
Abstract
In this study, we investigated the hepatoprotective effects of ethanol extracts from Chlorella vulgaris (CH) on animals. We measured its effect on the quinone reductase (QR) activity in Hepa1c1c7 cells, finding that CH induced a significantly higher QR activity in these cells. We isolated the active fraction (CH F4-2) from CH using chromatography methods. CH F4-2 may activate cellular antioxidant enzymes through upregulation of the Nrf2 pathway in hepatocarcinoma cells with CH F4-2 (25.0–200 μg/mL) for 48 h. Furthermore, CH F4-2 increased the expression of NQO1 [NAD(P)H: quinone oxidoreductase, also known as QR], heme oxygenase-1, and glutathione-S-transferase P. Moreover, we found that ethanol-induced hepatic pathological changes—elevations in glutamic oxaloacetic transaminase, glutamic pyruvic transaminase, γ-glutamyltransferase, and lactate dehydrogenase—were significantly decreased. The inhibitory effect of CH on alcohol-induced liver injury was associated with the suppression of alcohol-induced increases in intestinal permeability. The ethanol extract from CH was found to induce QR activation, making it a potentially good candidate for a hepatoprotection agent.
Introduction
L
Imbalance antioxidants in the cells were found to cause dysfunction in the cellular membrane system proteins and DNA oxidations and eventually cell injury. 7 An alternative and more effective strategy for combating the toxicity of ROS has recently emerged through the study of inducers of phase II detoxification enzymes. Another proposed hepatoprotective mechanism has been related to the capacity to induce phase II enzymes during the detoxification process. 8 Upregulation of phase II detoxification enzymes such as quinone reductase (QR) is thought to be a useful biomarker for anticarcinogenesis. 9 The QR assay is used as a simple screening method to detect the capacity of a compound to act as a potential hepatoprotection and anticarcinogen. 10
Chlorophyll is among the most widespread and numerous groups of biopolymers. 11 An increasing number of studies have indicated that many chlorophyll extracts from green plants, green algae, and microorganisms possess potent antioxidant abilities. 12 They have been recognized as having interesting clinical properties, such as anti-inflammatory, antibacterial, antiviral, antihistaminic, antioxidant, and antitumoral effects. 13 They can also increase the function of the endogenous antioxidant enzyme systems. 14 Moreover, chlorophyll and its water-soluble salts (chlorophyllins) have been proposed for their hepatoprotective effect. 13 Furthermore, their antioxidant effects may be a result of a combination of radical scavenging and interaction with enzyme function. 15
The objective of this study was to determine expression phase II enzyme, induction phase II enzyme, and QR activity of an ethanol extract and chlorophyll from chlorella, Chlorella vulgaris in hepatocarcinoma cells. Moreover, an ethanol extract and chlorophyll were separately administered to reduce ethanol-induced liver injury in rat. Four weeks after the oral administration of ethanol, the liver and blood of rats were evaluated to determine whether the animals had been protected against hepatotoxicity. It was also thought that the information obtained would shed new light on the mechanism of action of ethanol extract and chlorophyll from C. vulgaris.
Materials and Methods
Extraction of ethanol extract from C. vulgaris
C. vulgaris was purchased from Daesang Corp. (WellLife, Seoul, Korea) and lyophilized at−80°C using a freeze-dryer. One hundred grams of lyophilized C. vulgaris powder was mixed with 500 mL 99.9% ethanol and stirred at room temperature for 24 h. The supernatant obtained was filtered (with Whatman No. 1), then evaporated by a rotary evaporator at 40°C, and freeze-dried. Dried ethanol extract was isolated with gel chromatography and high performance liquid chromatography (HPLC). The active fraction was purified using the LH-20 sephadex (Sigma-Aldrich, St. Louis, MO, USA) column, which was eluted with methanol at a flow rate of 1.5 mL/min. The isolated fraction from the LH-20 column was purified on HPLC using a C18 ODS column (4.6×250 mm, 5 μm; Waters, Milford, MA, USA) at a flow of 1.0 mL/min. The purified fraction was monitored at 660 nm on a diode array detector. Finally, the purified active fraction was compared with commercial chlorophyll a (Sigma-Aldrich) on HPLC.
Cell culture
Hepa1c1c7 cells were obtained from the American Type Culture Collection (ATCC CRL-2026, Rockville, MD, USA), and the cells were maintained at subconfluence in a humidified atmosphere of 5% CO2 at 37°C. Dulbecco's modified Eagle's medium (Hyclone, Logan, UT, USA) was used for Hepa1c1c7 cells. Media were supplemented with 10% fetal bovine serum (Hyclone) and 1% of 100 U penicillin/streptomycin (Hyclone).
