SD.
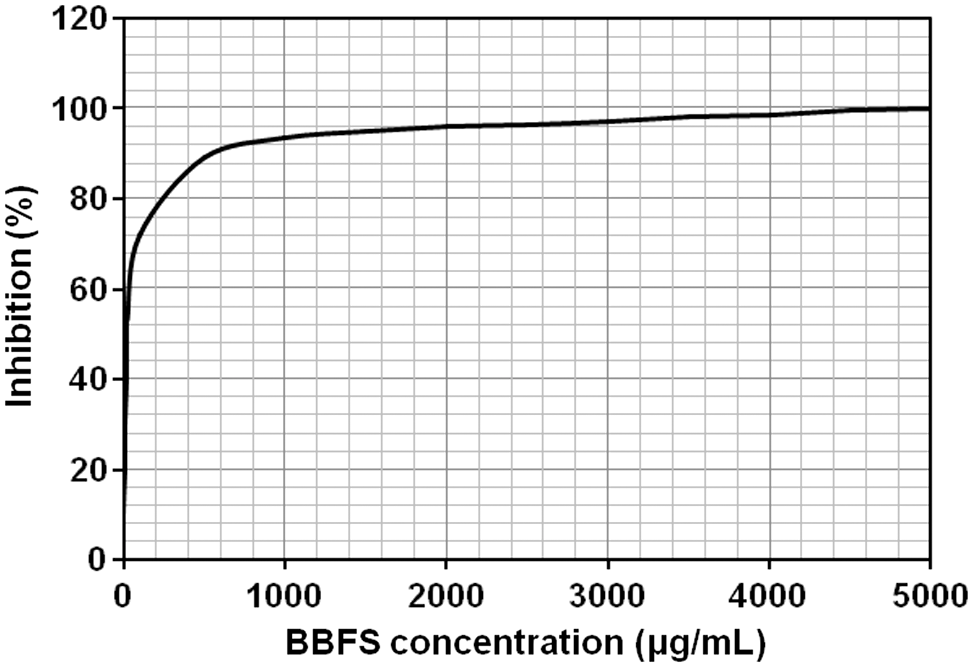
CI-ELISA against rabbit anti-PHA serum (R2-IS day 79) with Beldia bean flour suspension (BBFS) at concentrations ranging from 0 to 5000 μg/mL. The rabbit immunoserum (R2-IS day 79) was diluted at 1/1500. Data are presented as mean ± SD.
In summary, the examination of Beldia bean flour using the HA, SDS-PAGE, immunoprecipitation (following sections), and ELISA allowed us to demonstrate the existence of PHAs in fully biologically and functionally (agglutinant) active forms in the Beldia bean variety. Thus, the daily administration of 300 mg BBFS to experimental rats allowed direct contact between PHA and the mucosa of the small intestine.
Analyses of rabbit immunosera
Serum protein electrophoresis separates serum proteins into fractions of six distinct classes: albumin, α 1 globulins, α 2 globulins, β 1 globulins, β 2 globulins, and γ globulins. Approximately 60% of the total protein in the serum is albumin and the remaining fractions comprise mainly globulins, predominantly immunoglobulins (IgG, IgE, IgM, IgD, and IgA). The gamma fraction contains the greatest portion of immunoglobulins; hence, an increase in gamma globulin is referred to as gammopathy. Gammopathies can be either monoclonal or polyclonal. The most common increase in immunoglobulin levels is polyclonal and is due to immune system activity caused by infection or autoimmune disease. 35 When serum protein electrophoresis reveals a significant deviation from normal gamma globulin levels, a supplemental test, immunoelectrophoresis, should be ordered to identify the specific globulin(s) involved.
Analysis of rabbit R1 immunosera
As shown in Figure 4/electrogram R1, in rabbit R1, which had been immunized with purified PHA, the concentration of γ globulins was markedly increased compared with the nonimmunized animal. This result was confirmed by the results of the immunoelectrophoresis analyses. In contrast to the nonimmunized rabbit, all of the immunosera of R1 yielded distinct precipitation curves with the purified PHA and the antigenic determinant specific to PHA, which is present in the bean extract (Fig. 5/column R1). Figures 6/slide R1 and 7/slide R1 outline the results of Ouchterlony double diffusion and counter-immunoelectrophoresis, respectively. In contrast to the normal rabbit serum (NR-S), all of the rabbit R1 immunosera (R1-IS) produced distinct precipitin lines with the purified PHA and the PHAs in the CBBE.
