
Abstract
Vigna species are important food resources and are traditionally used for the treatment of various diseases. In this study, we examined the inhibitory effects of Vigna nakashimae (VN) extract on high-fat diet (HFD)-induced hepatic steatosis and elucidated the molecular mechanisms. C57BL/6J mice were fed an HFD with or without VN extract for 16 weeks. VN extract decreased HFD-induced body weight, liver weight, hepatic lipid accumulation, and plasma alanine aminotransferase, and suppressed oxidative stress and inflammation associated with hepatitis. VN extract decreased plasma lipid levels and the expression of lipogenic genes in the livers of HFD-fed mice. VN extract significantly increased phosphorylation of adenosine monophosphate (AMP)-activated protein kinase (AMPK) and acetyl-CoA carboxylase, and expression of fatty acid oxidation genes in the liver of VN-treated HFD mice and HepG2 cells. Further, VN extract inhibited insulin or glucose-stimulated lipogenesis in HepG2 cells. In conclusion, VN extract prevents HFD-induced hepatic steatosis and lipotoxicity through AMPK activation.
Introduction
N
Adenosine monophosphate (AMP)-activated protein kinase (AMPK), a metabolic sensor that acts as a cellular fuel gauge in eukaryotes, is involved in cellular energy homeostasis. 3 AMPK stimulates the pathways that increase energy production such as fatty acid oxidation, and it switches off the pathways that consume energy such as lipogenesis. In addition to controlling energy homeostasis, AMPK enhances insulin sensitivity by increasing glucose uptake and lipid oxidation in skeletal muscle and inhibiting glucose and lipid synthesis in the liver. 3,4 AMPK is, therefore, a key molecule for controlling metabolic diseases such as type 2 diabetes, obesity, and hepatic steatosis. Discovery and development of a natural AMPK activator could provide a novel strategy for overcoming these metabolic diseases.
The plant genus Vigna is an important member of the family Fabaceae. This genus contains several species that are of considerable economic importance and dietary staples for many millions of people. 5 Among the Vigna species, the cultigens of mungbean [Vigna radiata (L.); Wilczek], black gram [Vigna mungo (L.); Hepper], and azuki bean [Vigna angularis (Willd.); Ohwi & Ohashi] are the most important economically. In addition to being important food resources, Vigna species are traditionally used for the treatment of various diseases, including metabolic disorders, because they possess many bioactive phytochemicals, including phenolic compounds. 6 The extracts of V. angularis improved blood glucose and cholesterol in high-fat diet (HFD)-induced mice. 7 In addition, the hypoglycemic effects of extracts of V. angularis were also confirmed in type 2 diabetes mellitus model KK-A(y) mice and a streptozotocin-induced type 1 diabetes model. 8,9 Previously, we reported that Vigna nakashimae (VN), another species of Vigna that is widely cultivated in Korea, has antidiabetic effects in db/db mice via inhibition of α-glucosidase activity 10 and antiobesity effects in HFD obese mice. 11 Recently, the search for appropriate hepatic steatosis agents has focused on plants used in traditional medicine. The Vigna species of legumes has been reported to have polyphenols that suppress TG accumulation in hepatocytes. 12 –14
Therefore, in this study, we evaluated whether VN has the potential to prevent hepatic steatosis using an HFD obese animal model and the HepG2 hepatocyte cell line, and investigated the molecular mechanisms behind its effects.
Materials and Methods
Reagents
Antibodies against AMPK, phospho-AMPK (Thr172), S6 kinase (S6K), and phospho-S6K were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA), and antibodies against adenosine acetyl-CoA carboxylase (ACC) and phospho-ACC (Ser79) were purchased from Cell Signaling Technology (Beverly, MA, USA). Insulin, glucose, and T0901317 were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Preparation of VN extract
VN (IT178464) germplasm was provided by the Genetic Resources Division of the National Institute of Agricultural Biotechnology, Rural Development Administration in South Korea, and was extracted as previously described. 11 A voucher specimen was deposited at the National Institute of Agricultural Biotechnology. The dried seeds of VN (3.0 kg) were pulverized using a grinder and extracted with 10 L of 80% methanol for 3 days at room temperature. After filtration, methanol extracts were concentrated in vacuo at 35°C to give a brownish residue. The crude extract was freeze dried and ground into a powder (260 g). The powder was kept at −70°C until it was analyzed.