Cell viability
The cytotoxicity of CH and its fractions were evaluated using the EZ-Cytox Cell viability assay kit (tetrazolium salt; Daeillab, Seoul, Korea). In brief, Hepa1c1c7 cells (1×105 cells per well) were plated onto 96-well tissue culture plates, incubated at 37°C for 24 h, and the media were changed. Then, the cells were treated with various concentrations of CH and its fractions. After 24 h, 200 μL of tetrazolium salt solution was added to the wells and incubated at 37°C for another 1 h. The absorbance at 450 nm was measured with a multi-microplate reader (Infinite® 200 PRO; Tecan, Männedorf, Switzerland).
QR activity
The QR activity was measured as described previously, 16 with minor modifications. Briefly, Hepa1c1c7 cells (1×105 cells per well) were plated onto 96 wells and incubated for 24 h before treatment. Media contained various concentrations of the CH, and those fractions were added and incubated for an additional 24 h. As a positive control, 5 μM of sulforaphane (SFN; Calbiochem, Darmstadt, Germany), a monofunctional inducer of QR, was used. Treated cells were rinsed with phosphate buffer pH 7.4, lysed with 50 μL 0.8% digitonin in 2 mM EDTA, incubated, and agitated for 30 min. A 200 μL aliquot of mixed solution (24.5 mL of 25 mM Tris buffer; 25 mg of bovine serum albumin; 0.5 mL of 1.5% Tween-20 solution; 0.5 mL of thawed cofactor solution [92.7%, 150 mM glucose-6-phosphate; 6.15%, 4.5 mM NADP; 1.14%, 0.75 mM flavin adenine dinucleotide in Tris buffer]; 150 U of glucose-6-phosphate dehydrogenase; 7.5 mg of MTT (3-(4,5-dimethylthiazo-2-yl)-2,5-diphenyltetrazolium bromide); and 75 μL of 50 mM menadione in acetonitrile) was added into lysed cells. Readings were made at five time points, 50 sec apart, using a multi-microplate reader (Infinite 200 PRO; Tecan) at 610 nm. The protein content was also measured using the cell lysate according to the Bradford method, by using a Bio-Rad Protein Assay kit (Bio-Rad, Seoul, Korea). The induction of QR activity was expressed as the concentration required to the double QR activity (CD) value and as a concentration required to double QR-specific activity.
RNA extraction, cDNA synthesis, and polymerase chain reaction
All experiments were performed in differentiated L6 myotubes after 7 days. All RNAs in cell cultures were extracted with the Trizol reagent (Sigma-Aldrich), and 1 μg of total RNA was reverse-transcribed by using oligo-dT as a primer in 20 μL reverse-transcription solutions (Promega, Madison, WI, USA). The reverse transcription-polymerase chain reaction (PCR) conditions for NQO1, heme oxygenase-1 (HO1), and glutathione-S-transferase P (GST-P) were as follows: initial denaturation at 95°C for 1 min, 1 min of annealing time at 60°C, and 1 min of amplification time for 35 cycles after an activation step of 2 min at 95°C. PCR products were analyzed by electrophoresis on a 1% agarose gel. The primers used in this study were as follows: HO1 forward, 5′-GAGCCTGAATCGAGCAGAAC-3′; reverse, 5′-AGCCTTCTCTGGACACCTGA-3′; NQO1 forward, 5′-TTCTCTGGCCGATTCAGAG-3′; reverse, 5′-GGCTGCTTGGAGCAAAATAG-3′; GST-P forward, 5′-GGATGG AGACCTCACCCTTT-3′; reverse, 5′-CAGGGCCTTCACGTAGTCAT-3′; and GAPDH forward, 5′-GGCATTGCTCTCAATGACAA-3′; reverse, 5′-TGTGAGGGAGATGCTCA GTG-3′.