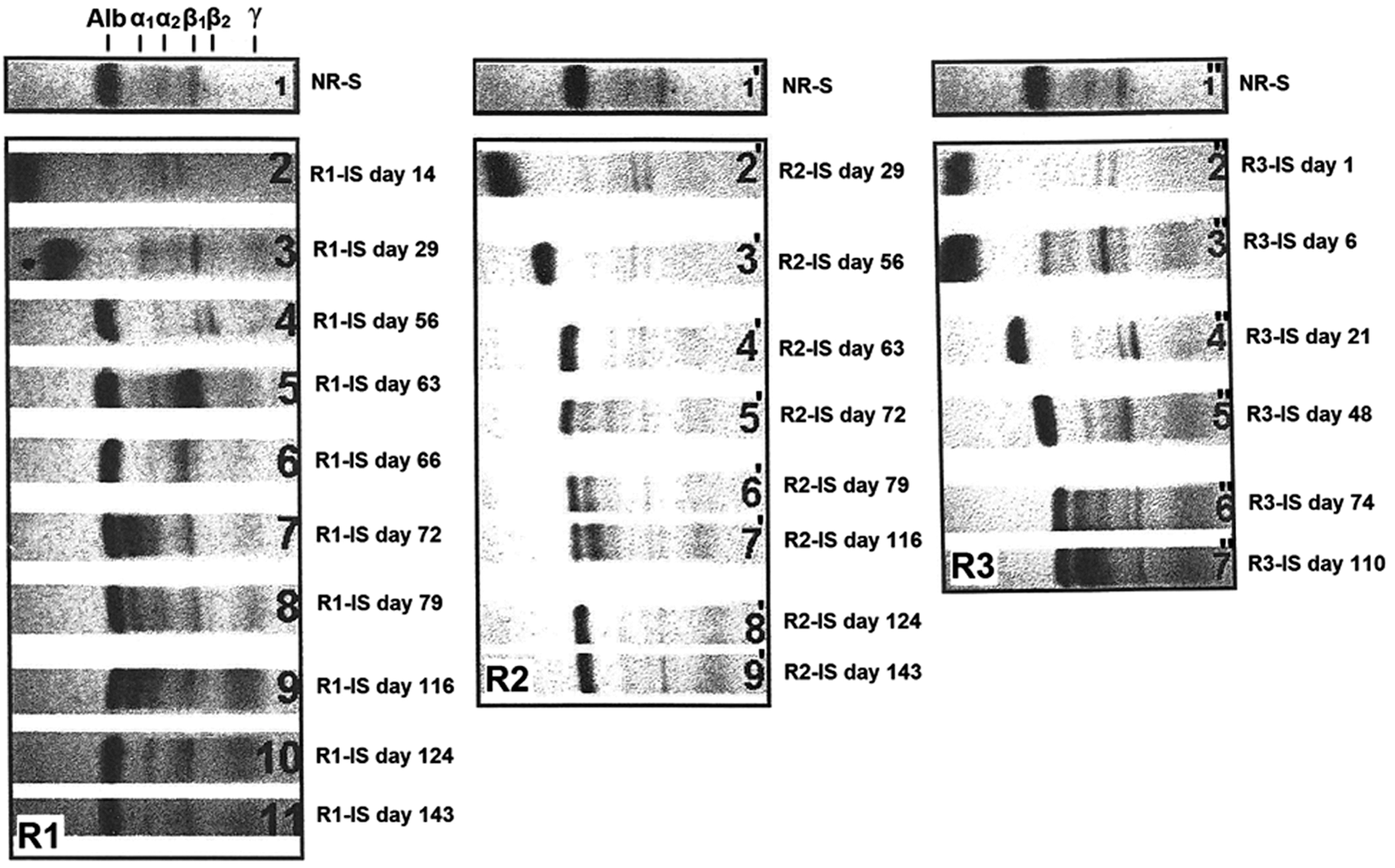
Analysis of the migration of rabbit sera by Cellogel electrophoresis. Rabbits, R1 and R2, were immunized with purified PHA, and rabbit R3 was immunized with crude Beldia bean extract (CBBE). The areas of migration of serum proteins, including albumin, α 1 and α 2 globulins, β 1 and β 2 globulins, and γ globulins, are indicated above the electrogram R1 site. The numbers n, n′, and n" indicate the track of the migration of the rabbit immunosera (R1-IS, R2-IS, and R3-IS) withdrawn at different points of time. R1, R2, and R3 denote the electrogram numbers.
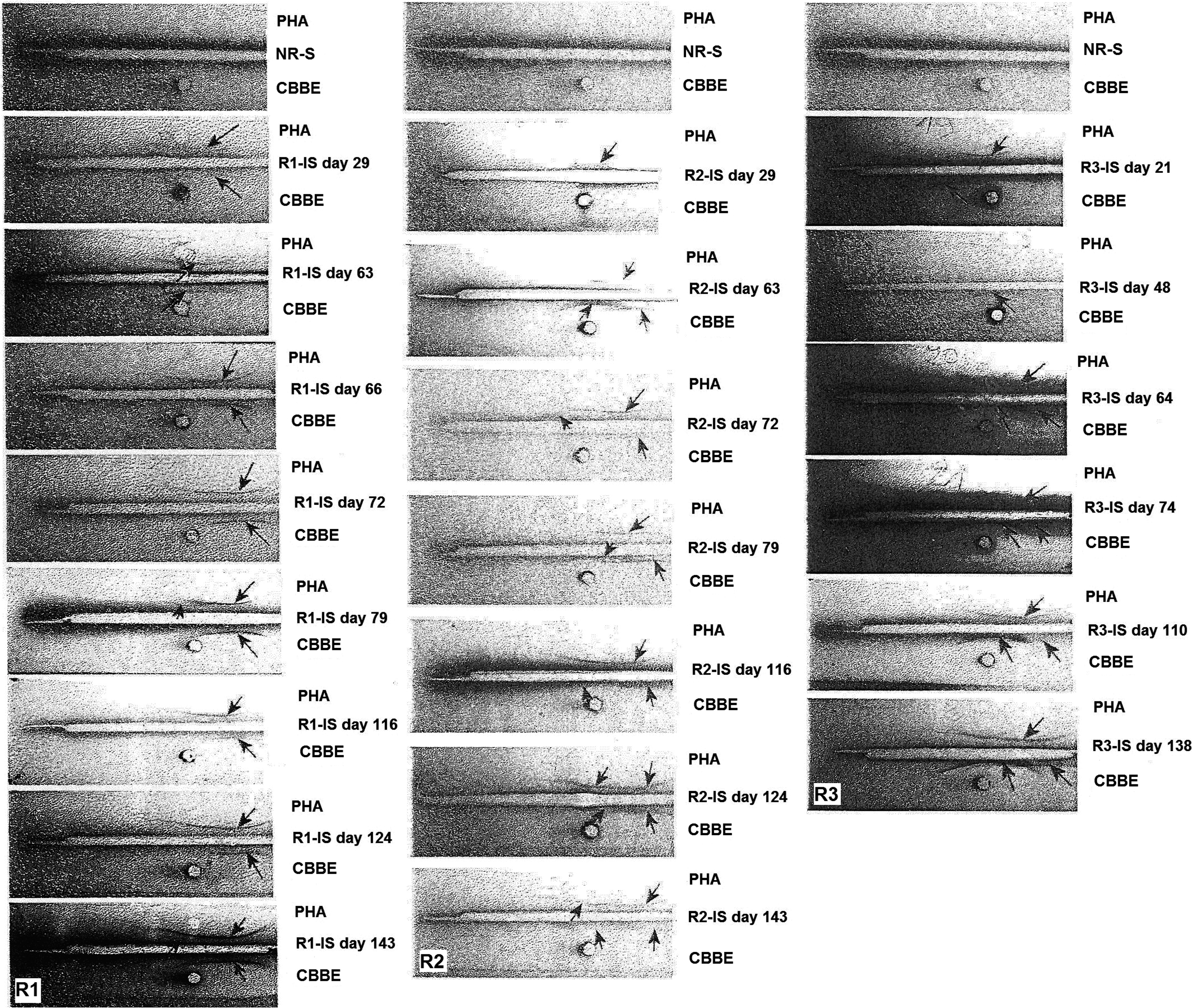
Immunoelectrophoretic patterns showing the precipitation curves of purified PHA and CBBE subjected to rabbit immunosera (R1-IS, R2-IS, and R3-IS). The precipitation curves indicate the presence of specific anti-PHA antibodies. Precipitin bands were visible with PHA and CBBE in all troughs, except the troughs containing normal rabbit serum (NR-S). Troughs were filled with NR-S (top