
Abstract
Intestinal microflora and inflammation are associated with the risk of inflammatory bowel diseases. Noni (Morinda citrifolia L.) has various bioactivities, but its effect on colon health remains unknown. This study focused on the effects of fermented noni fruit extracts on colon microflora and inflammation of colon epithelial cells. The anti-inflammatory activities of ethanol and ethyl acetate extracts on Caco-2 cells were evaluated including interleukin-8 (IL-8) and cyclooxygenase-2 (COX-2). The growth of Lactobacillus and Bifidobacterium species was promoted by ethanol extract. Ethyl acetate extract decreased intracellular reactive oxygen species and significantly suppressed COX-2, IL-8, and prostaglandin E2 production and neutrophil chemotaxis by suppressing the translocation of the p65 subunit. Quercetin was the main contributor to the anti-inflammatory activity. The fermented noni fruit promoted probiotic growths and downregulated the intracellular oxidation and inflammation in Caco-2 cells. These results suggest that fermented noni fruit might protect against inflammatory diseases of the colon.
Introduction
I
Dietary antigens, oxidative stress, and pathogens induce inflammatory responses causing intestinal immune imbalances. 6 Dysregulated expression of inflammatory molecules increase the risk of inflammation-related diseases such as colitis and colon cancer. 7 Cyclooxygenase-2 (COX-2) and interleukin 8 (IL-8) are common proinflammatory proteins, which are rapidly induced during acute or chronic inflammation. 8,9 The overexpression of COX-2 and IL-8 contributes to the formation of intestinal diseases, such as gastritis, colitis, ulcerative colitis, and colon cancer. 10,11
COX-2 widely presents in epithelia, endothelia, and immune cells, and directly regulates the production of prostaglandin E2 (PGE2), with multiple signals involved in inflammation. 10,12 High levels of COX-2 are found in local tissues and organs from patients with inflammatory bowel disease (IBD), ulcerative colitis, and Crohn's disease. 13 COX-2 is induced by lipopolysaccharide (LPS) and interleukin-1 beta (IL-1β) that promote immune signals to protect against foreign invasion. 12,14 IL-1β, an endogenous inflammatory molecule, is produced by immune cells such as macrophages. Long-term induction of IL-1β would cause severe inflammation and increase the risk of inflammation-associated diseases. 12,14
IL-8 is an α-chemokine, which can be secreted from macrophages, endothelial and epithelial cells. IL-8 plays an important role in the progress of gastrointestinal diseases. 15 IL-8 prompts neutrophils to eliminate foreign antigens, and activates acquired immune responses. 16 Bacterial molecules, such as LPS and flagellin, increase the secretion of IL-8 protein by activating the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) or activator protein-1 (AP-1) pathways on epithelial cells. 15,17 IL-8 protein increases the amount of neutrophils, and promotes the activation of transcription factors, such as NF-κB and hypoxia-inducible factor 1, which enhance inflammation or change cell growth. 17 High levels of IL-8 protein were found in the colon mucosa of patients with IBD. 18 Thus, long-term exposure of IL-8 protein would cause neutrophil infiltration and severe inflammation, thus increasing the risk of colon cancer. 19 Moreover, the oxidative stresses would induce inflammatory signals, such as NF-κB to enhance the production of proinflammatory cytokines. 20 Thus, elimination or inhibition of the production of reactive oxygen species (ROS) are appropriate strategies to suppress the synthesis of proinflammatory molecules.
Noni trees (Morinda citrifolia L.) are widely distributed from southeast Asia to Australia, and its fruit juice is commonly used as a beverage. 21 The extracts of noni leaf and root have been shown to activate AP-1 and prevent edema formation in carrageenan-induced rats. 22,23 Noni root extract regulates accumulation of macrophages to prevent colitis in dextran sulfate sodium-induced rats. 24 Moreover, noni fruit juice regulates inflammation and selectively inhibits cyclooxygenase enzymes. 25 The ethyl acetate extract of noni fruit could inhibit COX-2 and inducible nitric oxide synthase (iNOS) expression in LPS-induced macrophages. 26 Moreover, the extract of fermented noni juice shows great scavenging capacity against various free radicals. 27 The juice of noni fruit also provides hypoglycemic and hepatoprotective effects in diabetic rats. 28 In a previous study, the extracts of fermented noni fruit show great antiadhesive and anti-inflammatory properties during Helicobacter pylori infection, which were reflective of the changes in phenolic compounds after fermentation. 29 Therefore, fermented noni fruit is considered beneficial to human health but the effects of fermented noni fruit on colon function has never been reported.
In this study, the effect of extracts of fermented noni fruit on colon microflora was examined. The phenolic compounds and antioxidative activities of both extracts were also characterized. Both extracts on COX-2 and IL-8 signals of human colon epithelial cells were determined in this study.
Materials and Methods
Preparation of ethanol and ethyl acetate extracts
The mature noni (M. citrifolia L.) fruit (original strains from Hawaii and cultured in Taiwan) was purchased from a local farm in November 2012. The noni fruit was cleaned and processed using natural fermentation in an air-tight jar (10 pieces/25 L) at 27°C, 70–80% relative humidity for 3 months. After fermentation for 3 months, the fermented noni fruit (both fruit and juice were collected) was crushed in a homogenizer and dried in a heated air circulation proofer at 37°C. The dry powder of fermented noni fruit was stored at 7°C before extraction. One hundred grams of fermented noni fruit powder was extracted with 1 L of 95% ethanol or 99% ethyl acetate for 24 h at room temperature. The residues were removed by filtration and extracts were evaporated to dryness under vacuum at 37°C. Both extracts were freeze-dried for 48 h to remove moisture. The dried extracts were transferred into air-tight amber bottles and stored at −80°C until use. Both extracts were completely dissolved in culture medium containing dimethyl sulfoxide (DMSO; final concentration <0.02%) before use in the cell and microflora experiments. DMSO controls were included in relevant experiments.
Colon microflora determination
Human stool sample from a healthy volunteer (10 g of fresh stool was stored in a sealed anaerobic package and then packed in a round plastic bottle with screw cap. After collection, the case was placed in a sealed plastic bag with CO2 anaerobic pack) was collected (within 4 h) and mixed with phosphate-buffered saline (PBS) buffer (1.44 g Na2HPO3, 8.18 g NaCl, 0.204 g KH2PO4, and 0.201 g KCl, pH 5.0) at a ratio of 1:10. All diluent stool samples (20 μL) were smeared on selective and differential medium, and cultured with various concentrations (0.04, 0.08, 0.16, 0.31, and 0.63 mg/mL) of fermented noni fruit extracts in anaerobic condition for 48 h at 37°C. The selective and different media were used in this study respectively (Bifidobacteria iodoacetate medium-25 agar for Bifidobacteria species, Rogosa SL agar for Lactobacillus species, MacConkey agar for E. coli, and Tryptose sulfite D cycloserine agar for C. perfringens). All colony-forming units were recorded and compared with those in various concentrations of fermented noni fruit extracts.