Protein extraction and western blot analysis
Extraction of nuclear proteins
The Hepa1c1c7 cells were washed twice in Dulbecco's phosphate-buffered saline (DPBS), harvested by trypsinization, and suspended in 1 mL of DPBS, followed by centrifugation (800 g for 3 min at 4°C). The cell pellet was carefully resuspended in 200 μL of cold lysis buffer A, consisting of 10 mM HEPES (pH 7.9), 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM dithiothreitol, and complete protease inhibitor cocktail (Sigma-Aldrich). The pellet was then incubated on ice for 15 min to allow cells to swell. After this time, 15 μL of 10% Nonidet P-40 was added, and the tube was vortex mixed for 10 sec. The homogenate was then centrifuged at 800 g for 3 min at 4°C. The resulting nuclear pellet was resuspended in 30 μL of cold buffer B, consisting of 20 mM HEPES (pH 7.9), 0.4 M NaCl, 1 mM EDTA, 1 mM EGTA, 1 μM dithiothreitol, and protease inhibitors. The pellet was then incubated on ice for 15 min and vortex mixed for 10–15 sec every 2 min. The nuclear extract was finally centrifuged at 15,000 g for 5 min at 4°C. The supernatant containing the nuclear proteins was stored at−80°C.
Western blot analysis
Protein expression of Nrf2, NQO1, HO1, and GST-P was measured by western blotting. In cell culture experiments, the medium was removed, and cells were rinsed with DPBS twice. After the addition of 0.5 mL of cold RIPA buffer and protease inhibitors (Sigma-Aldrich), cells were scraped, followed by vortexing at 0°C for 20 min. The cell lysates were then subjected to centrifugation at 10,000 g for 30 min at 4°C. The protein (50 μg) from the supernatant was resolved on SDS-PAGE and transferred onto a polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA). The membrane was blocked with 5% skim milk in the TBS-T buffer (20 mmol/L Tris–HCl, 150 mmol/L NaCl, and 0.1% Tween-20, pH 7.4) for incubation for 1 h at room temperature and then incubated overnight at 4°C. It was probed using primary antibodies that recognized Nrf2, NQO1, HO1, and GST-P (Santa Cruz Biotechnology, Santa Cruz, CA, USA); these were diluted in TBS-T containing 1% skim milk. After washing in TBS-T, the membrane was then incubated for 1 h with the horseradish peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology) at room temperature. Immunoreactive proteins were detected using enhanced chemiluminescence reagents (Amersham Biosciences; GE Healthcare, Little Chalfont, United Kingdom) and measured by densitometry using a Las-4000 image reader and Multi Gauge 3.1 software (Fuji Photo Film, Tokyo, Japan).
Test animals
Male Sprague-Dawley rats (200±20 g) were purchased from the Orient Co. (Sungnam, Korea). They were housed in a light–dark cycle at 23°C±3°C in 50%±10% relative humidity. The animals were allowed to acclimate to the housing conditions for 7 days before experimentation. The animal study was performed according to the international rules governing animal experiments and the internationally accepted ethical principles for laboratory animal use and care. The experiments were performed at the Experimental Animal Center, Hallym University, Chuncheon, Korea.
Experiment diet
Rats were divided into six groups consisting of 10 animals in each group. Rats in the control group received only distilled water and standard diet. The ethanol group received 5 g of 20% ethanol/kg body weight. Test-group animals were administered a dose of 50 mg/kg body weight (CH 50) and 100 mg/kg body weight (CH 100), respectively. The SFN group received SFN 10 mg/kg body weight. Liver damage was induced by administration of 5 g of 20% ethanol/kg body weight for 16 days twice daily. After administration of ethanol intake for 17 days, diet containing the test substance was intake for 10 days, and continued CH intake of ethanol was administered orally. On day 27, animals were anesthetized using ethyl-ether; blood was collected, allowed to clot, and serum was separated for assessment of enzyme activity. The rats were then sacrificed by bleedings; the livers were carefully dissected and cleaned of excess tissue.
Blood and liver biochemical marker assay
The activity of the following serum enzymes was measured: plasma glutamic oxaloacetic transaminase (GOT), glutamic pyruvic transaminase (GPT), γ-glutamyltransferase (γ-GT), and lactate dehydrogenase (LDH). Plasma GOT, GPT, γ-GT, and LDH activities were determined by enzymatic colorimetric methods that were performed using appropriate kits (Wako Pure Chemical Co., Osaka, Japan).
Histopathological studies
Liver tissues were fixed in 10% formalin for at least 24 h, fixed in paraffin, and cut into 5-μm-thick sections using a rotary microtome. The sections were stained with hematoxylin–eosin dye and observed under a microscope (IX51; Olympus, Tokyo, Japan) to observe histopathological changes in the liver.
Statistical analysis
Biochemical estimates are reported as the mean±SEM of six animals from each group. The data were subjected to one-way ANOVA using SAS version 9.1 (SAS Institute, Cary, NC, USA) followed by Duncan's multiple range test. P value of<.05 was considered statistically significant.