troughs) and rabbit immunosera (remaining troughs); antigens in the wells are shown as purified PHA, CBBE, and NR-S. The arrows indicate the precipitin bands; R1, R2, and R3 denote the column numbers of the glass slides.
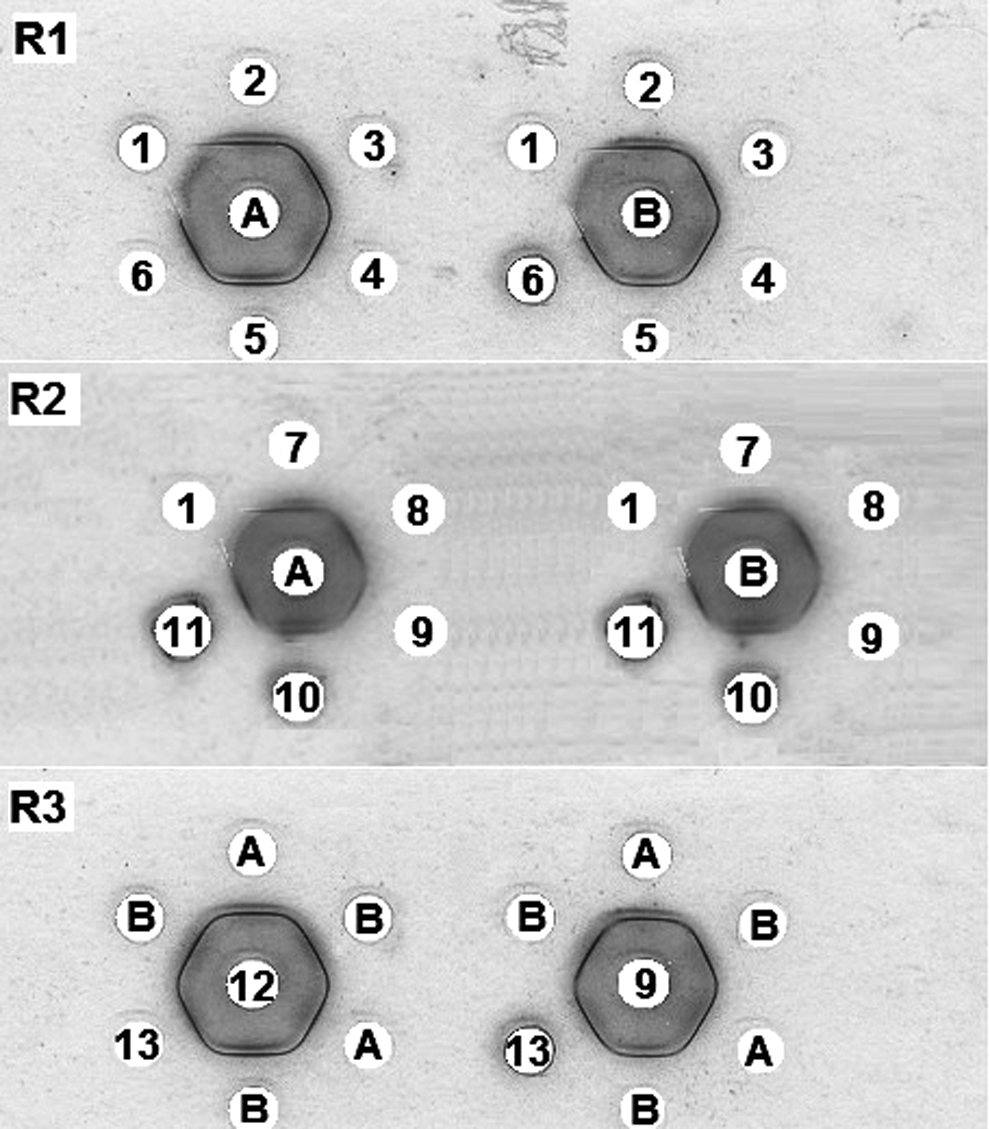
Double immunodiffusion showing precipitation reactions. The wells contained the following: A, purified PHA; B, CBBE; 1, NR-S; 2, rabbit R1 immunoserum day 72 (R1-IS day 72); 3, rabbit R1 immunoserum day 79 (R1-IS day 79); 4, rabbit R1 immunoserum day 116 (R1-IS day 116); 5, rabbit R1 immunoserum day 124 (R1-IS day 124); 6, rabbit R1 immunoserum day 143 (R1-IS day 143); 7, rabbit R2 immunoserum day 63 (R2-IS day 63); 8, rabbit R2 immunoserum day 72 (R2-IS day 72); 9, rabbit R2 immunoserum day 79 (R2-IS day 79); 10, rabbit R2 immunoserum day 116 (R2-IS day 116); 11, rabbit R2 immunoserum day 124 (R2-IS day 124); 12, rabbit R3 immunoserum day 72 (R3-IS day 72); and 13, intestinal secretions of BBFS-gavaged rats. R1, R2, and R3 denote the glass slide number.