Quantification of total phenolic, total flavonoid, and total anthocyanin contents
The VN extracts were dissolved in 70% ethanol, and used for determining the phenolic indexes. The total phenolic content was measured using the Folin–Ciocalteu method. 15 Total flavonoid content was measured using the colorimetric method as previously described by reading the absorbance at 510 nm. 16 The total anthocyanin content was determined according using a spectrophotometric method by measuring the absorbance at 520 nm. 17
High-performance liquid chromatography analysis of polyphenolic compounds in VN extract
To determine the contents of flavonoid compounds in VN extract, high-performance liquid chromatography (HPLC) analysis was performed on a Waters 1525 liquid chromatography system (Milford, MA, USA). The flavonoid compounds [(+) catechin, (−) epigallocatechin gallate (EGCG), rutin, and quercetin] were monitored at 280 or 368 nm using TSKgel ODS-100V (4.6×250 mm). The concentrations were calculated from the peak heights of the internal standard and each compound in the samples and in reference solutions.
Animal experiments
C57BL/6 mice (male, 4 weeks old) were purchased from Jung-Ang Lab. Animal, Inc. (Seoul, South Korea). The animals were housed in a controlled environment: temperature (21–23°C) and humidity (40–60%) control with a 12 h light/12 h dark cycle, and were given free access to food and water. After a 1-week adaptation period, the mice were randomly divided into four groups (n=7) with equal mean body weights: normal chow diet control (distilled water-treated) group, HFD (distilled water-treated) group, HFD [VN 100 mg/(kg·day−1) of body weight] group, and HFD [VN 300 mg/(kg·day−1) of body weight] group. The experimental diets were based on the HFD containing 30% lard (45% kcal% fat) and the AIN93M-based control diet (Research Diets, New Brunswick, NJ, USA). VN extracts were dissolved in distilled water and administered orally every day for 16 weeks. Body weight and blood glucose level were checked every 5 days. All animal experiments were approved by the Pusan National University Animal Experiment Ethics Committee and were conducted in accordance with the institutional guidelines for the care and use of laboratory animals.
Histological analysis
Fresh liver tissue was immediately fixed with 10% neutral-buffered formalin and embedded in paraffin wax. The liver slides were stained with hematoxylin and eosin, and the histopathology was photographed under a microscope.
TG measurement
The TG contents in liver homogenates and HepG2 extracts were determined using Sigma Diagnostic Triglyceride Reagents. Liver tissue was homogenized in 4 mL of chloroform-methanol (2:1). About 0.8 mL of 50 mM sodium chloride was added to homogenate, and the solution was incubated at 4°C for 12 h. The sample was centrifuged, and the organic layer was removed and dried using a speed vac. The resulting pellet was dissolved in phosphate buffer saline containing 1% Triton X-100 and then assayed for TG.
Assay of thiobarbituric acid reactive substances formation
The extent of lipid peroxidation was estimated in liver homogenates by the levels of malondialdehyde (MDA) measured using the thiobarbituric acid reactive substances (TBARS) assay at 535 nm. 18
Determination of glutathione peroxidase and catalase activities
The activities of the antioxidant enzymes, glutathione peroxidase (GPx) and catalase, were assayed in liver homogenates using commercially available kits, GPx (Cayman, Ann Arbor, MI, USA) and catalase assay kits (Sigma-Aldrich), respectively. GPx activity was determined by following the enzymatic NADPH oxidation at 340 nm, and catalase activity was spectrophotometrically measured by following the rate of hydrogen peroxide consumption at 240 nm.
Determination of reduced glutathione
The glutathione levels were determined in the 5,5′-dithiobis-(2-nitrobenzoic acid)-glutathione disulfide reductase recycling assay with some modifications using a glutathione (GSH) determination kit (Oxis International, Portland, OR, USA).
Analysis of blood biochemical parameters
Food was withheld for 12 h, after which the mice were sacrificed. The plasma was harvested from the mice and analyzed. Plasma alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were determined using Infinity ALT and AST Reagents, respectively (Thermo Scientific, Middletown, VA, USA). The concentrations of TG, non-esterified fatty acid (NEFA), total cholesterol (TC), and high-density lipoprotein (HDL)-cholesterol were determined using a commercial analysis kit (Asan Diagnostics, Seoul, South Korea).