Contents of total phenolics and phenolic profile assays by high performance liquid chromatography
First, 50 μL extract or standard solution was mixed with 1 mL of H2O and 0.5 mL of Folin–Ciocalteu phenol reagent (no.109001; Merck). Then, 2.5 mL of 20% Na2CO3 solution was added to the mixture, followed by incubation at room temperature in the dark for 20 min. The absorbance against a blank was measured at 735 nm. Gallic acid was used to prepare a standard curve. 30
The phenolic compounds were separated using high performance liquid chromatography (HPLC) with a binary gradient intelligent pump (L6200A; Hitachi) and a UV-VIS detection system (L4250; Hitachi). A Lichrospher 100 RP-18e (Merck) column (5 μm, 25 cm×4 mm i.d.) and security guard RP-18e (5 μm, 4×4 mm i.d.) were used. The mobile phase consisted of 2.5% (v/v) acetic acid water solution (solvent A) and acetonitrile (solvent B). The gradient program was obtained according to a previous study. 29 Chlorogenic acid (no. C-3878; Sigma), coumaric acid (no. C-9008; Sigma), gallic acid (no. G-7384; Sigma), myricetin (no. M-6760; Sigma), naringin (no. 71162; Sigma-Aldrich), quercetin (no. Q-0125; Sigma), and kaempferol (no. K0133; Sigma) were standards used for quantification of extract phenolics. The polyvinylpyrrolidone (PVP; Sigma) was used to remove all phenolic compounds to further verify the contribution of phenolic compounds.
Trolox equivalent antioxidant capacity
It was based on the suppression of the absorbance of radical captions of 2,2′-azinobis (3-ethylbenzothiazoline 6-sulfonate; ABTS) by antioxidants in the test extracts when ABTS incubates with a peroxidase and H2O2. A reaction mixture containing 20 μL of H2O2 (50 μM), 100 μL of peroxidase (4.4 U/mL), ABTS (100 μM), and a water-soluble derivative of vitamin E, Trolox (concentration range, 0–0.6 mM) as standard was incubated at 37°C for 1 min, absorbance changes were monitored at 734 nm. 31
Low-density lipoprotein oxidation lag time assay
The human peripheral blood (7 mL) was collected in a heparin-serum separation tube from healthy volunteer. The 7 mL of peripheral blood was centrifuged at 906 g for 5 min to obtain serum (3 mL). The 0.5 mL of NaBr solution (density=1.097) was added to 3 mL of serum to centrifuge (97,803 g) at 4°C for 16 h (to remove intermediate-density lipoprotein and very low-density lipoprotein [LDL]). After centrifugation, the 1 mL of suspension was mixed with 0.5 mL of NaBr solution (density=1.283) to centrifuge (97,803 g) at 4°C for 20 h. The LDL was obtained from the suspension after the second centrifugation. The LDL was processed by dialysis in PBS (8.2 g NaCl, 4.55 g Na2HPO4·H2O, and 0.17 g NaH2PO4·H2O, pH 7.5) solution over night to remove other metal ions. Extracts or ascorbic acid were mixed 1.2 mL of 0.2 mg/mL LDL (sample final concentration about 200 μg/mL) and added 6 μL of 1 mM CuSO4. Absorbance was monitored at 5 min intervals at 234 nm and compared to the start time of oxidation.
Cell lines and culture condition
The human colon epithelial cell line Caco-2 (ATCC; HTB-37) was used in this study. Caco-2 cells were cultured with Eagle's minimum essential medium containing 15% fetal bovine serum and 1% penicillin-streptomycin at 37°C in a humidified incubator containing 5% CO2.
Cell viability assay
Effects of both extracts on cell viability were investigated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT). Caco-2 cells were seeded in 96-well plate with 2×105/mL cells and 200 μL medium in each well overnight. The cells were treated with various concentrations of fermented noni extracts (25, 50, 100, or 200 μg/mL) at 37°C for 24 h. The medium was replaced by a fresh medium, containing 0.5 mg/mL MTT, and cultured at 37°C for 4 h. Then, 100 μL isopropanol was added to each well to dissolve the crystals. The optical density of each sample was read at 630 nm, against the blank prepared from the cell-free well.
Intercellular ROS determination by 2,7-dihydrodichlorofluorescein diacetate
Caco-2 cells were seeded in 96-well dish with 1×105/mL. About 50 μg/mL of LPS was cocultured with extracts for 24 h. To remove supernatant and wash three times, the medium containing 100 μM 2,7-dihydrodichlorofluorescein diacetate (H2DCF-DA) (Invitrogen) was added to each well and cultured for 30 min at 37°C. H2DCF-DA would be converted to 2′,7′-dichlorofluorescein (DCF) by ROS. The DCF was measured by fluorescence measurement system at 485 nm (excitation max) and 530 nm (emission max).
The expression of COX-2 and MyD88 and phosphorylation of IRAK-1, p38 MAPK, and jun N-terminal kinase assay
Caco-2 cells were seeded in 10-cm dish with 1×105/mL. IL-1β protein (lot. B138076; BioLegend) was added and co-cultured with extracts for 24 h. To collect the cells, the cells were disintegrated using a lysis buffer (no.9803; Cell Signaling). Cell proteins were collected after centrifuged (6708 g) for 10 min at 4°C. All proteins were measured by western blot. 29 COX-2 antibody (no. ab52237; Abcam), myeloid differentiation primary response gene 88 (MyD88) antibody (no. 4283S; Cell Signaling), phospho-interleukin-1 receptor-associated kinase 1 (IRAK-1) (Thr209) antibody (no. 12756S; Cell Signaling), Phospho-p38 mitogen-activated protein kinase (MAPK)-Thr180/Tyr182 antibody (no. 9216S; Cell Signaling), and Phospho-SAPK/JNK-Thr183/Tyr185 antibody (no.9251S; Cell Signaling) were used. The raw data of western blot were quantified by FUJIFILM LProcess (version 1.96). The target protein (COX-2) was corrected with the housekeeping protein (Actin), and the value was divided by induced group (IL-1β). The final value was expressed as percentage. All the results of western blot were obtained from at least five batches of culture, and triplicates for each culture.
NF-κB p65 subunit protein translocation assay
Caco-2 cells were seeded in 10-cm dish with 1×105/mL. After the extracts were treated with IL-1β protein or LPS and cultured for 24 h, the total nuclear protein was measured by nuclear protein isolation translocation assay kit (NPI-1; FIVEphotonBiochemicals). The p65 subunit protein was measured by western blot. 29 NF-κB p65 subunit antibody (no. GTX102090; GenTex) was used and lamin B1 (no. GTX103292; GenTex) served as an internal control of nuclear protein.
IL-8 and PGE2 protein secretion assay
Caco-2 cells were seeded in 10-cm dishes with 1×105/mL. LPS (from E. coli) or IL-1β was added and cocultured with extracts for 24 h. The supernatants were collected and the levels of IL-8 protein were determined using a human IL-8 protein commercial ELISA kit (no. K0031216; Koma Biotech, Inc.), while PGE2 protein was determined by human PGE2 express EIA kit (no. 500141; Cayman).