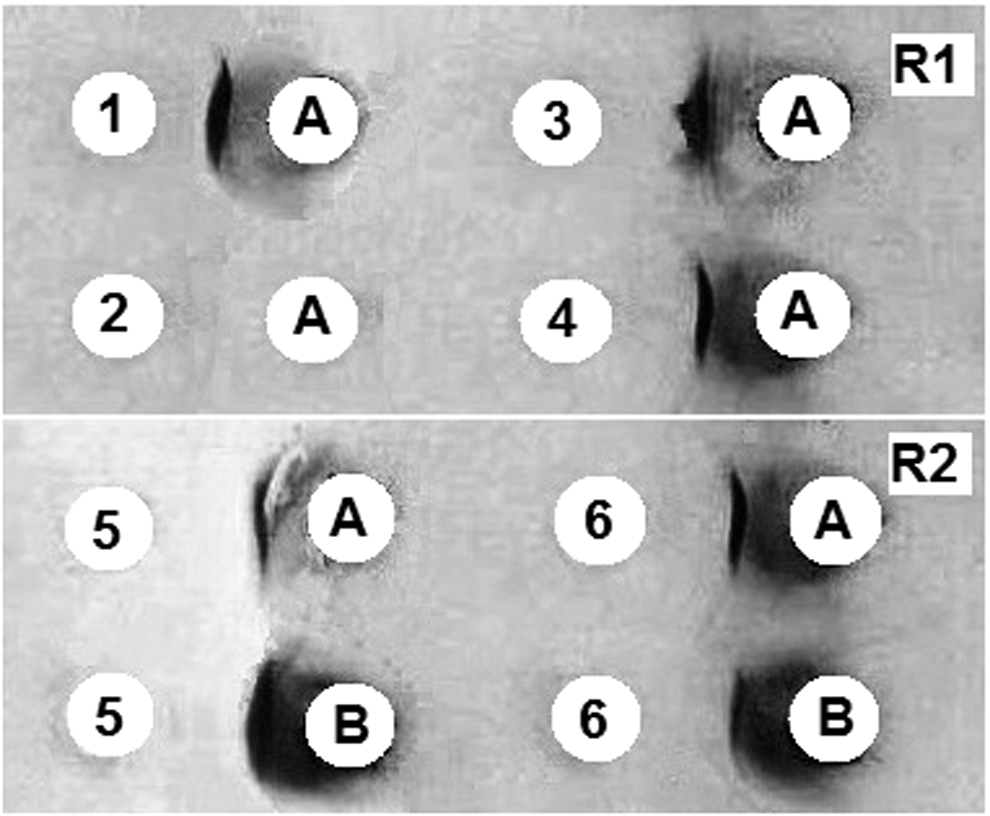
Counter-immunoelectrophoretic patterns showing precipitation curves of purified PHA and CBBE subjected to rabbit antisera. A, Purified PHA; B, CBBE; 1, rabbit R1 day 124 (R1-IS day 124); 2, NR-S; 3, rabbit R1 immunoserum day 79 (R1-IS day 79); 4, rabbit R1 immunoserum day 116 (R1-IS day 116); 5, rabbit R2 immunoserum day 116 (R2-IS day 116); 6, rabbit R2 immunoserum day 124 (R2-IS day 124). R1 and R2 denote the glass slide number.
Analysis of rabbit R2 immunosera
As shown in Figure 4/electrogram R2, rabbit R2 produced very similar results to those obtained from rabbit R1. Following immunization with purified PHA, R2 exhibited a substantial increase in the γ-globulin level in the electrophoretic profiles in sampling performed on day 29. No bands formed using the serum of the nonimmunized rabbit or R2 before immunization. The immunoelectrophoresis test showed complete identity between the purified PHA and the CBBE for all specific R2 immunosera (R2-IS) drawn from day 29 onward (Fig. 5/column R2). The bands obtained from CBBE were strong matches to PHA. The recognized antigen in the bean extract is most likely PHA. The same results were obtained with the Ouchterlony and counter-immunoelectrophoresis tests (Figs. 6/slide R2 and 7/slide R2, respectively).
These findings suggest that rabbits, R1 and R2, synthesized anti-PHA antibodies following their immunization with PHA. Therefore, intradermally administered PHA is immunogenic in rabbits. Together, these data suggest that BBFS contains immunogenic PHAs that are likely to have an effect when administered orally.
Analysis of rabbit R3 immunosera
Electrophoresis of immunosera from R3, which was immunized against CBBE, also revealed an increase in γ globulins compared with that of the control rabbit. This shift is indicative of chronic inflammation of dietary origin (Fig. 4/electrogram R3). This observation is strongly correlated with immunoelectrophoresis findings showing the precipitation curve with the crude bean extract (Fig. 5/column R3). These immunosera also recognized PHAs. The total identity of the reaction obtained in the immunodiffusion double gel agarose (Fig. 6/slide R3) between PHA and the antigen present in the Beldia extract strongly supports that this antigen recognized by R3 immunosera (R3-IS) (day 72) is PHA.
