
Abstract
Daucus carota L. ssp. carota (Apiacea, wild carrot, Queen Anne's lace) has been used in folk medicine throughout the world and recently was shown to possess anticancer and antioxidant activities. This study aims to determine the anticancer activity of the pentane fraction (F1) and the 1:1 pentane:diethyl ether fraction (F2) of the Daucus Carota oil extract (DCOE) against human colon adenocarcinoma cell lines (HT-29 and Caco-2). Treatment of cells with various concentrations of F1 or F2 fractions produced a dose-dependent inhibition of cell proliferation. Flow cytometric analysis indicated that both fractions induced sub-G1 phase accumulation and increased apoptotic cell death. Western blot revealed the activation of caspase-3, PARP cleavage, and a considerable increase in Bax and p53 levels, and a decrease in Bcl-2 level. Treatment of HT-29 cells with either fraction markedly decreased the levels of both phosphorylated Erk and Akt. Furthermore, the combined treatment of F1 or F2 with wortmannin showed no added inhibition of cell survival suggesting an effect of F1 or F2 through the phosphatidyl inositol 3-kinase (PI3K) pathway. This study proposes that DCOE fractions (F1 and F2) inhibit cell proliferation by inducing cell cycle arrest and apoptosis in HT-29 cells through the suppression of mitogen-activated protein kinase (MAPK)/Erk and PI3K/Akt pathways.
Introduction
C
Apoptosis is an important physiological process needed for tissue homeostasis and is induced by several artificial factors such as anticancer agents and irradiation. 10 Apoptosis is mediated by two pathways, the death receptor-dependent (extrinsic) pathway and the mitochondrial-dependent (intrinsic) pathway. 7 The mitochondrial pathway is mediated by the Bcl-2 family that includes the antiapoptotic protein Bcl-2 and the proapoptotic protein Bax. 11 As opposed to necrosis, apoptosis is very well marked by certain morphological changes such as membrane blebbing, DNA fragmentation, chromatin condensation, and formation of apoptotic bodies. 12 On the other hand, phosphatidyl inositol 3-kinase (PI3K) positively regulates cell growth and proliferation and leads to the inhibition of apoptosis. This is mainly through the activation of its downstream effector, the protein serine/threonine kinase Akt. 13,14 Another pathway involved in the regulation of the cell cycle and apoptosis is the mitogen-activated protein kinase (MAPK). The MAPK pathway consists of a family of serine/threonine kinases that leads to the expression of several transcription factors that positively induce cell cycle progression in response to mitogen and growth factor stimulation. 15 Accordingly, targeting either the PI3K or the MAPK pathway might constitute an important strategy in cancer treatment.
Wild carrot, Daucus carota L. ssp. carota, is a spiny-fruited herb that grows in moderate regions throughout the world.
16
The plant is commonly consumed as a salad in the Mediterranean diet or used as an additive in some recipes.
17
Its flower umbels are edible and usually french-fried, and its seed oil, which is commercially available, is used to flavor beverages and food products.
18
In Lebanon, people eat its young taproot cooked and use the plant in folk medicine for the treatment of cancer, gastric ulcer, diabetes, muscle pain, and to protect against hepatic diseases (
Recent studies in our laboratories, showed that DCOE possesses antioxidant, 26 antitumor, 26,27 anti-inflammatory, and antiulcer 28 activities. Preliminary data revealed that only two fractions of DCOE (pentane and 1:1 pentane:diethylether) showed promising anticancer effects. Therefore, this study evaluated the antiproliferative and proapoptotic effects of the above fractions against HT-29 and Caco-2 human cancer cell lines, and elucidated the mechanism of action involved.
Materials and Methods
Reagents
Dulbecco's modified Eagle's medium (DMEM) and dimethyl sulfoxide (DMSO) were purchased from Sigma (St. Louis, MO, USA). The Annexin V/PI apoptosis detection kit was purchased from Abcam (Cambridge, United Kingdom), and WST-1 reagent was purchased from Roche (Mannheim, Germany). All other chemicals used in this study were purchased from Sigma unless otherwise stated.