Results and Discussion
QR activity of CH and its individual fractions
We first investigated the QR activity of extracts and fractions isolated from CH on the QR activity using Hepa1c1c7 cells, a model cell line for measuring the QR activity. We isolated the active final fraction from CH F4-2. The final fraction was collected by HPLC and its identity confirmed by comparing spectral characteristics. The results for both final fraction and chlorophyll a were a maximum absorption spectrum of 660 nm. For comparison, the characteristics of the purified final fraction and chlorophyll a are shown in Figure 1. As shown in Figure 2, the purified final fraction (CH F4-2, 3.13–200 μg/mL) induced the QR enzyme by a concentration-dependent manner. Moreover, CH F4-2 did not show cell cytotoxicity in the Hepa1c1c7 cell. Chlorophyll and its various derivatives have a long established history of use in traditional medicine and for therapeutic purposes. 17 The association of diets rich in phytochemicals with the prevention of liver injury has intensified interest in chlorophyll as a class of plant pigments with potential hepatoprotective effects. Considering the traditional use of chlorophyll derivatives in medicine and their low toxicity, these pigments serve as attractive liver disease preventatives and potentially as therapeutic agents. 18 –20 According to Usuki et al., 21 a reaction medium that contains unsaturated fatty acids promotes the decomposition of chlorophylls, thereby reducing its antioxidant capacity. Natural chlorophyll derivatives have also been investigated with regard to antimutagenic and antigenotoxic activity. 22 Natural chlorophylls have demonstrated antigenotoxic and antimutagenic activity in mammalian cell-based assays. 23 We suggest that the purified final fraction from CH was identical to chlorophyll a, so far as inducing QR activity, and has hepatoprotective activity.

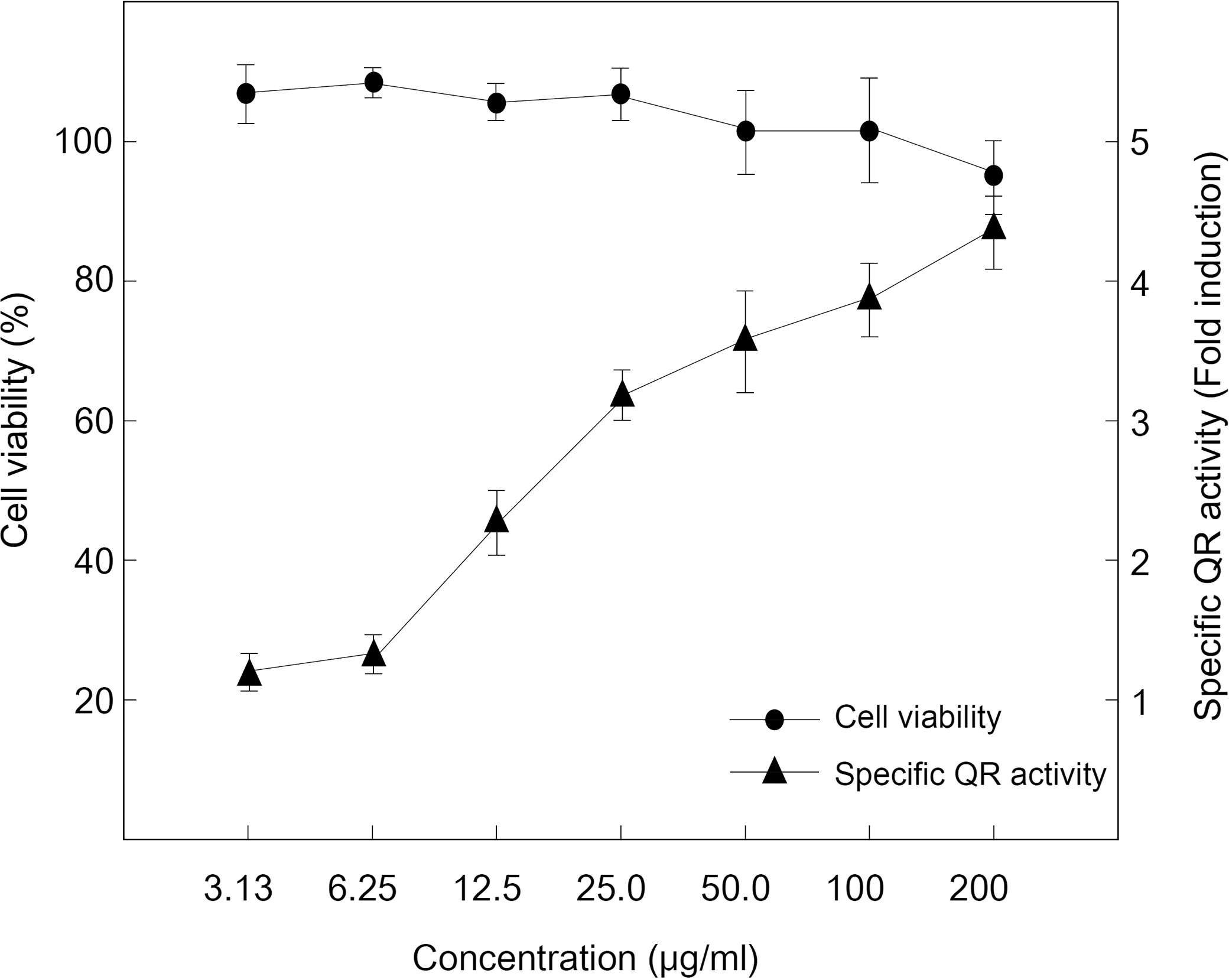
Effects of purified final fraction (CH F4-2) on cytotoxicity and quinone reductase (QR) activity (fold induction) in Hepa1c1c7 cells.