Neutrophil migration assay
The neutrophil isolation was described in a previous study. 29 Neutrophil chemotaxis was determined by two-chamber techniques. First, 30 μL medium of LPS-treated AGS cells or various concentrations of extracts were poured into the lower layer. The IL-8 protein (600 pg/mL) was used as a positive control. Then, 50 μL neutrophils (2×105/mL) were added to the upper layer. A polycarbonate-free membrane was placed between the lower and upper layers and cultured for 1 h at 37°C. The degree of neutrophil migration was calculated by using Giemsa stain.
Statistical analysis
All results were presented as the mean±SD (n=3). The differences were assessed by one-way ANOVA. Bonferroni correction and Duncan (post hoc analysis) were performed for multiple comparisons. A difference with P<.05 was considered significant (version 12.0; SPSS, Inc.).
Results and Discussion
Effects on colon microflora
Probiotics such as Bifidobacterium and Lactobacillus species regulate inflammation and enhance immunity to promote health. 32 On the other hand, pathogenic bacteria such as C. perfringens and E. coli can cause diarrhea, inflammatory disorders, carcinogenesis, and putrefaction. 32,33 Hence, the effect of fermented noni fruit extracts on Bifidobacterium species, Lactobacillus species, C. perfringens, and E. coli was examined.
Our results revealed that the noni ethanolic extract significantly promoted the growth of Bifidobacterium and Lactobacillus species, (0.16–0.63 mg/mL for Bifidobacterium and Lactobacillus spp.) whereas the ethyl acetate extract was ineffective (Fig. 1A, B). Noni extract had no effect on the growth of C. perfringens and E. coli (Fig. 1C, D).
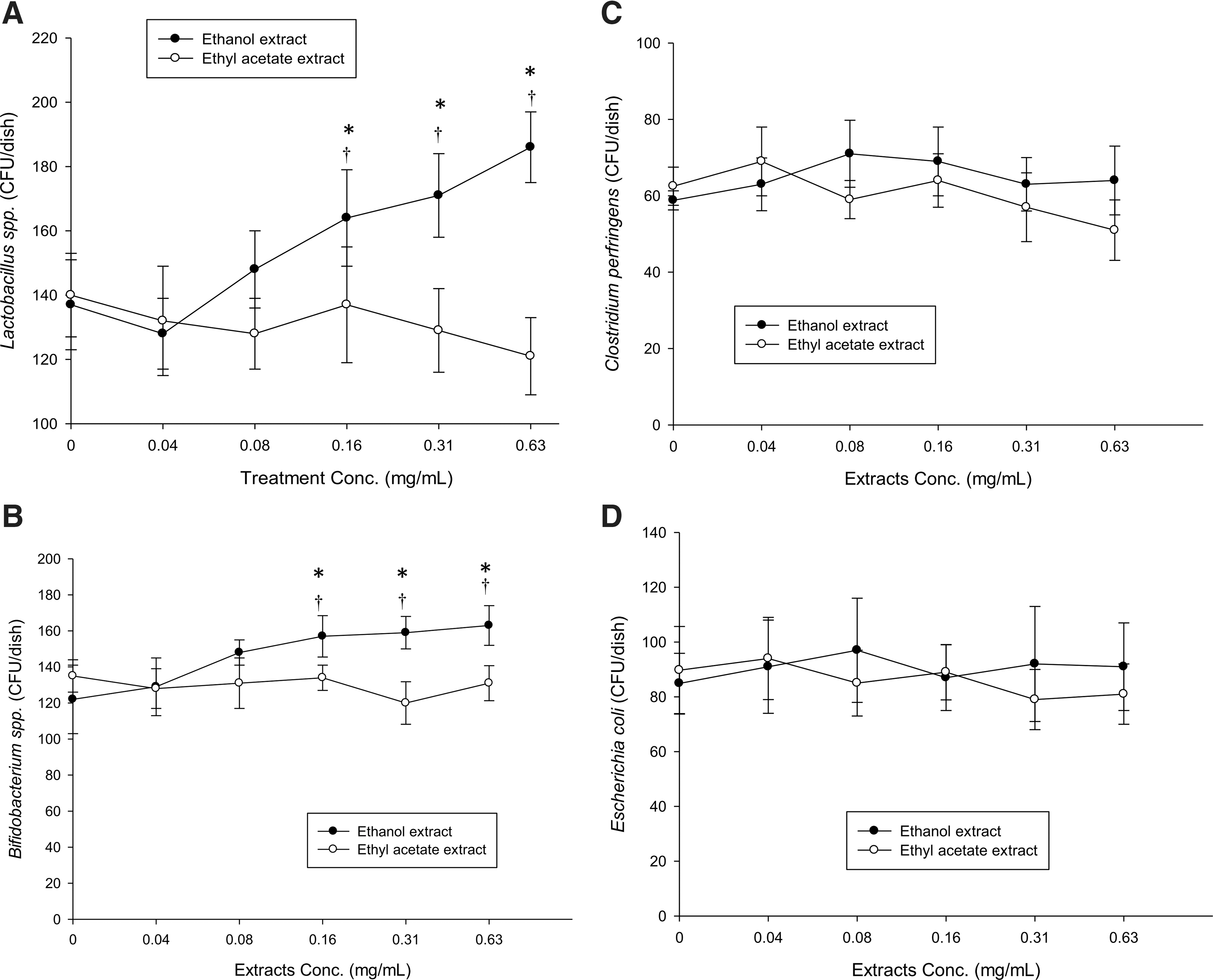
Effects of ethanol and ethyl acetate extract on growth of Lactobacillus spp.
Most prebiotics, such as fructo-oligosaccharides or inulin, are water soluble or highly polar, which stimulates the growth of probiotics. 34,35 Noni fruit juice increases the number of Bifidobacterium longum and Lactobacillus plantarum after 48 h of fermentation. 36 Some phenolic compounds can regulate the growth or colonization of bacteria, such as quercetin and proanthocyanidin. 29,37
Previous studies indicated that phenolic compounds can inhibit pathogen growth, such as E. coli and H. pylori. 29,38 On the other hand, various phenolic compounds also inhibit probiotic growth. 38 Those studies suggest that phenolic compounds are probably unsuitable as prebiotics, even though phenolics can suppress pathogen growth. Moreover, noni fruit has been found to contain various phenolic and flavonoid compounds. 39,40 In this study, the fermented noni fruit extracts did not suppress probiotic growth (Fig. 1A, B). Thus, the phenolic compounds in both extracts were measured to clarify their effect on probiotic growth.
Phenolic profiles in fermented noni fruit extracts
The ethanol extract contained 45.68±3.21 mg/g of total phenolics and 39.21±2.21 mg/g of total flavonoid; and the ethyl acetate extract contained 70.21±1.42 mg/g of total phenolics and 44.68±1.25 mg/g of total flavonoids.