Sample collection and oil extraction
D. carota (Linnaeus) ssp. carota mature umbels were collected at the post flowering season between June and August from Byblos, Lebanon. The plant was identified according to the characteristics described in the “Handbook of Medicinal Herbs” 23 and confirmed by Dr. A. Houri, a Lebanese plant expert at the Lebanese American University. The extraction procedure was carried out according to the method described by Zeinab et al. Briefly, umbels were air dried in the shade and then cut into small pieces for oil extraction in methanol/acetone (1:1) for 72 h. The extract was then filtered and evaporated to dryness under reduced pressure. The residue was centrifuged and the oil was dried over anhydrous sodium sulfate. The final yield (3.47%) was stored in a closed amber bottle at 4°C until use.
DCOE fractionation
Thirty grams of DCOE were chromatographed on a silica gel column (35–70 mesh). The first fraction (F1) was eluted with pentane (100%), the second fraction (F2) with pentane:diethyl ether (50:50). Fractions were analyzed by thin layer chromatography using hexane:ethyl acetate (70:30) as the mobile phase and plates were stained with 2% anisaldehyde.
Gas chromatography and mass spectrometry analysis
The compositions of the F1 and F2 fractions were analyzed using gas chromatography and mass spectrometry (GC-MS) (HP6890 series; Hewlett Packard, Palo Alto, CA) fitted with a fused silica HP5-MS 5% phenyl methyl siloxane cap column (30 m×0.25 mm i.d., film thickness 0.25) and directly coupled to the MS. The carrier gas was helium with splitless injection and the flow rate of 1.2 mL/min was applied. The temperature program was as follows: 2.0 min at 70°C, from 70°C to 130°C at 8°C/min and hold for 5 min, from 130°C to 180°C at 2°C/min and hold for 10 min, from 180°C to 220°C at 15°C/min and hold for 2 min, and then from 220°C to 280°C at 15°C/min and hold for 22 min. Identification of the components was performed by comparing their mass spectra with the literature (NIST11 and W9). Percentage composition was computed from GC peak areas. 26
Cell lines and culture
Human colon cancer cell lines HT-29 and Caco-2 were purchased from American Type Culture Collection (ATCC, Rockville, MD, USA). Both cell lines were cultured in a humidified incubator at 37°C and 5% CO2 atmosphere in DMEM, supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin.
Cell proliferation assay
The proliferation of the HT-29 and Caco-2 cells was tested using the WST-1 assay. Cells were plated in 96-well plates at a concentration of 105 cells/mL for 24 h. Both cell lines were then treated with increasing concentrations (10×10−6, 25×10−6, 50×10−6, and 100×10−6 g/mL) of F1 and F2 fractions in 0.5% DMSO (v/v) for 48 h. At the end of the treatment period, WST-1 reagent was added to the cells and incubated in a humidified incubator at 37°C and 5% CO2 atmosphere for 3 h. The intensity of the produced formazan was quantified at 450 nm using a microplate ELISA reader. For wortmannin treatment, HT-29 cells were incubated with or without wortmannin (1 μM) for 1 h in a serum-free complete MEM before treating cells with 25 and 50 μg/mL of F1 and F2 for 48 h. 26
Apoptosis assay
The apoptotic effect of F1 and F2 on HT-29 cells was determined by Annexin V-FITC staining assay and measured on a C6 flow cytometer (BD Accuri Cytometers, Ann Arbor, MI, USA). The HT-29 cells (1×105 cells/mL) were treated with different concentrations (25 and 50 μg/mL) of both fractions and cultured in six-well plates for 48 h. Treated cells were harvested, washed with phosphate-buffered saline (PBS), and then centrifuged at 500 g for 5 min. Then, cell pellets were suspended in 500 μL using Annexin V-FITC/PI apoptosis detection kit at room temperature for 5 min in the dark. Annexin V-FITC binding was analyzed by flow cytometry (Ex=488 nm; Em=530 nm) FL1 channel for detecting Annexin V-FITC staining and FL3 channel for detecting propidium iodide (PI) staining. Annexin V-positive and PI-negative cells were scored as early apoptotic, and double-stained cells were considered as late apoptotic.