mRNA induction and protein levels
Treatment of purified fraction (CH F4-2) (25–200 μg/mL) for 12 h increased HO1 and NQO1 mRNA expression in a concentration-dependent manner, whereas the GST-P expression at 12 h was lower compared with the HO1 or NQO1 (Fig. 3A, B). In contrast, CH F4-2 did not significantly affect HO1 or NQO1 mRNA expression after 24 h of incubation. The bifunctional inducers upregulate phase II enzymes, including QR and a few phase I enzymes such as CYP1A1. 24 These results suggest that the isolated and purified CH F4-2 from CH induces QR activation through upregulation of the NQO1 and HO1 gene mRNA in Hepa1c1c7 cells. To further confirm that CH affects protein expression of NQO1, HO1, GST-P, and Nrf2 in Hepa1c1c7 cells, we found that the expression of NQO1 and HO1 protein also increased in a dose-dependent manner comparable to that of 5 μM of SFN. In particular, NQO1 and HO1 expression levels were dramatically increased for 24 h after treatment with CH 200 μg/mL. Although the level of GST-P at 48 h was lower than at 24 h (Fig. 3C, D), the results indicate that CH increases NQO1, NO1, and GST-P at both the mRNA and protein levels. It has been known that the Nrf2, a key regulator for the induction of phase II detoxification enzymes, acts through Nrf2-mediated ARE activation. 25 To determine if CH F4-2 affected the translocation of Nrf2 to the nucleus, we performed western blot analysis of the cytoplasmic and nuclear fractions of Hepa1c1c7 treated with CH F4-2. A significant increase in Nrf2 protein levels in the nuclear fractions was observed after treatment with CH F4-2 for 12 h at 200 and 100 μg/mL. We also found that Nrf2 protein levels were very low in the cytoplasmic fractions (Fig. 3E, F). These results suggest that the induction of NQO1 by CH F4-2 in Hepa1c1c7 cells was mediated by the Nrf2-ARE pathway through the translocation of Nrf2 from the cytoplasm to the nucleus. Previous reports have revealed that the activation of Nrf2-ARE, signaled by antioxidative phytochemicals that induce cytoprotective enzymes, may at least, in part, be due to their pro-oxidant activity. 25 Oxidative stress is also closely related to inflammatory responses, such as chronic obstructive pulmonary diseases. HO1 plays an important role in attenuating the production of ROS through degrading heme to equimolar amounts of carbon monoxide and biliverdin/bilirubin and the release of free iron. Therefore, pharmacologic modulation of the HO1 system may be an effective and cooperative strategy to intervene in the protection against alcoholic liver disease. We found that the mRNA and protein levels of HO1 in chlorella extract-treated hepatoma cells were significantly increased. As HO1 can be activated by Nrf-2, a major transcription factor regulating ARE-driven phase II gene expression, 26 the cytosolic and nuclear protein levels of Nrf-2 were then detected by western blot. The activation of the HO1/Nrf-2 system could be attributed to many upstream kinases, including protein kinase C, protein kinase A, phosphatidylinositol 3-kinase (PI3K), and mitogen-activated protein kinases (MAPKs). Among these upstream signaling kinases, MAPKs have been considered to play major roles in controlling the upregulation of HO1. 27 These data suggested that the purified final fraction of CH, chlorophyll promoted the nuclear translocation of Nrf-2 and then the induction of HO1.