Analyses of biological fluids of the BBFS-gavaged rats
JRL and IRL were recovered from their respective loops several seconds after the injection of 1.5 mL 140 mM Ringer's solution. These fluids correspond to the intraluminal content of the small intestinal mucosa 3 h and 30 min after the last gavage. Intestinal secretions were withdrawn 4 h and 30 min after the last gavage and most likely correspond to intraluminal secretions. Sera were obtained by cardiac puncture just after the collection of intestinal secretions.
Analysis of JRL and IRL
ELISA is an effective and sensitive technique for determining the presence of lectins, both free in the digestive tract and bound to the gut. 36 The presence of intraluminal PHA in the rinse liquids of jejunal (JRL) and ileal (IRL) loops was tested by competitive inhibition using rabbit anti-PHA serum. The rinse liquids of the jejunal and ileal loops obtained from control and experimental rats were taken on day 10, pooled, and then tested using ELISA. Total inhibition of binding by rabbit anti-PHA (100%; P < .05) was observed in the rinse liquids of the jejunal loop from rats gavaged with BBFS. This result strongly suggests that PHA remained intact in the jejunum of experimental rats 3 h and 30 min after the last gavage with the raw ground beans (Fig. 8). In contrast, no inhibition was observed with the rinse liquids of the ileal segment (IRL; P > .05). Based on these observations, it can be concluded that PHA was present in a fully immunogenic form in the jejunum, but not in the ileum. The absence of proteins of dietary origin from the ileum has been extensively described in the literature. 37
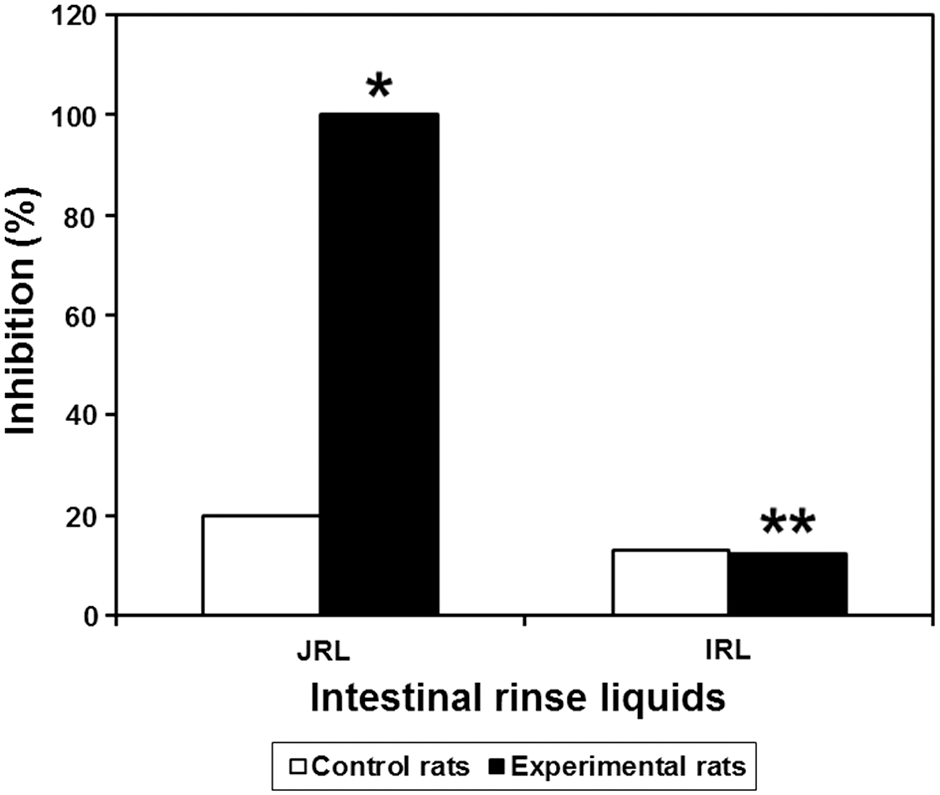
CI-ELISA against rabbit anti-PHA serum with the rat jejunal rinse liquid (JRL) and ileal rinse liquid (IRL). The rabbit anti-PHA serum used here was R2-IS day 79 diluted at 1/1500, and the intestinal rinse liquids were diluted to 1/2. Data are the mean percent inhibition value ± SD. *Significantly different from the corresponding control value, P < .05. **Not significantly different from the corresponding control value, P > .05.
Analysis of secretions in the small intestine
The presence of native PHA in the intestinal secretions of BBFS-treated rats was checked using double immunodiffusion. The total identity of the reactions between purified PHA and the secretions with rabbit anti-CBBE serum established the presence of PHA in the intestinal secretions (Fig. 6/slide R3). This result is supported by the results of the CI-ELISA. The JS and the IS were collected 4 h and 30 min after the last gavage and pooled for the control and experimental rats. Inhibitions of ∼36.40% and 14.50% were recorded for the JSs (P < .05) and ISs (P < .05), respectively; however, the intestinal secretions of the control rats did not display any significant inhibition (Fig. 9).