DAPI staining
HT-29 cells were seeded on 25 mm square glass cover slips. After treatment, cells were washed with PBS and fixed with 3.7% paraformaldehyde for 10 min at room temperature. Fixed cells were permeabilized with a 0.1% solution of Triton X-100 in PBS and then stained with 1 μg/mL of DAPI solution for 20 min. The cells were washed with PBS and observed with fluorescence microscope.
Cell cycle analysis
The effect of F1 and F2 fractions on cell cycle distribution was assessed by flow cytometry after staining the HT-29 cells with PI. Briefly, the HT-29 cells (1×105 cells/mL) were treated with different concentrations (25 and 50 μg/mL) of both fractions for 48 h. The treated cells were harvested, washed with PBS, and fixed with 70% ethanol on ice. Then cells were washed with cold PBS, suspended in 200 μL 1×PI+RNase staining solution, and incubated at 37°C in the dark for 30 min. Propidium Iodide Flow Cytometry Kit was used for cell cycle analysis. DNA content of the cells was measured using a C6 flow cytometer and the population of each phase was determined using C Flow Plus analysis software (BD Accuri Cytometers).
Western blot
HT-29 cells were treated with different concentrations of F1 and F2 (25 and 50 μg/mL) for 48 h. The adherent and nonadherent HT-29 cells were collected on ice, washed twice with PBS, lysed with lysis buffer, and centrifuged at 12,000 g for 10 min at 4°C. The cell lysate was heated at 100°C for 5 min, and the protein content was determined by the Bio-Rad protein assay (Bio-Rad, Hercules, CA, USA). The same amount of proteins was loaded onto a 10% SDS-PAGE. Proteins were then transferred to PVDF membrane (Pall Corporation, Ann Arbor, MI, USA) and blocked with 5% skim milk for 2 h. The membranes were probed with primary antibodies against actin, p53, Bcl-2, Bax, caspase-3, PARP, Akt, p-Akt, Erk, and p-Erk (Abcam) at 4°C overnight. Later, the primary antibodies were washed away with phosphate-buffered saline Tween (PBST) for 1 h and the membranes were treated with horseradish peroxidase-coupled secondary antibodies (Promega Corp., Madison, WI, USA) for 1 h, and then washed with PBST. Finally, detection of each protein was performed using the ECL kit (Abcam plc, Cambridge, United Kingdom).
Statistical analysis
Results were expressed as the mean±SEM of three experiments. Comparison of each group with the control was made by independent t-tests. A P value less than .05 was considered to be significant.
Results
GC-MS analysis of the F1 and F2 fractions with their respective components are shown in Table 1. The F1 fraction was predominantly composed of sesquiterpenes mainly α-humelene (28.2%), β-caryophyllene (17.2%), α-longipinene (7.18%), α-selinene (4.26%), γ-selinene (3.53%), β-selinene (2.86%), and β-himachalene (2.58%). The F2 fraction was markedly dominated by the sesquiterpene 2-himachalen-6-ol (61.4%) and noticeable amounts of three phenylpropanoids: elemicin (10.4%), (E)-methyl isoeugenol (11.8%), and methyl eugenol (2.22%).
HT-29 and Caco-2 human cancer cells were treated with different concentrations of F1 or F2 for 48 h (Fig. 1). The treatment with the extract appeared to be dose dependent with maximum inhibition of proliferation at higher concentrations (50 and 100 μg/mL). The IC50 values of F1 and F2 at 48 h were 22±1.12 and 17.5±0.73 μg/mL for HT-29 cells and 18.5±0.6 and 19±0.55 μg/mL for Caco-2, respectively.