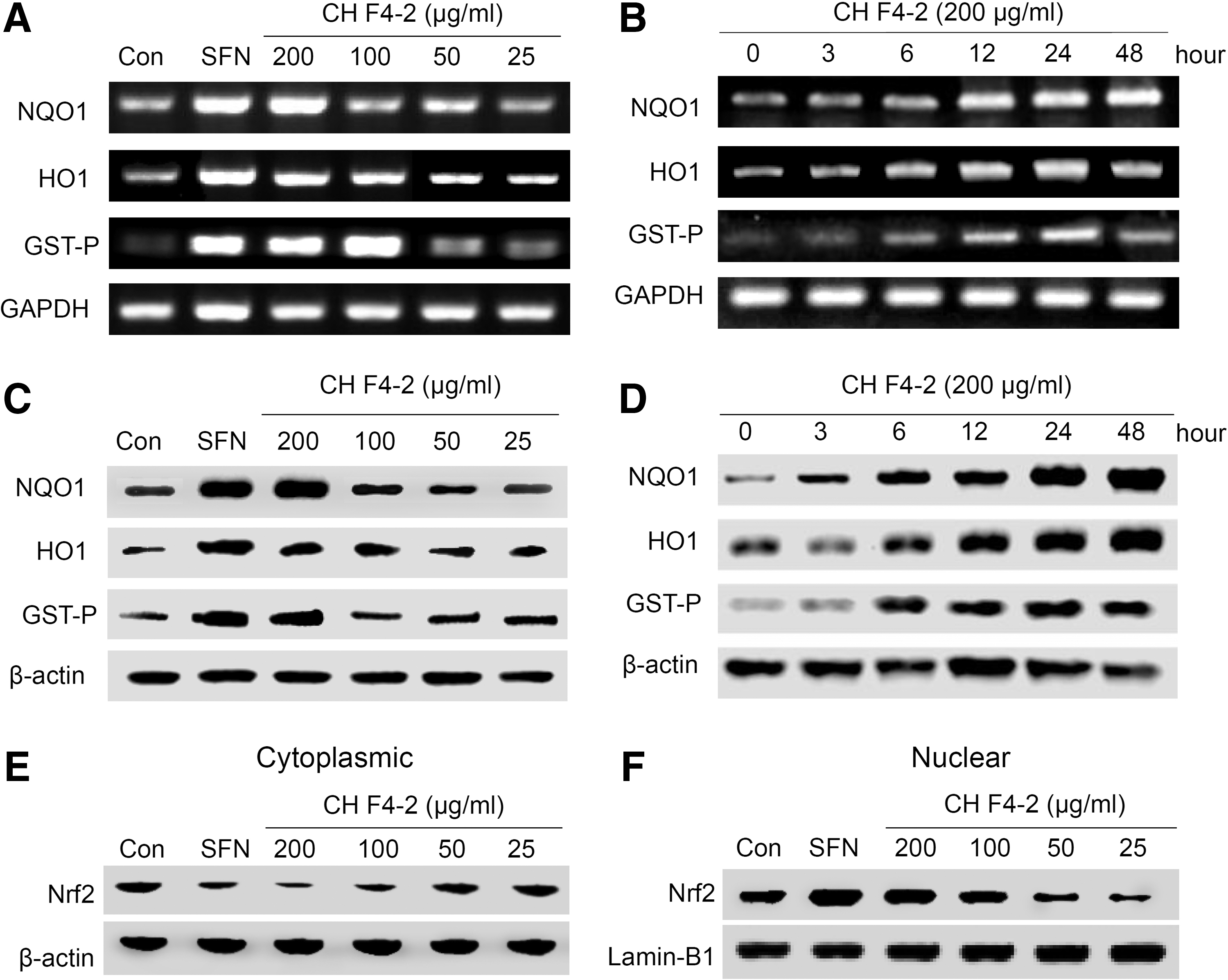
The purified final fraction (CH F4-2) treatment cause
Plasma GOT, GPT, γ-GT, and LDH activities
Plasma GOT and GPT are primary indicators of liver injury that may be detected efficiently when there are high levels of GOT and GPT activities, with enzymes abundant in the liver and present in the plasma. While the ratios may vary under different circumstances, liver damage due to chronic ethanol has been indicated by the ratio of GOT to GPT. 26 In our study, liver injury was sustained by individual rats suffering from chronic ethanol treatment, as indicated by the high concentrations of hepatic enzymes in the plasma. However, it was also apparent that the CH supplementation effectively suppressed injuries by decreasing the plasma level of GOT and GPT (Fig. 4A, B). GOT and GPT were present at high concentrations in the ethanol-intake group, while they leaked in the CH-intake group. Levels of plasma GOT and GPT on the CH supplementation group were effective in decreasing up to 22.3% and 22.8% than the ethanol-intake group, respectively. In addition, GOT and GPT levels decreased in a dose-dependent manner by the increase in CH concentration. LDH levels in the CH-intake group were dramatically lower than in the ethanol-intake group (Fig. 4C, D). γ-GT and LDH are the most sensitive of the markers of liver damage and disease on alcohol induced. 28 LDH is an enzyme found inside almost all cells, but normally it exists in only trace amounts in the blood. LDH in the bloodstream indicates that cells have been damaged or destroyed. Therefore, LDH is a general marker of cell injury, but is not useful for identifying particular damaged cells. 28 Previous studies have demonstrated that chlorophyll is effective in countering alcohol intoxication by suppressing the chlorophyll through delayed stomach emptying, but not by enhancing the clearance of alcohol. 20 Similarly, our data showed that the GOT, GPT, γ-GT, and LDH levels in rats given CH were significantly lower than that in the ethanol group. It has been suggested that oxidative stress might contribute to functions associated with ethanol administration in vivo. CH contains a high level of chlorophyll, a potent antioxidant that might directly decrease damage induced by ethanol administration.