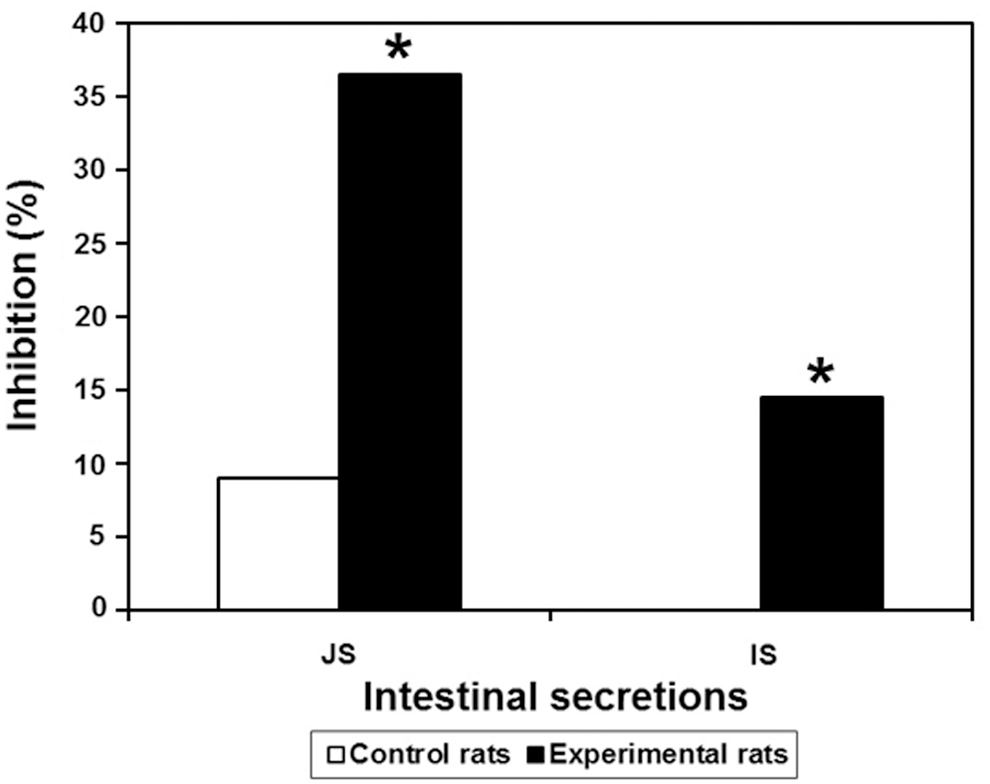
CI-ELISA against rabbit anti-PHA serum with the rat jejunal secretion (JS) and ileal secretion (IS). The rabbit anti-PHA serum used here was R2-IS day 79 diluted to 1/1500, and the intestinal secretions were diluted to 1/2. Data are the mean percent inhibition value ± SD. *Significantly different from the corresponding control value, P < .05.
Analysis of rat sera
The rat sera were analyzed using several serological techniques, including Ouchterlony double immunodiffusion, immunoelectrophoresis, and ELISA.
Analysis of sera with Ouchterlony and immunoelectrophoresis
As shown in Figure 10, no bands appeared in the agar gel immunodiffusion for the sera of BBFS-gavaged rats, the rabbit anti-PHA serum (R2-IS day 79), and the rabbit anti-CBBE serum (R3-IS day 74). This result could be explained either by the complete absence of PHA in the rat sera or by a very low quantity in the latter, below the limits of sensitivity of this technique. The sera of the control and experimental rats were analyzed as well using immunoelectrophoresis, with the sera as a source for antibodies and purified PHA and CBBE as antigens. No immunoprecipitates were formed, suggesting the absence of anti-PHA antibodies and/or the absence of antigens from the Beldia extract (photograph not shown).

Ouchterlony double immunodiffusion. The center wells contained the following: a, rabbit anti-PHA serum (R2-IS day 79); b, rabbit anti-CBBE serum (R3-IS day 74). The outer wells contained: A, purified PHA; B, CBBE, c, serum of rats gavaged with BBFS.
CI-ELISA against rabbit anti-PHA serum with rat sera
To compare their systemic anti-CBBE immune responses to those of BBFS-gavaged rats, growing rats were subcutaneously immunized against CBBE. Serum samples were obtained on days 7 and 36 from the immunized rats and on day 10 from the Beldia-gavaged rats. The CI-ELISA against rabbit anti-PHA serum (R2-IS day 79) exhibited 36% inhibition on day 7 (P < .05) and 40.50% inhibition on day 36 with sera from rats immunized subcutaneously (Fig. 11; P < .05). As shown in Figure 11, the sera derived from gavaged rats inhibited this binding by only 12.50% (P < .05), showing a very weak systemic immune response on day 10.