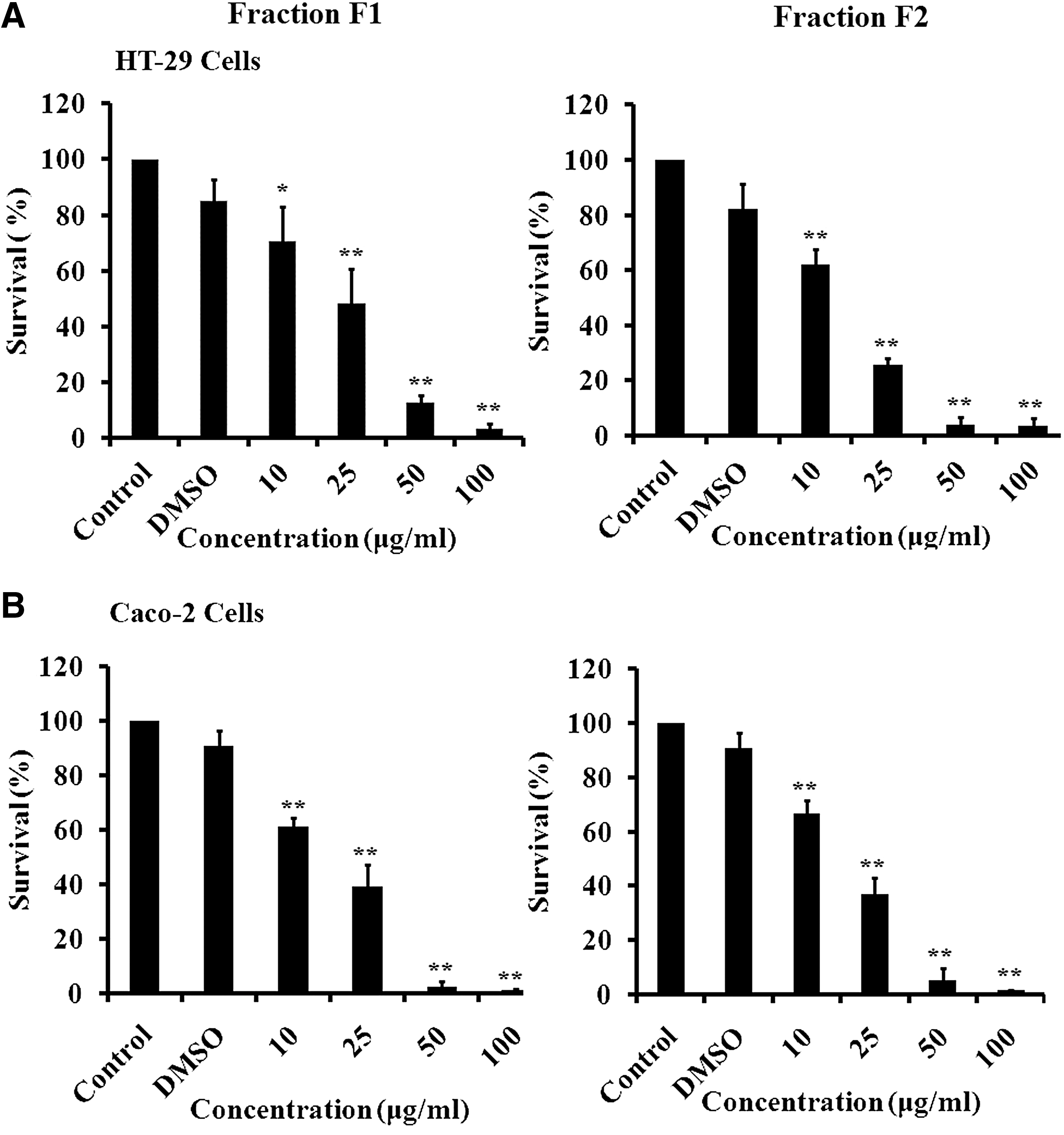
Antiproliferative effect of F1 and F2 fractions. HT-29
To determine whether F1 and F2 inhibitions of HT-29 cancer cell proliferation was due to cell cycle arrest or apoptosis, the cells were stained with PI after permeabilization to look at the cell cycle profile or with annexin and PI to look for apoptosis. Figure 2 shows that treatment of HT-29 with F1 or F2 (25 and 50 μg/mL) for 48 h increased the percentage of cells in the sub-G1 phase with hypodiploid nuclei, indicating that both fractions could cause DNA fragmentation. This increase was coupled with the decreased percentage of cells in the S and G2/M phases. On the other hand, Figure 3A and B revealed a significant increase in Annexin V/PI-positive cells indicating the induction of apoptosis by either fraction. Treatment with F1 or F2 mainly increased the late apoptotic cells. The apoptotic effect was also confirmed by chromatin condensation and formation of apoptotic bodies in treated cells (Fig. 4).