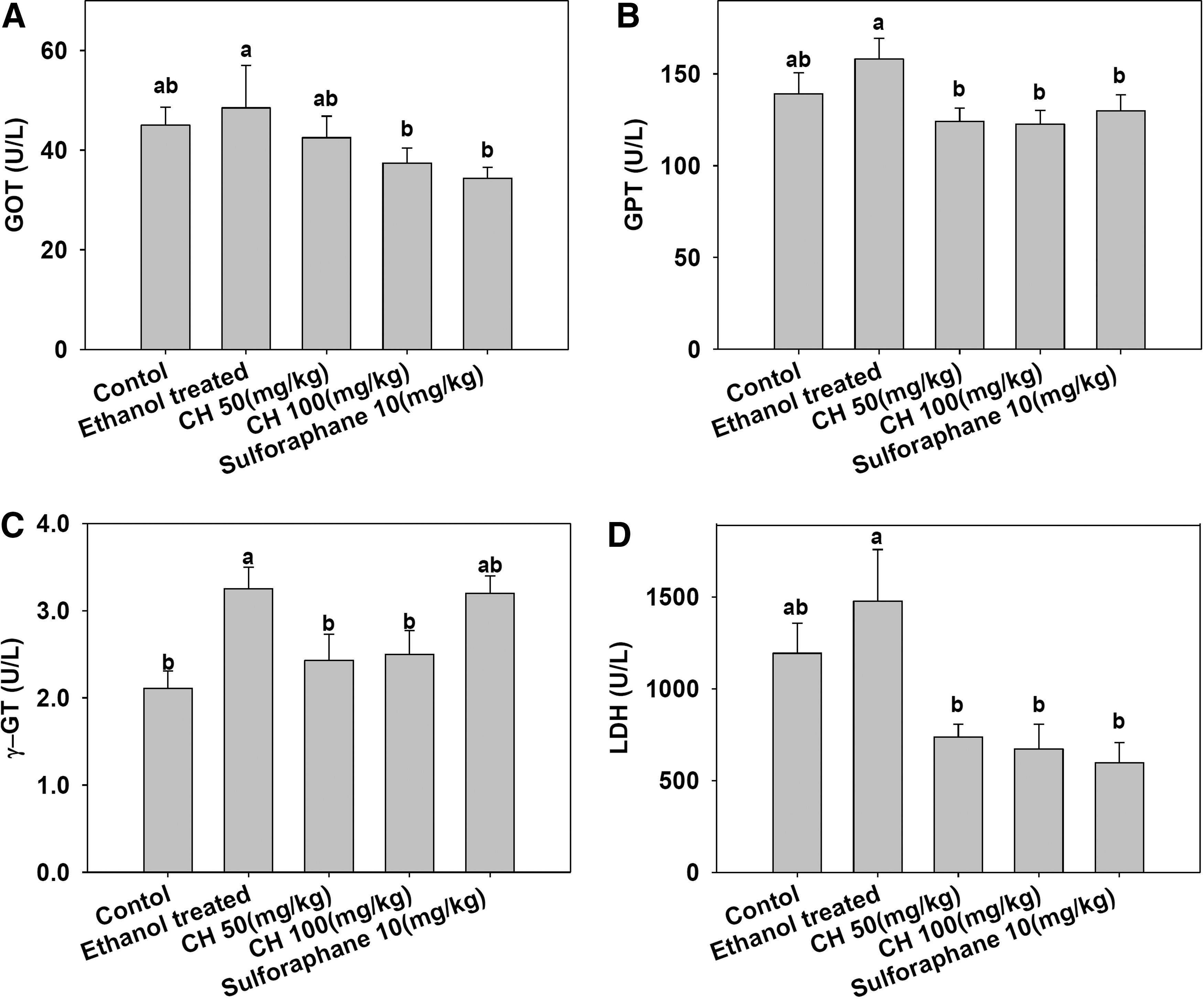
Effects of CH on plasma
Long-term alcohol consumption in immoderate quantities can lead to chronic alcoholic liver disease, which is one of the most common causes of death worldwide. 29 Various studies reported that excessive ROS are produced during ethanol metabolism through alcohol dehydrogenase, catalase, and especially by CYP2E1. 30,31 Our study has shown that CH possesses potential hepatoprotective effects in vivo and in vitro against ethanol. Moreover, CH may reserve potential to be further developed for its pharmacological function as a prophylactic and therapeutic agent for chronic liver disease caused by alcohol. The CH significantly increased HO1 expression and activity in the hepatoma mouse cell, Hepa1c1c7, although it was slightly less effective than SFN. It is notable that HO1 inducers from CH may possess potential toxicity, while a naturally occurring phytochemical may act as nonstressful and noncytotoxic HO1 inducers. 32 The findings in this study suggested a direct contribution of phytochemical-mediated HO1 expression in protecting hepatocytes against ethanol-induced oxidative stress.
The activation of Nrf2 occurs when cells exposed to oxidative stress, allowing Nrf2 to translocate and accumulate in the nucleus, further drive the transcriptional activation of genes whose expression is essential to control cellular redox homeostasis. 33 To identify the signal cascade used by CH to control Nrf2 activation and upregulate HO1 in the hepatoma mouse cell, Hepa1c1c7, MAPKs, and PI3K pathways were investigated. It has been accepted that MAPKs play a central role in the signaling pathway involved in cell growth and stress response. 34 The three major subfamilies of MAPKs, extracellular signal-regulated kinases (ERK), jun amino-terminal kinases (JNK) and p38 appear to be related to the upregulation of HO1 expression by diverse stimuli. 