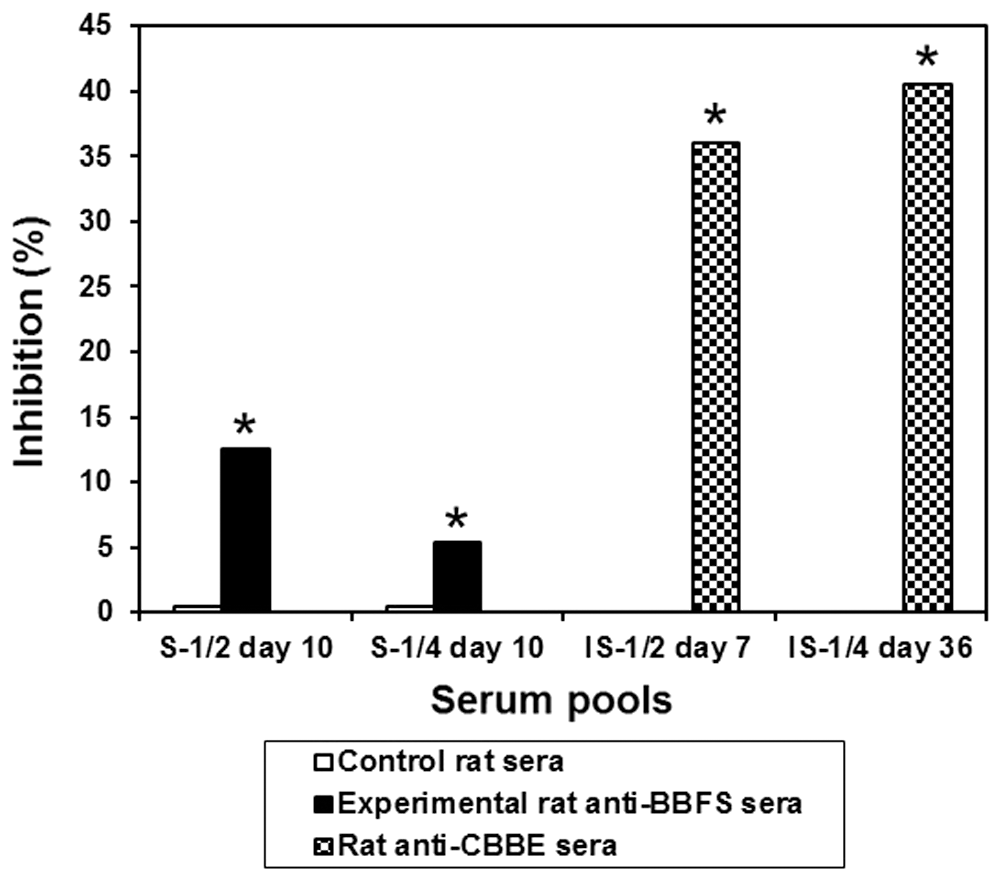
CI-ELISA against rabbit anti-PHA serum with serum pools of control rats, experimental rats gavaged with BBFS, and sera from rats immunized against CBBE. The rabbit anti-PHA serum used here was R2-IS day 79 diluted to 1/1500; serum pools of control and experimental rats collected on day 10 were tested at 1/2 and 1/4 dilutions, respectively (S-1/2 day 10, S-1/4 day 10); rats anti-CBBE sera were drawn on days 7 and 36 after the first injection and diluted to 1/2 and 1/4, respectively (IS-1/2 day 7, IS-1/4 day 36). Data are the mean percent inhibition value ± SD. *Significantly different from the corresponding control value, P < .05.
In conclusion, the current findings indicate that the degree of inhibition of rabbit anti-PHA serum binding corresponds to the presence of PHA or of anti-PHA antibodies in biological fluids such as intestinal rinse liquids, intestinal secretions, and sera. In the intestinal rinse liquids and secretions, the existence of PHA was determined by the Ouchterlony test and the absence of anti-PHA antibodies was demonstrated using immunoelectrophoresis (data not shown). It can be argued that these results support competitive inhibition for PHA and not for secretory anti-PHA antibodies.
Discussion
The presence of PHAs in the small intestine after the ingestion of white kidney beans (P. vulgaris L.) has received scant attention in the literature, which is surprising given that white beans belong to widely consumed varieties and that multiple studies have addressed the effect of purified PHAs or red kidney beans on the small bowel. Our immunological investigations clearly demonstrated the presence of intraluminal PHA in the JRLs as well as in the JSs and ISs of experimental rats 3–4 h after the last oral administration of the 300 mg BBFS.
Several hypotheses can be proposed regarding the presence of PHA in the lumen of the small intestine. The first is that native PHA could be present in a free and fully reactive form inside the intestinal lumen 3 h and 30 min after gavage due to its strong resistance to the digestive proteolytic enzymes. The partial digestion of dietary PHA might also explain its persistence in the lumen.
The second is that the PHA could be bound to the epithelial microvilli glycocalyx or to the receptors and/or specific antibodies. Indeed, the intestinal plasmocytes may release anti-PHA antibodies (predominantly IgA) in response to the oral administration of PHA. The latter antibodies could cross the epithelium and permeate the mucus blanket covering the intestinal mucosa. Accordingly, a specific antigen–antibody interaction could be established between the intraluminal PHA and IgA. Indeed, competitive inhibition between IgAs in the intestinal rinse liquids and intestinal secretions cannot be ruled out. Direct light microscopy showed a striking increase in the number of goblet cells in the small intestinal villi, whereas electron microscopy revealed a substantial release of mucins in the mucosa of Beldia bean-gavaged rats. 26 These observations, coupled with the stimulation of water secretion, 24 suggested an intestinal defense reaction against an antigen of dietary origin, presumably primarily PHA.
The third is that the PHA in the intestinal secretions could have resulted from an exudation; that is, the glycoproteins were secreted from the basolateral side of the enterocytes into the lumen through the systemic circulation. This mechanism is well recognized in celiac disease and inflammatory bowel syndrome. 38 PHA, known for mitogenic properties and its interaction with the enterocyte microvilli, 28,29,39 has been implicated in morphological changes in the jejunal and ileal mucosa of Beldia bean-fed rats relative to control rats (increase in crypt depth, villi height, and mitotic index). 26 The daily administration of a single bolus of 300 mg of BBFS by gavage to rats for 10 consecutive days did not appear to be accompanied by the inhibition of bean protein tolerance.
Indeed, compared with the sera of rats immunized against the CBBE, the sera from Beldia bean-gavaged rats showed very weak competitive inhibition based on ELISA (12.50%). This finding could be ascribed either to the transcellular passage of intraluminal PHA to the bloodstream of the animals 40 or to the presence of anti-PHA antibodies. One argument in favor of the first hypothesis is the absence of PHA from the rinse liquids of the ileal loops [very low inhibition (12.16%) by the rabbit anti-PHA serum on ELISA] and its detection 1 h later in the ISs (Ouchterlony and ELISA data). In the second case, the systemic anti-PHA immune response of gavaged rats is much lower than that generated by conventional subcutaneous immunization. This finding is consistent with the mechanisms of oral tolerance described for the antigens of dietary origin.