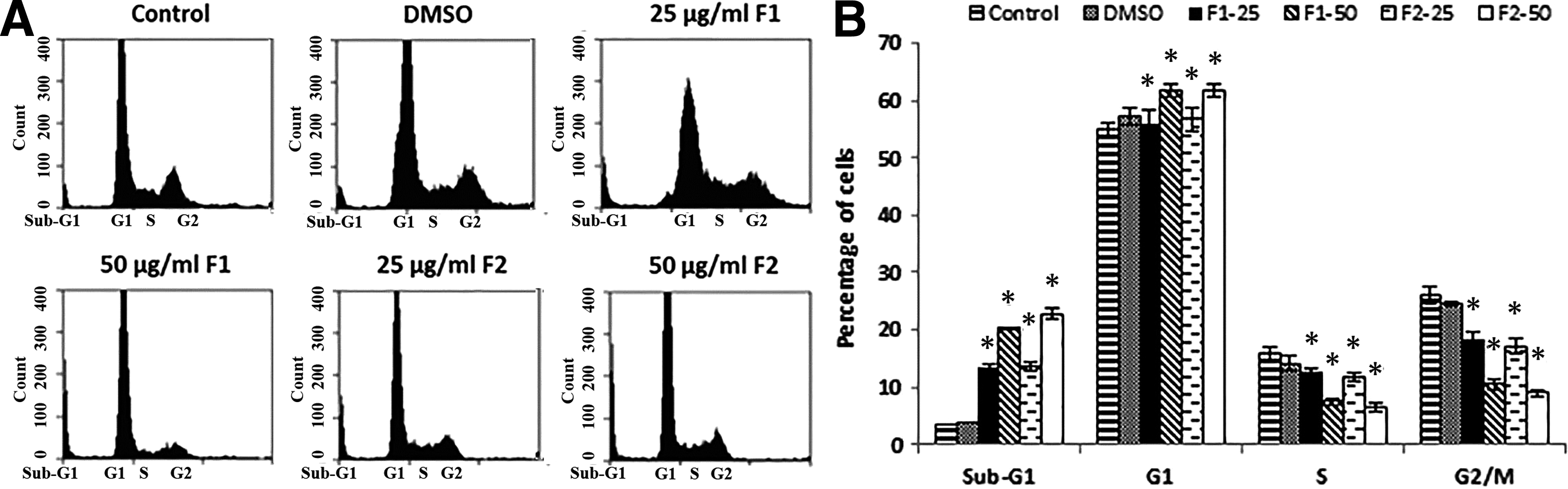
Effect of F1 and F2 fractions on the cell cycle distribution in HT-29. (