The phenolic composition of both extracts was determined by HPLC-UV. Both extracts contained various phenolic compounds, including chlorogenic acid, coumaric acid, gallic acid, naringin, myricetin, quercetin, and kaempferol (Fig. 2A, B). Ethanol extract contained abundant coumaric acid (21.3 mg/g extract) and quercetin (4.4 mg/g extract), whereas the ethyl acetate extract also contained high levels of quercetin (34.6 mg/g extract) (Table 1). When both extracts were treated with PVP to remove all the phenolic compounds and further cultured with Bifidobacterium, Lactobacillus species, C. perfringens, and E. coli the ethanol extract lost its ability to promote growth of Bifidobacterium and Lactobacillus species (data not shown). Those results indicated that the phenolics in fermented noni fruit extract exerted it prebiotic effects.
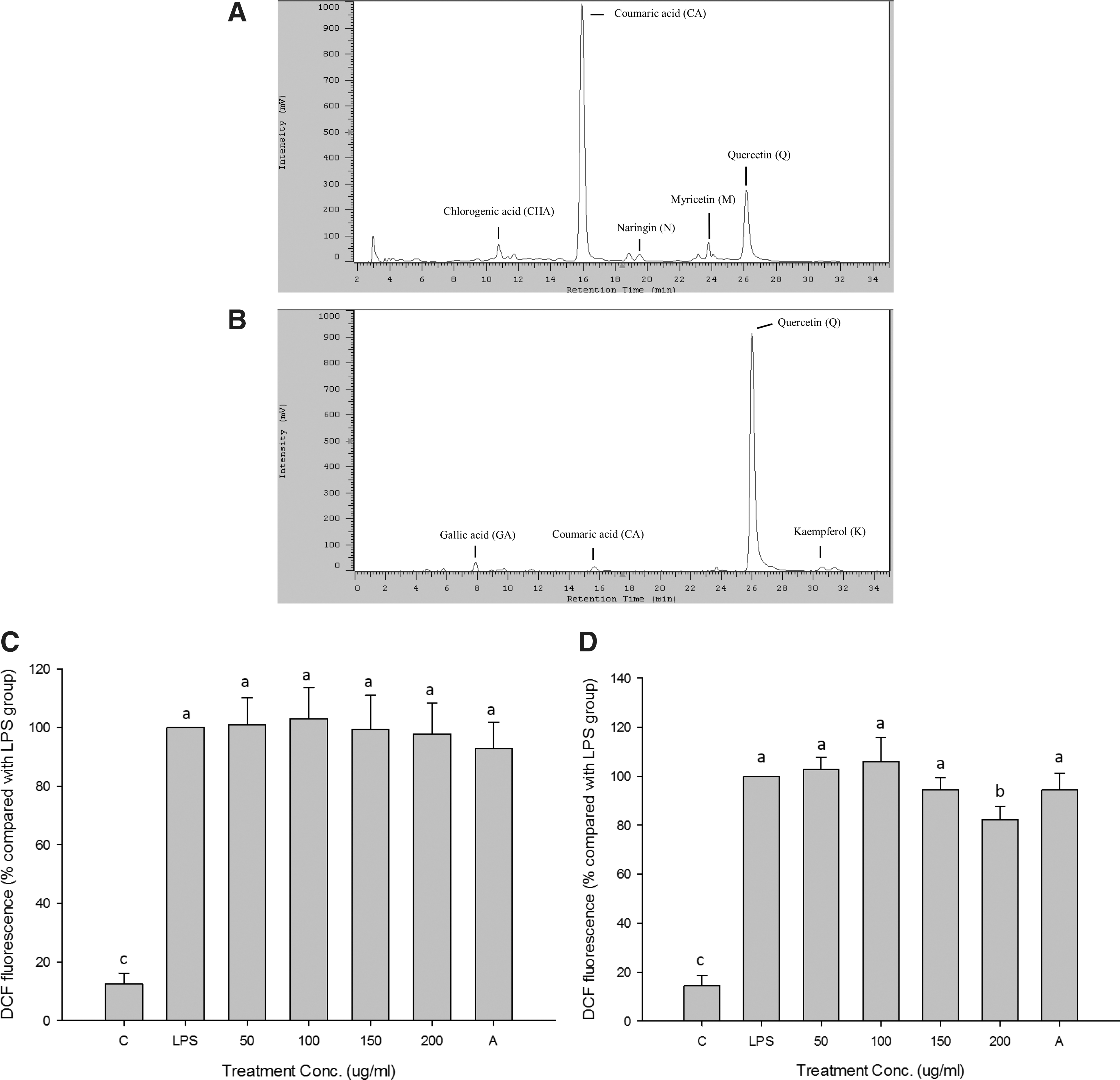
High performance liquid chromatography profiles of ethanol
All results represent the mean±SD (n=3).
Antioxidative activity and intracellular ROS inhibition
The excessive release of free radicals and ROS during infection or inflammation can cause tissue destruction or cancer induction. 41,42 Oxidative stress enhances inflammatory signals and exacerbate tissue damage. Moreover, some pathogens such as H. pylori secrete hydrogen peroxide causing oxidative damage in the gastrointestinal tract. 43 Fermented noni fruit extracts contained abundant phenolic compounds act as antioxidants and protect against oxidative damage or relieve inflammation. Accordingly, the antioxidative activities of both extracts were measured.
Ethyl acetate extracts exhibited the best total antioxidant activity, LDL protection, and anti-inflammatory properties (Table 2). Ethyl acetate showed better effects than ethanol extract on total antioxidant activity and LDL protection because of abundant phenolics in the ethyl acetate extract (Table 1). Furthermore, the effects of both extracts on intracellular ROS of Caco-2 cells induced by LPS were measured. Both extracts showed no effects on cell viability (25–200 μg/mL) after 24 h of culture (data not shown). The results showed that ethanol extract did not affect intracellular ROS (Fig. 2C), but ethyl acetate extract (200 μg/mL) significantly decreased ROS in LPS-induced Caco-2 cells (Fig. 2D). The intracellular ROS could be induced during inflammation to enhance various inflammatory signals, such as NF-κB and AP-1. 20,44 Ethyl acetate extract may impede the loop of inflammatory response induced by ROS to relieve the production of proinflammatory molecules.
All values expressed as mean±SD (n=3).
Different letters are significant difference.
LDL, low-density lipoprotein.
Inhibition on COX-2 expression
IL-1β protein (human) is an endogenous inducer of COX-2 expression, while LPS (from E. coli) is an exogenous inducer for stimulating IL-8 production. The production of COX-2 and IL-8 proteins showed dose-dependent trends with IL-1 and LPS concentrations. Hence, 10 ng/mL of IL-1β and 50 μg/mL of LPS were used to induce COX-2 and IL-8 expression in the following studies. The ethanol extract showed no effect on COX-2 expression (Fig. 3A). In contrast, high doses of ethyl acetate extract (100 and 200 μg/mL) markedly reduced IL-1β-induced COX-2 expression (no significant difference as compared with the control group), but low doses of ethyl acetate extract (25 and 50 μg/mL) showed no effect (Fig. 3B).