Cell culture
HepG2 cells, which were purchased from American Type Culture Collection (ATCC, CL-173TM; Manassas, VA, USA), were maintained in Dulbecco's modified Eagle's medium (HyClone, Logan, UT, USA) containing 10% fetal calf serum (HyClone), 50 units/mL penicillin, and 50 mg/mL streptomycin at 37°C in a humidified atmosphere with 5% CO2. The cells were treated with vehicle or liver X receptor (LXR) agonist (10 mM T0901317) in the presence or absence of VN extract for 24 h.
Reverse transcription-polymerase chain reaction
Total RNAs were isolated from the livers of the mice and HepG2 cells with Trizol™ (Invitrogen, Carlsbad, CA, USA). The cDNAs were generated from 1 μg of total RNAs using Moloney Murine Leukemia Virus reverse transcriptase (Promega, Madison, WI, USA) at 42°C for an hour and then used for polymerase chain reaction (PCR) analysis. The primers used in this study are listed (Supplementary Table S1; Supplementary Data are available online at
Western blot
Equal amounts of protein (40 μg/lane) from liver homogenates and HepG2 cell lysates were resolved by 8% SDS130 polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride 131 membranes (Millipore, Massachusetts, MA, USA). The levels of pAMPK, AMPK, pACC, ACC, pS6K, S6K, and actin were detected using an enhanced chemiluminescence western blot detection kit (Amersham, 137 Uppsala, Sweden).
Statistical analysis
All data are presented as the mean±standard deviation. The data were evaluated by one-way analysis of variance, and the differences between means were determined using Tukey–Kramer post-hoc test. Values were considered statistically significant at P<.05.
Results
Analysis of polyphenolic contents in VN extract
Since Vigna species have been reported to possess polyphenols, which are reported to have efficacy for treating metabolic disorders, 6 we determined the contents of polyphenols in VN extract. The total phenolic, flavonoid, and anthocyanin contents were 38.72 mg/g gallic acid, 2.53 mg/g quercetin, and 5.25 mg/g cyanidin, respectively (Table 1). Among the phenolic compounds, HPLC analysis of VN extract showed that it contained 3.72 mg/g (+) catechin, 1.78 mg/g (−) EGCG, 0.11 mg/g rutin, and 0.038 mg/g quercetin (Table 1), indicating that VN extract contains high levels of catechins, including (+) catechin and EGCG.
Data are expressed as mean±standard deviation of triplicate samples.
Total phenolic content is expressed as mg of gallic acid equivalents/g of VN extract.
Total flavonoid content is expressed as mg of quercetin equivalents/g of VN extract.
Total anthocyanin content is expressed as mg of cyanidin-3-glucoside equivalents/g of VN extract.
VN, Vigna nakashimae.
VN extract inhibited HFD-induced hepatic steatosis and protected against HFD-induced liver injury
Since EGCG, rutin, and quercetin have been demonstrated to suppress dyslipidemia and oxidative stress in food-induced obese animals, 19 –21 we evaluated the effects of VN extract on HFD-induced hepatic steatosis. C57BL/6J mice were fed an HFD and orally administered VN extracts [100 mg/(kg·day−1), 300 mg/(kg·day−1)] daily for 16 weeks. Consistent with previous results, 11 the weight gain in the VN extract-treated HFD mice was significantly less than that in HFD control mice (Fig. 1A). Liver weight was also significantly lower in VN (300 mg/kg) extract-treated HFD mice compared with HFD control mice (Fig. 1B). No significant difference in food intake was observed between the control and VN extract-treated mice (data not shown). To examine the effects of VN extract on hepatic steatosis, lipid accumulation was assessed in the livers of HFD mice by Oil Red O staining and measuring TG content. As shown in Figure 1C, Oil Red O staining of liver sections revealed that 300 mg/kg VN extract reduced HFD-induced lipid accumulation in the mice. TG content was also reduced in 300 mg/kg VN extract-treated HFD mice compared with that in HFD control mice (Fig. 1D). Consistent with this, 300 mg/kg VN extract significantly reduced HFD-induced cytoplasmic lipid droplets in the liver, as revealed by hematoxylin and eosin stain (H&E) staining (Fig. 1E). These results suggest that VN extract ameliorates fatty liver in HFD-induced obese mice. Fat accumulation in the liver is known to contribute to hepatitis. 2 To determine the effect of VN extract on HFD-induced hepatitis, we measured the levels of AST and ALT, indicators of hepatitis, in the plasma. As shown in Figure 1F, HFD increased the activities of AST and ALT in plasma, which was prevented by administration of 300 mg/kg VN extract. All these results suggest that VN extract attenuates hepatic steatosis and protects against liver injury in the progression of steatosis to hepatitis.