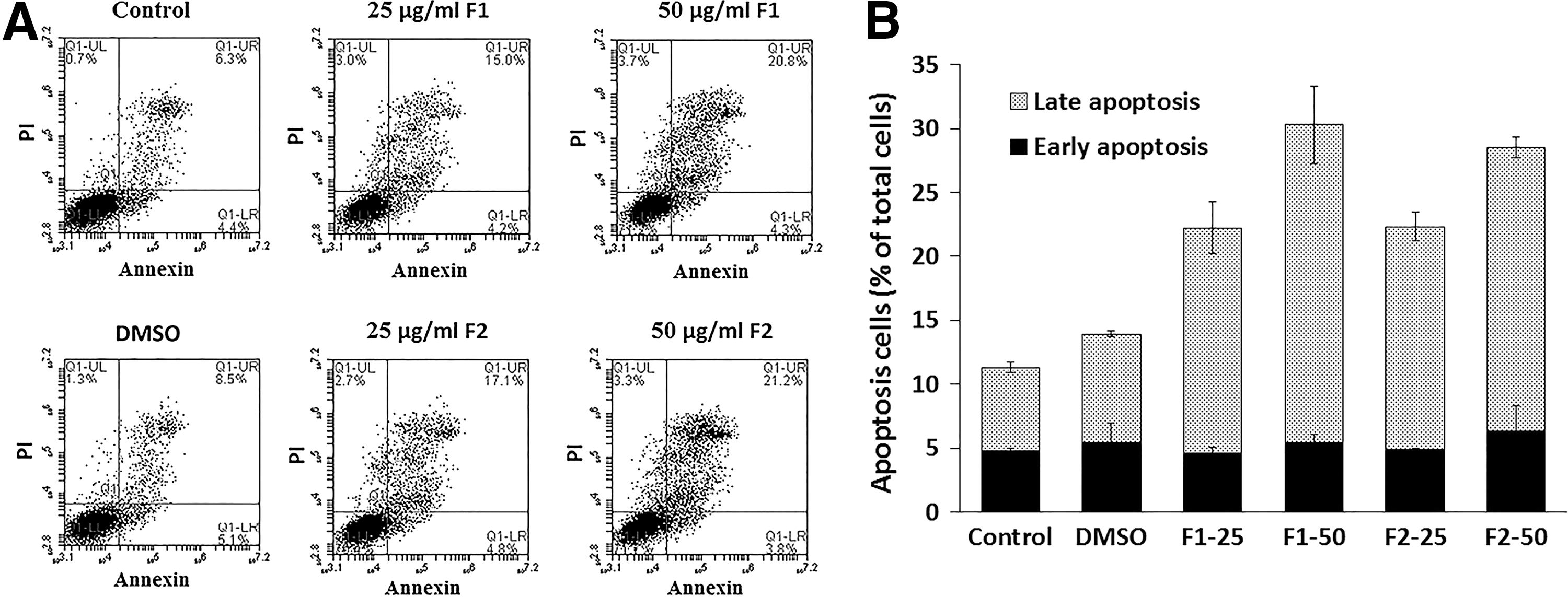
Apoptotic effect of F1 and F2 treatment on HT-29 cells.