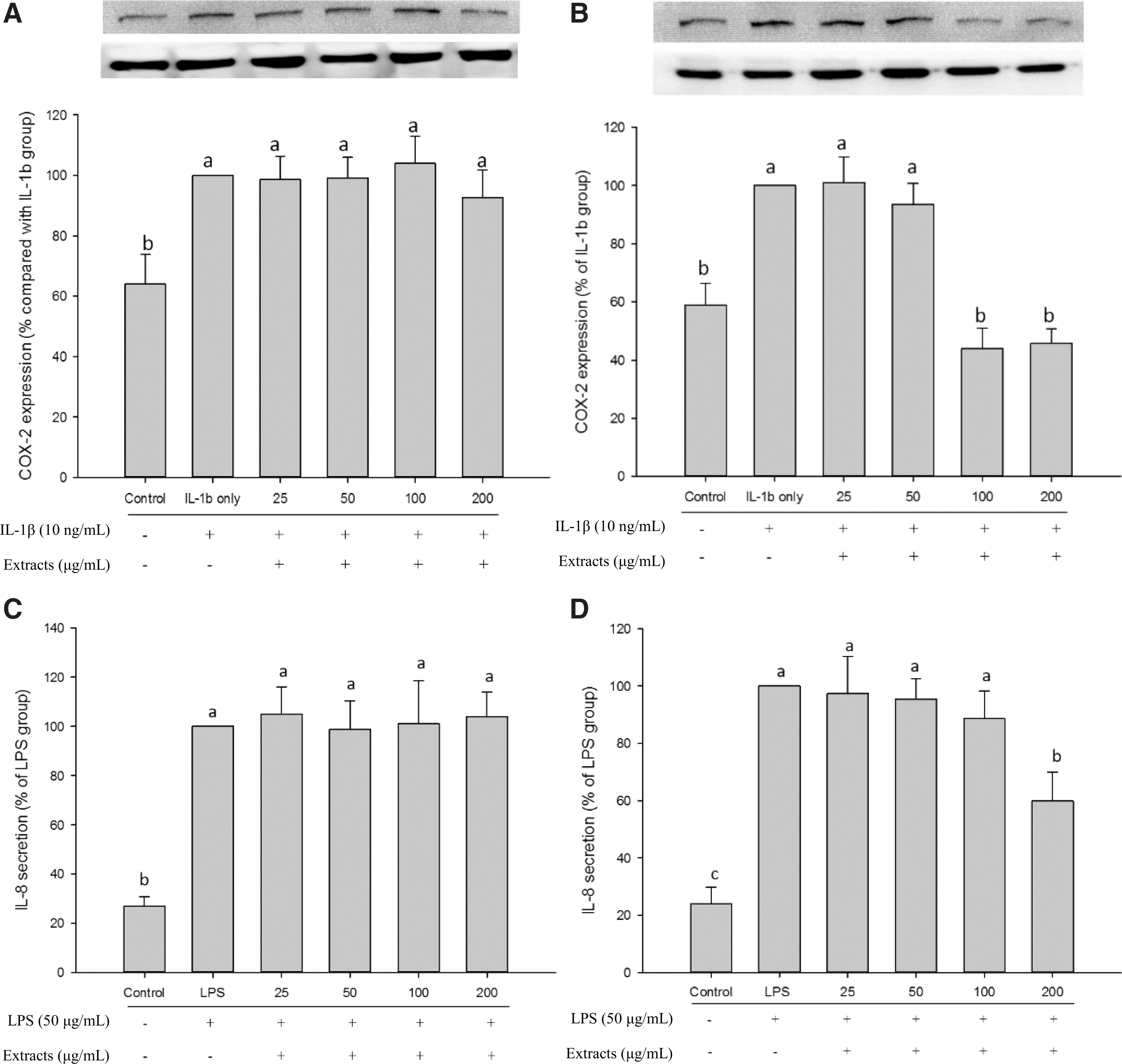
COX-2 and IL-8 expression of Caco-2 cells by IL-1β or LPS-induced after extracts treated for 24 h. The ethanol extract on COX-2
COX-2 catalyzes the production of precursors of inflammatory molecules, such as PGE2. 12 Previous research indicated that COX-2 overexpression could increase the risk of IBD. 13 In a normal state, COX-2 is rapidly induced by bacterial infection, which could increase the secretion of gastric acid and intestinal fluid to block bacterial invasion. 10 Macrophages produce IL-1β protein during microbial infection. IL-1β stimulates the immune responses and increases COX-2 expression. 14,45 However, overexpression of COX-2 can cause intestinal damage. COX-2 and PGE2 inhibitors are used to block COX-2 signals to reduce the risk of colitis, IBD, and colon cancer. However, these agents probably cause unknown sequela or side effects, such as ulceration. 13 In this study, the ethyl acetate extract of fermented noni fruit mitigated COX-2 expression (Fig. 3B), thus showing high potential as a safe substitute for anti-inflammatory drugs.
Inhibition on IL-8 secretion
The ethanol extract had no effect on IL-8 secretion (Fig. 3C). However, IL-8 secretion was significantly reduced by 200 μg/mL of ethyl acetate extract (Fig. 3D). These results were similar to the effects on COX-2 expression (Fig. 3A, B).
IL-8 is a neutrophil chemotactic factor that causes neutrophil chemotaxis and accumulation. 16 IL-8 mediates the activation of polymorphonuclear leukocytes and further regulates inflammatory responses. In a normal state, IL-8 not only reinforces immunization to eliminate foreign antigens, but it could also cause tissue damage during long-term secretion. Indeed, the overexpression of IL-8 increases the risk of gastric ulcer, colitis, Crohn's disease, and IBD. 3,19,45,46 Moreover, IL-8 also regulates the adhesion, migration, and invasion of cancer cells. 17 The ethyl acetate extract significantly suppressed LPS-induced IL-8 secretion (Fig. 3D), indicating that ethyl acetate extract has the potential to relieve IL-8 secretion and to disrupt the excessive chemotaxis of neutrophils during bacterial infection.
Inhibition on IL-8 and COX-2 by NF-κB pathway
The ethyl acetate extract was a potent inhibitor of COX-2 and IL-8 protein expressions (Fig. 3B, D). Accordingly, the signal pathways of COX-2 and IL-8 were determined to verify the possible mechanisms involved.
COX-2 expression is dominated by p38 mitogen-activated protein kinase (p38 MAPK), NF-κB, and jun N-terminal kinase (JNK) pathways. 47,48 These signal pathways activate transcription factors, such as NF-κB and AP-1 to enhance COX-2 expression. 49 –51 IL-1β mediates the COX-2 expression through NF-κB and AP-1 to cause inflammation and oncogenesis. 1,14 IL-8 secretion is regulated by MAPK and IκB pathways activate AP-1 and NF-κB. 46,52,53 Thus, the ethyl acetate extract might regulate NF-κB or AP-1 by inhibiting COX-2 and IL-8 production.
p38-MAPK and JNK are involved in AP-1 activation. 54 p38-MAPK would carry out phosphorylation during LPS or IL-1β treatment that dominates AP-1 activation. On the other hand, AP-1 can also be activated by JNK phosphorylation to enhance the expression of inflammatory cytokines. 55 Both p38-MAPK and JNK pathways can activate AP-1 to induce COX-2, iNOS, and IL-8 expressions. Similarly, IκB phosphorylation, which induces translocation of p65 and p50 subunits, combines with NF-κB to promote transcription of target genes. 12 Thus, the phosphorylation of p38-MAPK and JNK, and translocation of p65 subunit were measured to explore the inhibition pathway of ethyl acetate extract.