VN extract inhibited hepatic steatosis in HFD obese mice. Male C57BL/6 mice were fed either a normal or an HFD for 16 weeks.
VN extract inhibited HFD-induced oxidative stress and inflammation
To determine whether the diminished steatosis in the livers of mice treated with VN extract was associated with attenuated oxidative stress, we examined the oxidative stress-mediated MDA formation in the livers of HFD obese mice. HFD increased formation of TBARS, which was inhibited by 300 mg/kg VN extract treatment (Fig. 2A). Oxidative injury may result from a decrease in hepatic antioxidant enzymes such as GPx and catalase and in GSH content. HFD feeding decreased the activities of GPx (Fig. 2B) and catalase (Fig. 2C), whereas treatment with 300 mg/kg VN extract significantly restored them. In addition, VN extract prevented the HFD-mediated decrease in GSH content (Fig. 2D). We further defined whether VN extract attenuates inflammation associated with steatohepatitis. As shown in Figure 2E, treatment with 300 mg/kg VN extract inhibited the HFD-mediated increase in the expression of inflammatory genes, including tumor necrosis factor (TNF)–α, interleukin (IL)–6, and monocyte chemotactic protein (MCP)–1 (Fig. 2E). VN extract (300 mg/kg) also reversed the expression of cyclooxygenase-2 and inducible nitric oxide synthase induced in HFD obese mice (Fig. 2F). These results suggest that VN extract prevents oxidative stress and inflammation associated with hepatitis.