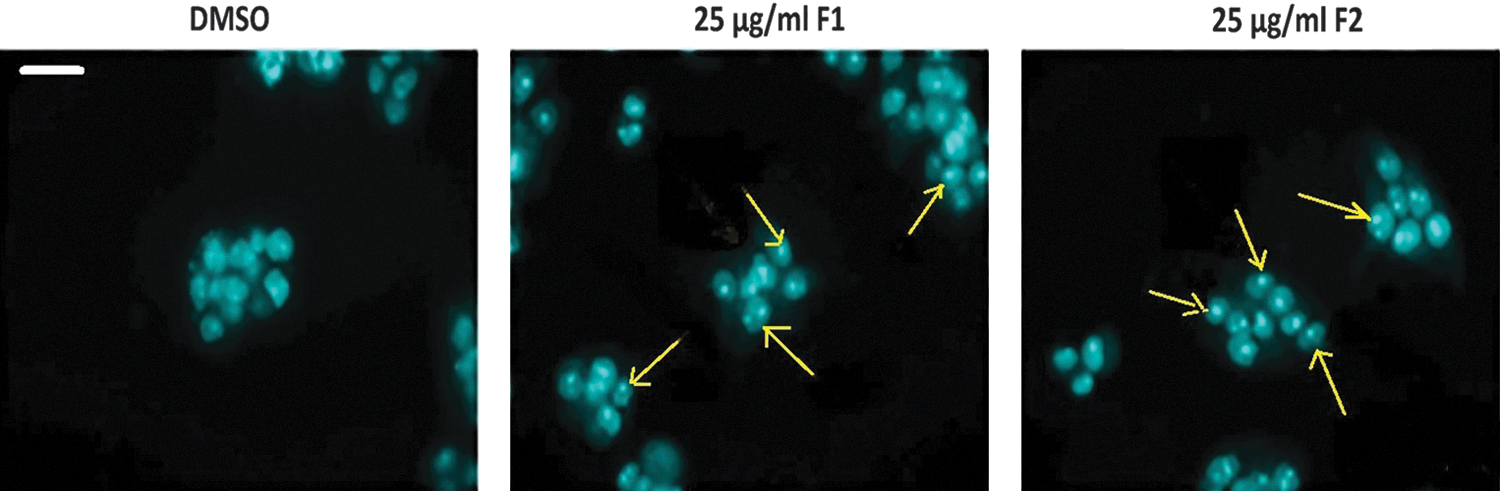
Apoptotic morphological changes in HT-29 cells treated with F1 or F2 fractions. Treatment of cells with 25 μg/mL of F1 or F2 fractions for 48 h. Nuclei were stained by DAPI and examined by fluorescence microscopy. Control cells treated with 0.5% DMSO. Arrows indicate apoptotic bodies of nuclear fragmentation. Scale bar is 10 μm. Color images available online at
To explore the mechanism of apoptosis induced by F1 and F2, the expression levels of various proapoptotic and antiapoptotic proteins were evaluated by western blotting. Treatment of HT-29 cells with F1 or F2 caused a decreased expression of procaspase-3 and an increase in the proteolytic cleavage of PARP (Fig. 5). The results of western blots also indicated that the expression of the antiapoptotic protein Bcl-2 was decreased, whereas the proapoptotic protein BAX was increased (Fig. 5). It was also observed that treatment of cells with either fraction induced an increase in p53 levels in a dose-dependent manner (Fig. 5).

Western blot analysis of apoptosis-related proteins in HT-29 cells.
The PI3K/Akt signaling pathway plays a central role in cellular proliferation and apoptosis. Thus, the effect of both fractions on the activity of Akt protein was studied by western blotting. Results showed that the level of phosphorylated Akt decreased after treatment with increasing concentration of either fraction (Fig. 6A, B). To evaluate the effect of F1 and F2 fractions on the expression of the MAPK pathway, the pErk levels in HT-29-treated cells were measured using western blotting. As shown in Figure 6A and B, the level of pErk decreased in a dose-dependent manner.

Western blot analysis of phosphatidyl inositol 3-kinase (PI3K)/Akt and mitogen-activated protein kinase (MAPK)/Erk pathways in HT-29 cells.
We further investigated whether the combined treatment of either fraction with wortmannin, a selective PI3K inhibitor, would have an additive effect on the inhibition of cell survival. WST results showed no significant decrease in cell survival of HT-29 cells treated with 25 μg/mL of F1 or F2 along with wortmannin (1 μM) compared to cells treated with fractions alone (Fig. 6C, D).
Discussion
Natural products play a major role in the treatment of different types of cancers and exert their effect through different mechanisms, including cell cycle arrest, induction of apoptosis, or inhibition of metastasis and tumor angiogenesis. 29,30 Recently, we reported that wild carrot oil possesses antioxidant and anticancer activities against breast and colon cancer cell lines. 26 This study demonstrates that the pentane fraction (F1) and the pentane/diether fraction (F2) of DCOE have significant dose-dependent antiproliferative effect against HT-29 and Caco-2 human colon cancer cell lines.