LPS and IL-1β can induce phosphorylation of p38-MAPK and JNK, and cause translocation of the p65 subunit (Fig. 4). The results showed no significant change in phosphorylation of p38-MAPK and JNK after ethyl acetate extract treatment (Fig. 4A–C). In contrast, the ethyl acetate extract could suppress the translocation of p65 subunit regardless whether it was induced by LPS or IL-1β (Fig. 4D). p65 translocation showed a trend similar to the production of COX-2 and IL-8 but not in the phosphorylation of p38-MAPK and JNK after ethyl acetate extract treatment (Figs. 3B, D and 4). Thus, the ethyl acetate extract could inhibit COX-2 and IL-8 expression through suppressing NF-κB activation.

The changes
The ethyl acetate extract suppressed COX-2 and IL-8 by the NF-κB pathway but the main contributors were still unknown. To clarify the influence of phenolics on the inhibition of COX-2 and IL-8 production, PVP was mixed with the ethyl acetate extract to remove the phenolics. The PVP-treated ethyl acetate extract (EA-P) lost its inhibitory effect on COX-2 and IL-8 (Fig. 5), indicating that phenolics play crucial roles in the inhibition of COX-2 and IL-8 production.
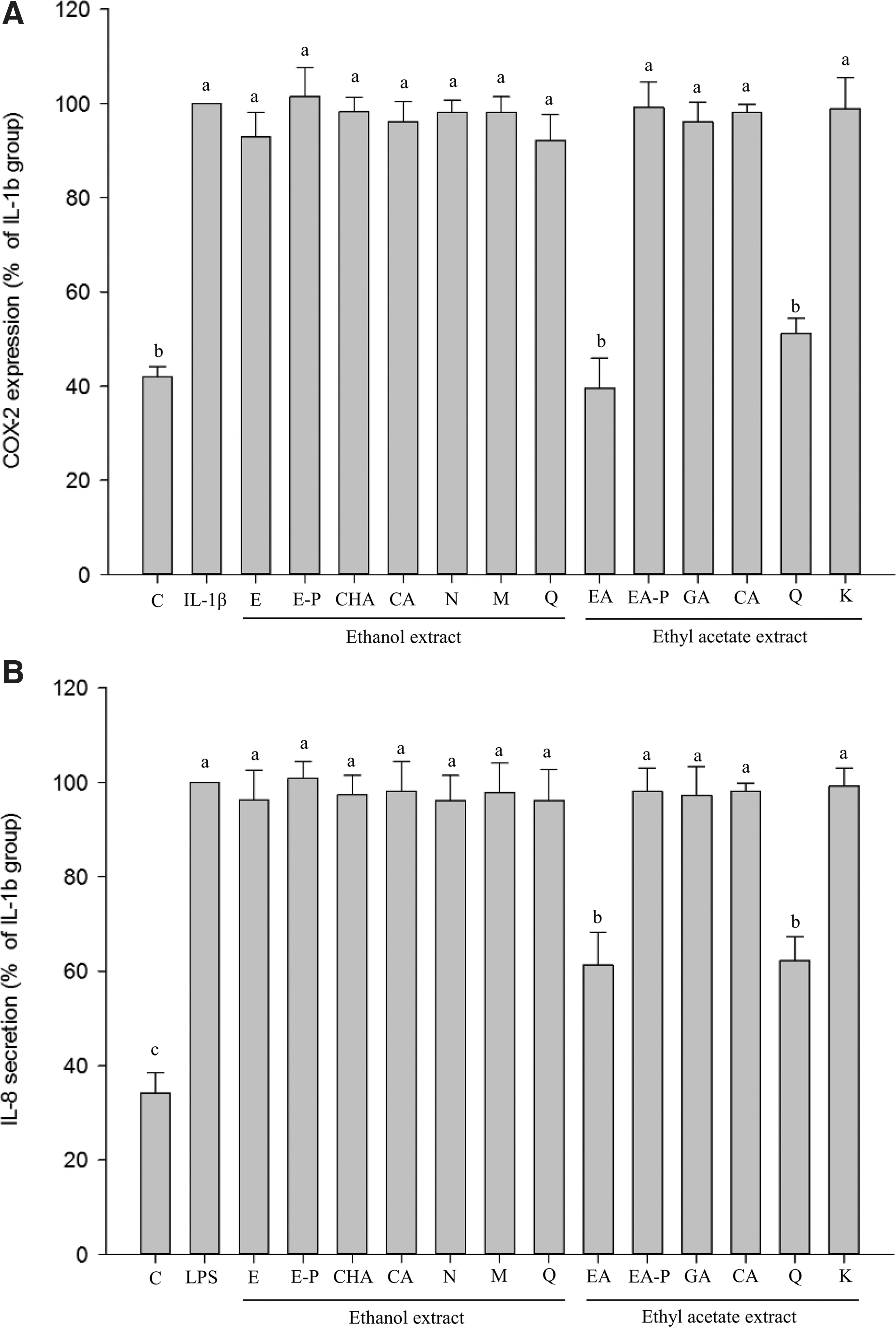
The production of COX-2
To examine the main contributors to increased COX-2 and IL-8 production, pure phenolic compounds were used. Each phenolic in both extracts was prepared as the equivalent levels in 200 μg/mL of individual extract. About 0.62 μg/mL of chlorogenic acid (CHA), 4.26 μg/mL of coumaric acid (CA), 0.49 μg/mL of naringin (N), 0.3 μg/mL of myricetin (M), and 0.88 μg/mL of quercetin (Q) were equivalent to ethanol extract; whereas 1.17 μg/mL of gallic acid (GA), 0.52 μg/mL of coumaric acid, (CA), 6.92 μg/mL of quercetin (Q), and 0.62 μg/mL of kaempferol (K) were equivalent to 200 μg/mL of ethyl acetate extract.
All the equivalent contents of phenolic compounds in the ethanol extract showed weak suppression of COX-2 and IL-8 (Fig. 5). Not surprisingly, coumaric acid exhibited weak anti-inflammatory activity even though it is a potent antioxidant. 56,57
In contrast, the equivalent quercetin in the ethyl acetate extract (6.92 μg/mL quercetin of 200 μg/mL ethyl acetate extract) strongly inhibited COX-2 and IL-8 (not significantly different as compared with control group) (Fig. 5), clearly indicating that quercetin in ethyl acetate could be the major contributor to the inhibition of COX-2 and IL-8.