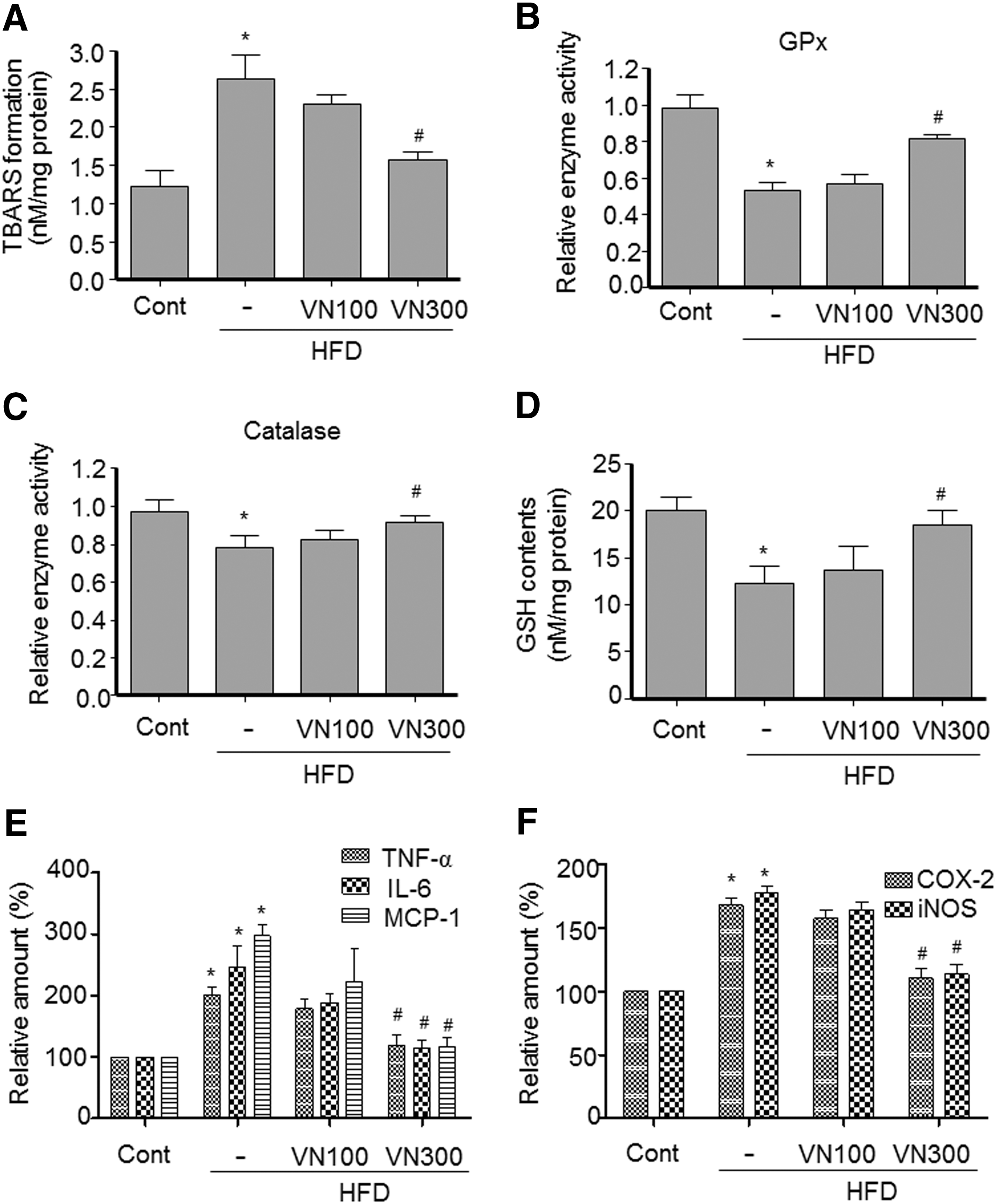
VN extract inhibited HFD-induced oxidative stress and inflammation. Male C57BL/6 mice were fed either a normal or an HFD for 16 weeks.
VN extract inhibited hepatic lipogenesis and ameliorated plasma lipids in HFD obese mice
To investigate whether VN extract modulates lipogenesis in the livers of HFD obese mice, we examined the effect of VN extract on the expression of LXR and sterol regulatory element binding protein-1c (SREBP-1c), master lipogenic transcription factors, 22 and their target genes. The expression of LXR and SREBP-1c was increased in the liver of HFD obese mice, whereas treatment with 300 mg/kg VN extract inhibited HFD-induced LXR and SREBP-1c gene induction (Fig. 3A). Consistent with this, expression of their target lipogenic genes, including L-pyruvate kinase, fatty acid synthase (FAS), stearoyl-CoA desaturase (SCD)–1, and Spot 14 (S14), was also increased in the liver of HFD obese mice but decreased by treatment with 300 mg/kg VN extract (Fig. 3A), indicating that VN extract represses the expression of hepatic lipogenic genes and inhibits fat accumulation in the liver.