35 PI3K was also reported to be involved in the pathway regulating Nrf2 activation as well as HO1 expression in response to the phytochemical antioxidant in PC12 cells. 36 Therefore, HO1 induction was involved in the hepatoprotective effect of CH. The ERK/Nrf2 signaling transduction pathway probably participated in HO1 induction by CH in the hepatoma mouse cell, Hepa1c1c7. These findings warrant future in vivo studies of the supplementation of CH food/medicine for the prevention and/or treatment of ethanol-induced hepatotoxicity.
Histopathological
The effect of CH on the liver histopathology of ethanol-treated rats is presented in Figure 5: a liver section of a normal rat shows distinct hepatic cells with a well-preserved cytoplasm and a prominent nucleus and nucleolus. Ethanol-induced liver damage can be observed directly: the figure 5B section shows massive fatty change, necrosis, lymphocyte infiltration, loss of cellular boundaries, and the joined together nucleus. Figure 5C and D shows a section from among the group treated with high-dose CH 50 and 100 mg/kg. Sections of these two groups were nearly the same as those of the normal group, with no obvious necrosis observed. In these results, it can be supposed that the ethanolic extracts of the CH extract seem to have hepatoprotective effects in rats. Moreover, we have investigated how CH and its individual fractions affect the induction of phase II detoxification enzymes. We found that all extracts and fractions affected induction of QR activation; we also found increased mRNA levels and the protein level of QR through the Nrf2-ARE pathway. CH and CH F4-2 (chlorophyll a) could be developed as hepatoprotective agents for treatment in humans.

Liver section from ethanol-treated rats showing hepatic cell.
Footnotes
Acknowledgment
This research was supported by a grant from the Marine Bioprocess Research Center of the Marine Biotechnology Program funded by the Ministry of Land, Transport, and Maritime, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.