Quercetin showed significant inhibition of p65 translocation during LPS and IL-1β treatment (not significantly different as compared with the ethyl acetate extract-treated group) (Fig. 6B, E). Quercetin also inhibited COX-2 and PGE2 production but the effect was weaker than that of the ethyl acetate extract-treated group (Fig. 6A, C). The effect of quercetin on neutrophil chemotaxis was measured. IL-8 protein and LPS caused migration of numerous neutrophils (Fig. 6F). Quercetin showed powerful inhibition of IL-8-induced neutrophil migration (no significant difference as compared with the ethyl acetate extract-treated group) (Fig. 6F). Results also showed no effects on signals of COX-2 and IL-8 when ethyl acetate was treated with PVP (EA-P) (Fig. 6). All the results showed that quercetin was the major contributor to the inhibition of IL-8 and COX-2 production by NF-κB pathway, which further suppressed PGE2 production and neutrophil migration.
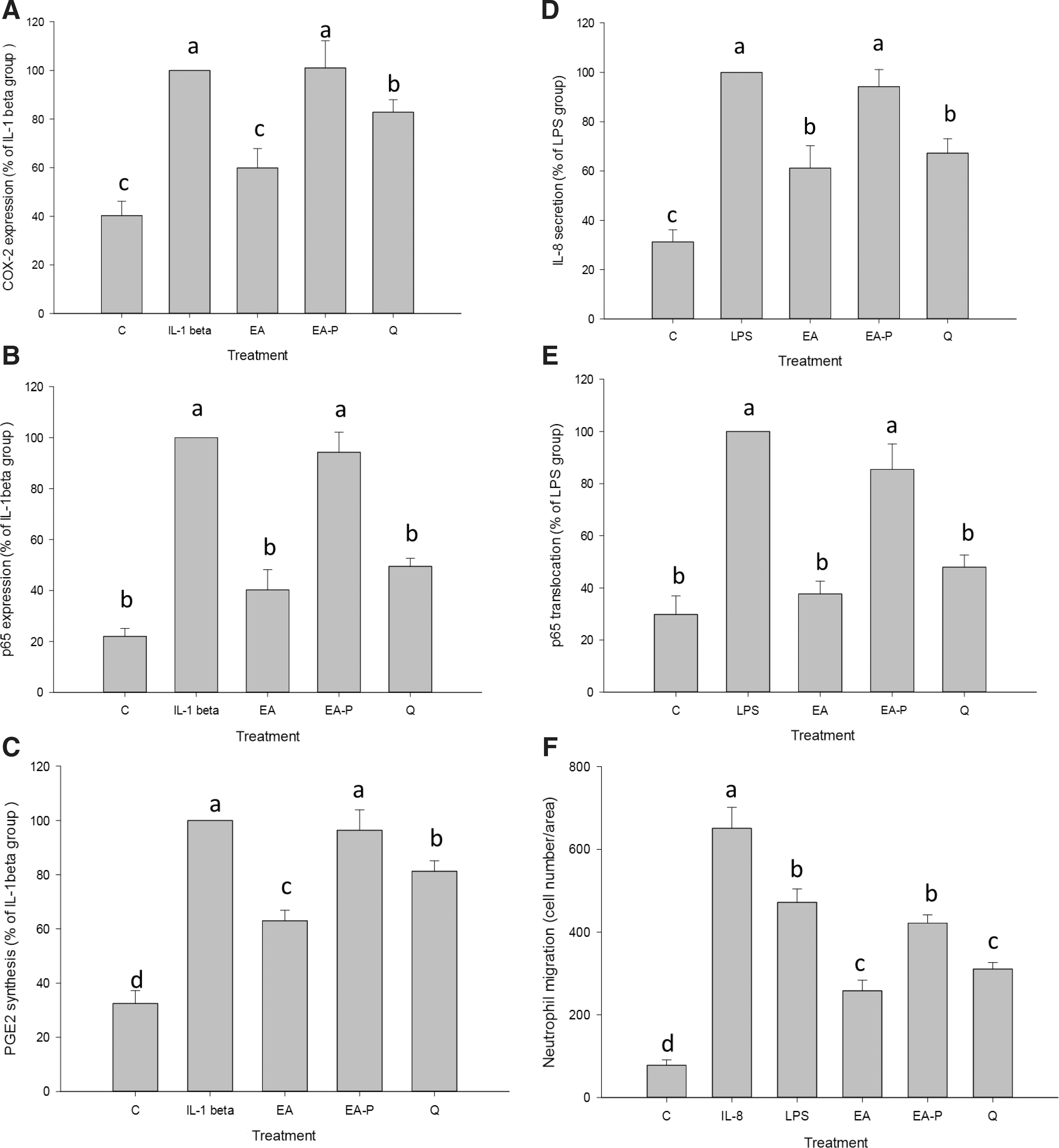
COX-2 and IL-8 signals in Caco-2 cells cultured with treatments after induction with IL-1β or LPS for 24 h. Effects of treatment on COX-2 expression
Quercetin is known to inhibit the expressions of inflammatory cytokines including TNF-α, IL-1β, IL-6, and COX-2 by NF-κB pathway on human immune cells and human intestinal cells. 58,59 Similar results were also obtained in this study, showing that the ethyl acetate extract effectively inhibited COX-2 signal including upstream of NF-κB and downstream of PGE2 induced by IL-1β (Fig. 6A–C). Thus, quercetin relieved the inflammation of colon cells from an endogenous inducer such as macrophages to avoid excessive immune responses. Nevertheless, both COX-2 expression and PGE2 production of the quercetin-treated group (Q) were higher than ethyl acetate extract-treated group (Fig. 6A, C), suggesting that unknown components in the ethyl acetate extract could affect or interact with quercetin to regulate COX-2 signaling through other pathways.
The ethyl acetate extract suppressed IL-8 secretion and further inhibited LPS-induced neutrophil migration (Fig. 6D, F). Continuous infiltration of neutrophils can cause tissue damage (tight junction impairment). 60 IL-8 induces neutrophil accumulation and strengthens IL-8 secretion, thus causing intense inflammation and increasing the risk of gastrointestinal diseases. 15 The ethyl acetate extract blocked IL-8 secretion and decreased the risk of inflammation-related diseases. In this study, quercetin significantly suppressed IL-8 secretion (Fig. 6D). Previous studies also show that quercetin regulates IL-8 secretion in the same cell line. 61,62 Quercetin in fermented noni fruit was found to play a critical role in the suppression of IL-8 secretion.
To further investigate the feasible mechanisms of ethyl acetate extract inhibition of NF-κB, the upstream signal was measured. The interaction of IRAK-1 and MyD88 molecules is the first step of NF-κB activation. LPS and IL-1β can bind with the dedicated receptor, namely Toll-like receptor (TLR) to activate the phosphorylation of IRAK-1. The phosphorylation IRAK-1 would disconnect from MyD88 and further induce IκB phosphorylation and ubiquitination. The p50 and p65 subunits translocate to the cell nucleus from cytoplasm to further activate NF-κB after IκB degradation. 63 –65 Thus, the phosphorylation of IRAK-1 could be decreased when ethyl acetate extract impedes the cross-linking or activation of LPS or IL-1β with TLR by extracellular action. Conversely, IRAK-1 would conduct phosphorylation if ethyl acetate extract inhibits NF-κB activation by midstream or downstream intracellular pathways during LPS or IL-1β-induced.