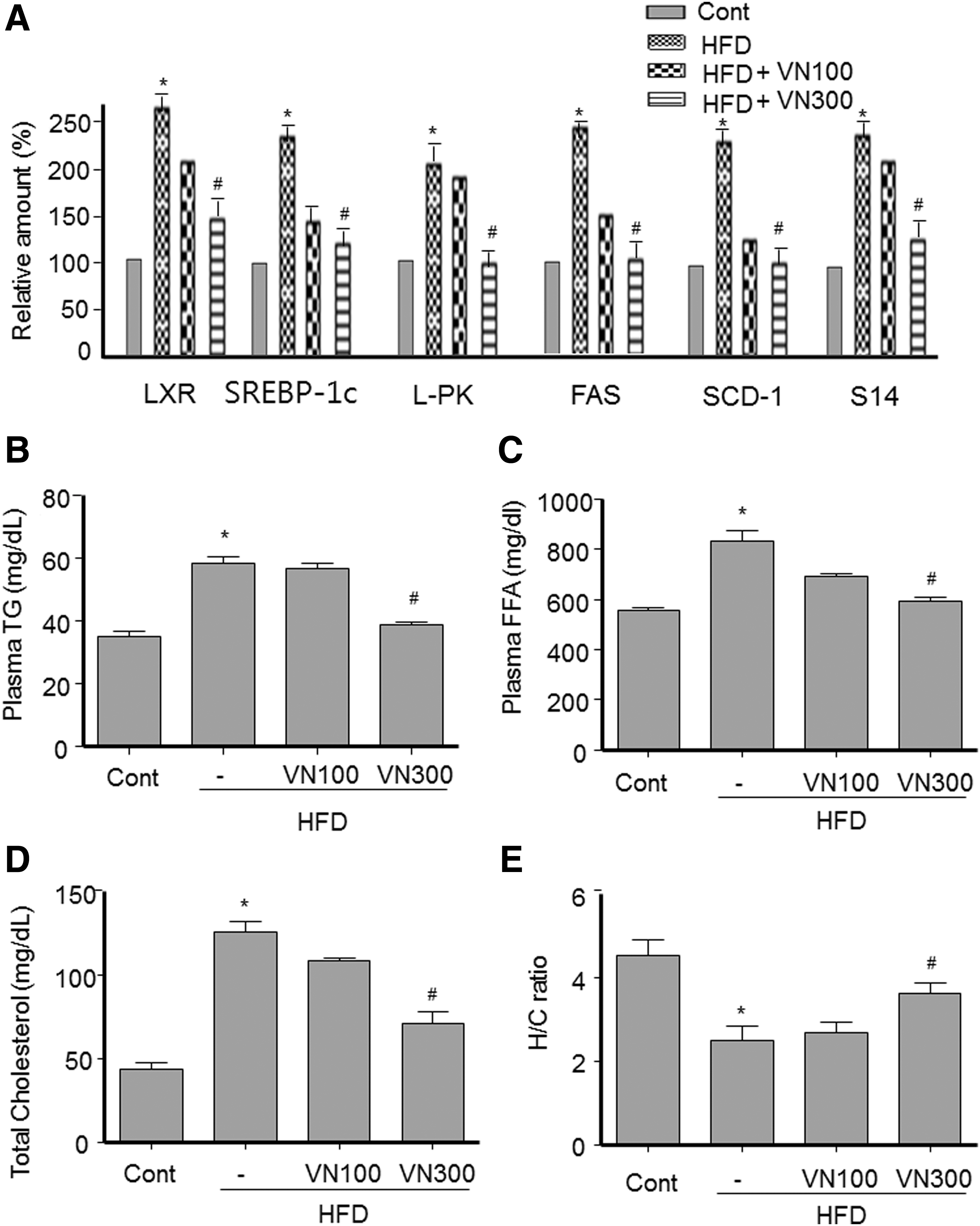
VN extract repressed expression of lipogenic genes and decreased blood lipid levels in HFD obese mice.
To investigate whether alleviation of hepatic steatosis by VN extract was correlated with changes in plasma levels of lipids, blood samples were obtained from each group of mice. As shown in Figure 3, administration of 300 mg/kg VN extract reduced the HFD-induced levels of TG (Fig. 3B), NEFA (Fig. 3C), and TC (Fig. 3D), which was consistent with decreased TG accumulation in the liver tissues. In contrast, 300 mg/kg VN extract increased the ratio of HDL-cholesterol/TC (Fig. 3E). These data indicate that VN extract ameliorates blood lipid levels.