The regulatory mechanisms of dietary antigen tolerance are numerous, complex, and poorly understood. They involve several agents, such as gut commensal flora and lymphocyte populations, especially MAIT lymphocytes (mucosal-associated invariant T lymphocytes). Under experimental conditions, the gavage of animals with 300 mg BBFS over 10 subsequent days associated with standard feeding did not appear to cause dysregulation of oral tolerance to bean proteins.
Several studies have shown that the oral intake of raw kidney bean (P. vulgaris L.) induces morphometric and functional alterations in the intestinal mucosa. 8,41,42 These alterations are mainly the result of reactions to PHAs. In the first part of this work, P. vulgaris lectins were immunobiochemically characterized in the bean flour suspension used for the rat gavage. The results clearly showed that raw Beldia beans contain biologically active PHAs. Preliminary evidence from related morphological studies 26 indicates that the alterations to the intestinal mucosa are most likely linked to the interaction of PHA with the epithelial cells lining the small intestine, resulting in the stimulation of nonspecific defense mechanisms such as a significant increase in goblet cell numbers and an associated increase in mucus discharge and the hyperproliferation of crypt cells known to be involved in intestinal secretion (noticeable increases in water and electrolyte secretion were detected in response to administration of Beldia bean flour). 26 Consistent with this stimulation of nonspecific defense mechanisms, the inhibition observed by ELISA using sera from gavaged rats could have resulted either from an anti-PHA immune response accompanied by the transepithelial transfer of PHA or one immunogenic fragment or from the presence of PHA of dietary origin in the serosal compartment. The first case would be associated with an increase in the oral tolerance to PHA most likely related to the modification of the intestinal permeability. The second case suggests the development of standard oral tolerance to food antigens combined with the transport of PHA from the lumen of the small intestine to the systemic compartment.
Dietary lectins, including both wheat germ agglutinin 43 and PHA, 44,45 have been shown to cross the gastrointestinal barrier rapidly and enter the peripheral circulation. To date, no PHA studies in humans have been conducted; however, in laboratory animals, PHA easily crosses the gut barrier and enters the bloodstream, where it may travel to many organs and tissues and disrupt normal cell function and cause disease. 40,46 In rats dosed with PHA, up to 10% of the lectin is found in the circulation 3 h after feeding. 44,45 Thus, PHA is a powerful oral immunogen that produces a high titer of IgG anti-PHA antibodies in animals and most likely man. 14 The anti-PHA antibodies reach measurable levels 10 days after the first dose, and further feeding or reintroduction results in booster effects. 14 Thus, the gut anti-lectin IgA system is ineffective against PHA because it cannot prevent its absorption after reintroduction. 45 This abrogation of the gut IgA response to PHA and possibly to other lectins has important consequences for autoimmune disease because it allows dietary lectins continuous access to T lymphocytes at the gut mucosal surface.
Human and animal tissue experiments reveal that PHA and other food lectins can cause a leaky gut and can enter the circulation. 40,43,46 –51 A leaky gut is one of the first steps implicated in many autoimmune diseases. 52 Impaired intestinal integrity induced by dietary lectins may also cause low-level inflammation in the bloodstream, which is a precursor to developing atherosclerosis and cancer. 14,53 –55 In addition to kidney beans and other P. vulgaris varieties, all other legumes contain lectins with varying degrees of toxicity, ranging from mild to lethal. Soybean lectin (SBA) is also known to impair intestinal permeability and cause a leaky gut. 51,56 Peanut lectin (PNA) is the only legume lectin to have been tested in living humans by Dr. Rhodes's research group in London. Within less than an hour after ingestion by healthy normal subjects, PNA entered their bloodstreams, where it can interact with virtually every cell in the body, with potentially detrimental effects. 50 The lectins found in peas (PSA) and lentils (LCA) seem to be much less toxic than PHA, SBA, or PNA, but they are not completely without adverse effects based on tissue and animal experiments. 57 –60 Unfortunately, no long-term lectin studies have ever been conducted in humans. Nevertheless, based on animal and tissue studies, it is known that these antinutrients damage the intestinal barrier, impair growth, alter normal immune function, and cause inflammation.
Several aspects of this system remain to be elucidated: the transepithelial transfer of PHA (using HPLC, ELISA, and RIA); local and systemic humoral anti-PHA immune responses under different experimental conditions; and the role of some GALT lymphocyte subpopulations (gut-associated lymphatic tissue) in the regulatory mechanisms of immune tolerance to food antigens, especially of MAIT.
Conclusions
This work demonstrated that daily gavage of rats with raw white Beldia kidney beans (P. vulgaris L.) for 10 subsequent days resulted in the retention of fully immunoreactive PHAs in the jejunum 3–4 h after the last challenge with the BBFS. These findings suggest that when Beldia seeds are consumed raw or undercooked, the PHAs act as metabolic signals. They can also alter the glycosylation state of the intestinal epithelium through binding to pre-existing membrane glycosyl groups and thus further amplify their potent physiological effects.
Footnotes
Acknowledgments
The authors are grateful to Prof. Khaled Zarrouk, Head of the Food Technology Service (STA), National Institute of Nutrition and Food Technology (INNTA), Tunis, for his technical assistance in analysis of the rat diet. The authors wish to give equal thanks to Prof. Mohamed Boussaïd, Department of Chemical and Biological Engineering, National Institute of Applied Science and Technology (INSAT), Tunis, for his kind help regarding the extraction of proteins from white beans. The authors are indebted to Dr. Lotfi Bitri, Professor in Animal Physiology, Faculty of Mathematical, Physical, and Natural Sciences (FST), Tunis, for his expertise in animal care and rabbit immunization. This work was financially supported by the Intestinal Immunophysiology-Research Unit (02/RU/09–02), Faculty of Medicine of Tunis, University of Tunis El Manar, Tunis, Tunisia.
Author Disclosure Statement
No competing financial interests exist.