The transcription of the p65 subunit was suppressed when ethyl acetate extract was used in combination with LPS or IL-1β (Fig. 7A, B). The phosphorylation of IRAK-1 was identical to levels after ethyl acetate extract treatment (Fig. 7A, C). The expression of MyD88 was not changed by various treatments (Fig. 7A, D). These results indicated that ethyl acetate extract suppressed COX-2 and IL-8 signals by changing the intracellular signal of NF-κB, but not by extracellular interference. The quercetin-treated group also showed results similar to ethyl acetate extract in IRAK-1 phosphorylation (data not shown). Quercetin is a potent inhibitor of inflammation, which suppresses COX-2 expression and transcription by multiple pathways. 66,67 In this study, quercetin also showed strong inhibition on COX-2 by the NF-κB pathway. Moreover, quercetin was not a single contributor to COX-2 inhibition even though quercetin inhibited the translocation of p65 subunit (Fig. 6). Both ethyl acetate extract and quercetin showed analogous inhibition of IL-8 signaling (Fig. 6D–F). Hence, the ethyl acetate extract may provide comprehensive inhibition on the expressions of inflammatory cytokines by NF-κB pathway. The ethyl acetate extract of noni fruit was found to be a potent inhibitor of COX-2 and iNOS expressions in RAW 264.7 cells induced by LPS. 26 Thus, the ethyl acetate extract of fermented noni fruit probably possesses anti-inflammatory properties in other immune and epithelial cells via similar pathways.
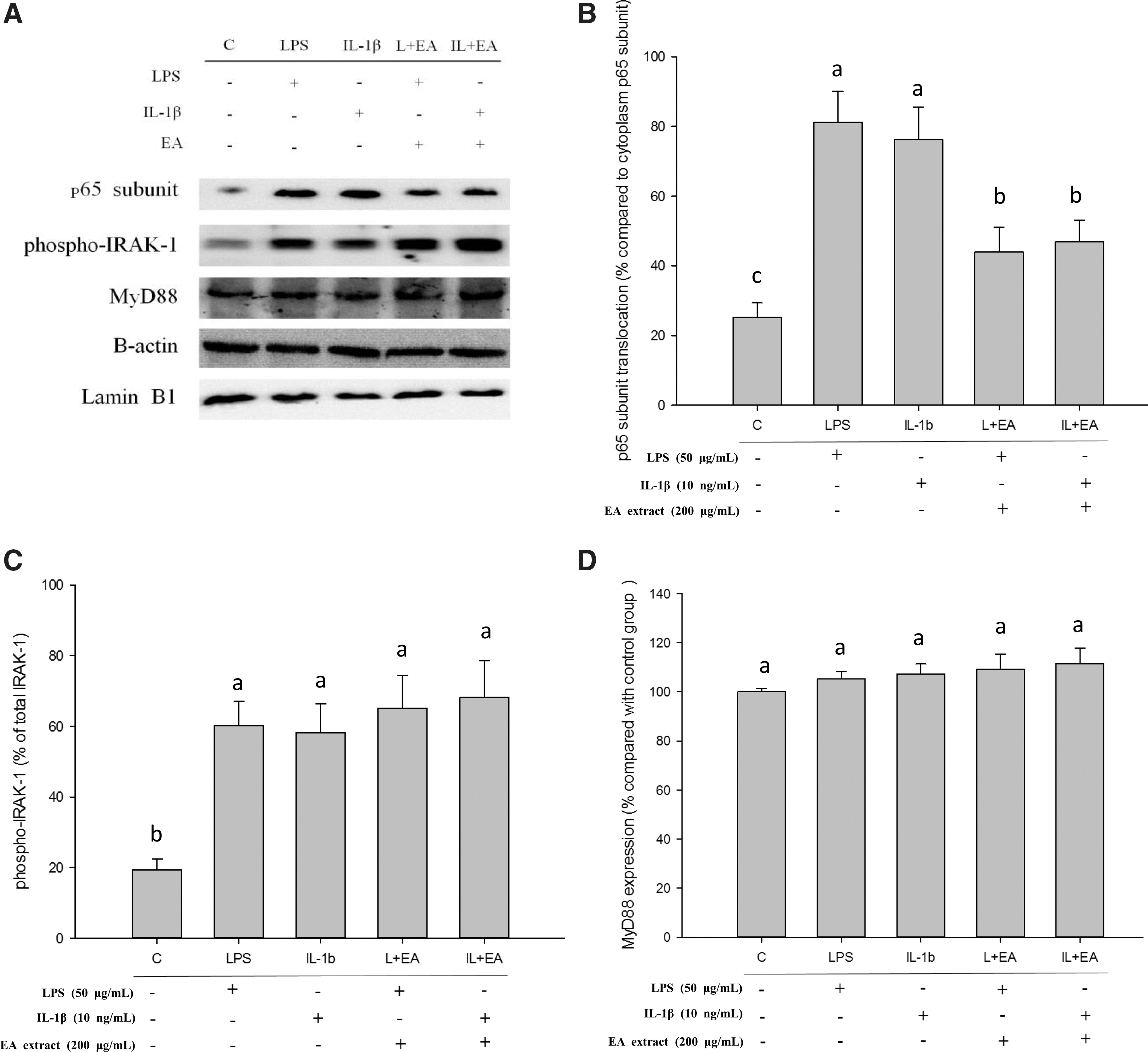
The changes
A previous study indicated that noni fruit contains other phenolic compounds such as scopoletin and rutin. 26 However, these two phenolics were not found to be abundantly present in this study, but quercetin was predominant (Table 1). A previous study also shows that quercetin levels of onion were increased after fermentation, which enhanced the antioxidation activities. 68 Therefore, the discrepancy was probably due to the biotransformation after noni fruit fermentation, which altered the composition of phenolic compounds. 69 –71 The combined forms of phenolics may hydrolyze to release the free form phenolic compounds during fermentation. 72 Thus, difference in phenolic compounds between nonfermented and fermented noni fruit is foreseeable. Moreover, most studies focus on fermented noni juice rather than fermented noni fruit, which may ignore other middle or low polar components of fermented noni fruit. 27,40 In this study, the whole fermented fruit was extracted and analyzed to identify all phenolic compounds. This was the first study to describe the anti-inflammatory effect of fermented noni fruit in colon epithelial cells. These results indicated that fermented noni fruit may contribute to the prevention of IBDs.
Summary
The ethanol extract of fermented noni fruit increased the number of beneficial bacteria. The ethyl acetate extract of fermented noni fruit relieved oxidative stress and suppressed inflammatory signals to achieve anti-inflammatory activities in colon epithelial cells. Quercetin was essential for anti-inflammatory actions, and other unknown components further enhanced the inhibition of COX-2 expression by quercetin.
This research clearly demonstrated that that fermented noni fruit may be beneficial for colon health. Clinical trials are needed to support the efficacy of fermented noni fruit for preventing or treating IBDs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.