VN extract activated AMPK in liver of HFD obese mice and HepG2 cells
Since AMPK activation stimulates fatty acid oxidation and inhibits LXR-mediated SREBP-1c induction, 23,24 we assessed the activation of AMPK and its downstream signaling in the livers of HFD obese mice to elucidate the molecular mechanism involved in antihepatic steatosis of VN extract. As shown in Figure 4A, the level of phosphorylated AMPK was significantly decreased in the livers of HFD obese mice. However, treatment with 300 mg/kg VN extract restored AMPK phosphorylation in the liver. AMPK activation inactivates ACC, a downstream target of AMPK, by phosphorylation at serine 79 and stimulates the expression of fatty acid oxidation genes. 3 Therefore, the level of ACC phosphorylation and the expression of fatty acid oxidation genes were investigated in the liver of the mice. Consistent with activation of AMPK, the level of phosphorylated ACC (Fig. 4A) and the expression of carnitine-palmitoyltransferase-1 (CPT-1) and acyl-CoA oxidase (ACO) (Fig. 4B) were increased in the liver of 300 mg/kg VN extract-treated HFD mice compared with HFD control mice. Further, we also investigated the effect of VN extract on S6K1 activity in HFD obese mice, because AMPK inhibits the activation of LXR by S6K1. 24 As shown in Figure 4A, HFD feeding increased phosphorylation of S6, a substrate of S6K1, whereas treatment with 300 mg/kg VN extract blocked the HFD-mediated increase in phosphorylation of S6. Collectively, these results suggest that VN extract activates AMPK, which stimulates fatty acid oxidation and inhibits LXR-SREBP-Ic- mediated lipogenesis.

VN extract-activated AMPK in the liver of HFD obese mice and HepG2 cells.
Next, to confirm whether VN extract activates AMPK and inhibits hepatic lipogenesis in a cell model, we examined the effect of VN extract on AMPK activation and LXR-SREBP-1c-mediated lipogenesis in HepG2 cells. As shown in Figure 4C, VN extract increased the phosphorylation of AMPK and ACC in a concentration-dependent manner without affecting the protein level of AMPK and ACC, suggesting that VN extract activates AMPK in HepG2 cells. Then, we assessed expression of fatty acid oxidation genes in HepG2 cells treated with VN extracts. VN extract significantly increased fatty acid oxidation genes such as CPT-1 and ACO (Fig. 4D). We also examined the effect of VN extract on LXR agonist T090137-activated SREBP-1c induction in HepG2 cells, because AMPK has been reported to inhibit LXR-mediated SREBP-1c induction. 24 T090137 treatment increased the protein and mRNA levels of SREBP-1c (Fig. 4E). However, the increases in SREBP-1c level by T090137 were antagonized by VN extract (Fig. 4E). Consistent with this, VN extract also prevented the increase in the expression of T090137-induced SREBP-1c and its target genes, including FAS, SCD-1, and S14 (Fig. 4F). These results suggest that VN extract inhibits LXR-mediated induction of SREBP-1c, which inhibits hepatic lipogenesis.
VN extract inhibited insulin or glucose-stimulated lipogenesis in HepG2 cells
High insulin or glucose is known to induce fatty liver. 25,26 To verify the inhibitory effect of VN extract on fat accumulation in HepG2 cells, we measured TG content in HepG2 cells treated with 100 nM insulin or 25 mM glucose for 24 h. As shown in Figure 5, treatment with insulin or glucose increased TG levels, whereas treatment with VN extract inhibited the increase of TG level (Fig. 5A, B; left). Consistent with the decrease in TG content, VN extract also prevented the increased expression of lipogenic genes by insulin or glucose (Fig. 5A, B; right), suggesting that VN extract inhibits insulin or glucose-stimulated fat accumulation in HepG2 cells, which is consistent with the reduction of fat accumulation in an animal model.

VN extract inhibited high insulin or high glucose-induced lipogenesis in HepG2 cells.
Discussion
Previous studies reported that Vigna species exert several pharmacological effects, 11 –14 including lowering glucose concentration and inhibiting obesity. However, antihepatic steatosis effect has not yet been reported. Therefore, this study first elucidated whether VN inhibits HFD-induced hepatic steatosis and if so, the mechanistic basis. Our finding demonstrated that VN extract treatment significantly ameliorated hepatic fat accumulation in HFD-induced obese mice, as demonstrated by reduced liver weight, hepatic TG content, and histological evaluation of liver sections by Oil Red O staining. Pathological examination of liver tissue by H&E revealed fewer lipid droplets in the livers of HFD obese mice on VN extract. Consistent with decreased fat accumulation in liver tissue, VN extract also suppressed the increased levels of TG, NEFA, TC, and the hepatic injury markers in the plasma induced in HFD obese mice, indicating that VN extract has antihepatic steatotic effects.
Hepatic fat accumulation results in oxidative stress and inflammation, which is implicated in the second hit of NASH and exacerbates liver injury and NASH. 1,2 HFD feeding promoted oxidative stress and inflammation, as shown by the increase in TBARS and the increased expression of the proinflammatory genes TNF-α, IL-1, and MCP-1. However, treatment with VN extract prevented the increase in TBARS formation and inflammatory gene expression, and significantly restored the decrease in GPx, catalase, and nonenzymatic GSH levels, suggesting that VN extract inhibits oxidative stress by upregulating antioxidant enzymes and GSH levels. According to these results, VN extract attenuates steatohepatitis induced by fat accumulation.
Although the molecular mechanisms underlying fatty liver are not fully understood, dysregulation of hepatic lipid homeostasis caused by pathological conditions, such as reduced fatty acid oxidation, enhances de novo lipogenesis, elevates hepatic fatty acid influx, and/or increases systemic insulin resistance and are considered important in the development of fatty liver. Phosphorylated AMPK cascades have emerged as novel targets in the treatment of fatty liver, because activation of hepatic AMPK leads to increases in fatty acid oxidation and simultaneously inhibits hepatic lipogenesis, cholesterol synthesis, and glucose production. 3,4 Considering the key role of AMPK activation in regulating lipid metabolism, we investigated the signaling pathway involved in AMPK activation in VN extract-treated HFD obese mice. We observed that HFD decreased phosphorylation of AMPK and ACC, whereas treatment with VN extract restored the decrease in phosphorylation of AMPK and ACC in HFD obese mice. This result was also confirmed in HepG2 cells. Consistent with AMPK activation, treatment with VN extract increased the expression of CPT-1 and ACO-1 in HepG2 cells as well as in HFD obese mice, whereas it inhibited the expression of SREBP-1c, suggesting that upregulation of fatty acid oxidation genes and downregulation of SREBP-1c are also involved in VN extract-mediated antihepatic steatosis.
LXR plays a key role in hepatic steatosis by stimulating expression of SREBP-1c and its target lipogenic genes by binding to the LXR-responsive element in the promoter regions. 27,28 AMPK has been reported to inhibit LXR by inhibiting the activation of LXR by S6K1. 24 Therefore, we examined whether VN extract inhibits S6K in livers of HFD obese mice to characterize another molecular mechanism of the protection against hepatic steatosis. HFD feeding significantly increased the phosphorylation of S6. However, treatment with VN extract decreased the phosphorylation of S6K1, indicating that VN extract inhibits LXR activity through AMPK-mediated inhibition of S6K. Moreover, we confirmed the inhibition of LXR-mediated lipogenesis by VN extract in HepG2 cells. Treatment with T01390 induced the expression of SREBP-1c and its lipogenic genes, including FAS, ACC, and S14. However, VN extract prevented the T01390-mediated increase in the expression of SREBP-1c and its lipogenic genes, demonstrating that VN extract inhibits LXR-mediated SREBP-1c-dependent lipogenesis.,
Evaluation of phenolic contents of VN extract revealed that it possessed high amounts of catechins, including (−) catechin and EGCG. Polyphenolic catechins have been reported to have hypolipidemic, thermogenic, antioxidant, and anti-inflammatory activities. EGCG reduces hepatic steatosis and the severity of liver injury in an NAFLD mice model. 20 Further, VN extract also contains rutin and quercetin. Rutin and quercetin attenuate most of the symptoms of metabolic syndrome, including NAFLD and obesity, and decrease TG synthesis and cholesterol levels in an oleic acid-induced in vitro model of fatty liver. 19,21 Therefore, the flavonoids, including EGCG, rutin, and quercetin, may contribute to antihepatic steatosis effects of VN extract.
In conclusion, VN extract prevents hepatic steatosis through activation of AMPK. Therefore, VN could be a useful, natural antihepatic steatosis agent that could reduce the risk of metabolic disorders.
Footnotes
Acknowledgments
This research was supported by the Globalization of Korean Foods R&D program, funded by the Ministry of Food, Agriculture, Forestry, and Fisheries, Republic of Korea (2012), and was supported by the Research Fund Program of Research Institute for Basic Sciences, Pusan National University, Korea, 2013, Project No. RIBS-PNU-2011-00012180001.
Author Disclosure Statement
The authors declare no conflicts of interest exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.