The anticancer activity of F1 and F2 might be attributed to some of the major components present in these fractions. For example, α-humulene and β-caryophyllene, which are major components of F1, have been reported to exhibit cytotoxic activity against several human cell lines. 31,32 Similarly, β-selinene, (E)-methylisoeugenol and elemicin, identified in F2, are also known to possesses antiproliferative activity and tumoricidial effects against several human cancer cell lines. 33 The antiproliferative activity of the F2 fraction may also be attributed to the presence of the major compound 2-himachalene-6-ol, which constitutes more than 60% of the bioactive compounds. To the best of our knowledge, this compound has not been reported in any of the wild carrot species around the world and there are no previous studies involving its anticancer activity. However, widdrol, a structural isomer of 2-himachalen-6-ol, extracted from Juniperus chinensis, exhibits significant anticancer activity against HT-29 colon adenocarcinoma cells by inhibiting their proliferation and inducing cell cycle G1 arrest. 34 In another study, widdrol induced G1 arrest through the induction of Chk2, p53 phosphorylation, and CDK inhibitor p21 expression in addition to inhibition of cyclin E, cyclin-dependent kinase-2, and retinoblastoma protein. 35
Current antineoplastic approaches target hallmark characteristics of cancer cells, including constitutive cell division and insensitivity to cell death-inducing factors. 36 Hence, many studies have been conducted to develop drugs that induce cell cycle arrest or apoptosis. 37 In this study, F1 and F2 were found to induce apoptosis in colon cancer cells, as verified by flow cytometry and immunostaining. The increase in the p53 protein levels along with the previously mentioned observations suggests that these fractions induce p53-dependent apoptosis. Flow cytometry reveals an increase in the percentage of cells in the sub-G1 phase, which is consistent with an increase in apoptosis. It also shows a decrease in cycling cells (cells in S and G2/M phases), which is indicative of cell cycle arrest. It is well established that activation of p53 can lead to cell cycle arrest or apoptosis. 38 Thus, the observed upregulation of p53 could explain the cell cycle arrest and increase in apoptosis in colon cancer cells. This increase in p53 could be due to the decrease in MAPK activity. In fact, recent reports showed that treatments inducing increased extracelluler signal regulated kinase (ERK) activation cause a simultaneous diminishing of p53 activity. 39 In addition, ERK inhibitors reversed this effect suggesting a direct effect of the MAPK pathway on p53. 39
In addition to p53, other apoptotic markers were examined to further confirm the increase in apoptosis. Treatment of HT-29 cells with both fractions causes a decrease in the expression of the antiapoptotic protein Bcl-2 and an increase in the proapoptotic protein BAX. The caspase family of proteins also participates in the regulation and execution of apoptosis. 40 Among the downstream effectors of apoptosis, caspases lead to the cleavage and inactivation of PARP, a protein that normally plays a crucial role in DNA repair. The present results reveal the proteolytic cleavage of caspase-3 and PARP upon treatment with F1 and F2, which is indicative of apoptosis.
Many anticancer natural products induce apoptosis through the inhibition of the MAPK signaling pathway. 41,42 ERK1/2 (MAPK) is a transcription factor that leads to the expression of several gene products that positively regulate cell survival. 43,44 Western blot results demonstrate that both F1 and F2 significantly decrease the level of phosphorylated Erk, which is associated with a decrease in cell survival and proliferation. In addition, the PI3K/Akt pathway is another important signaling pathway in controlling cell proliferation and apoptosis. 43 The serine/threonine protein kinase Akt, the main downstream effector of PI3K, is a key mediator of cell survival. 45 The present data also show that both F1 and F2 cause a decrease in Akt phosphorylation reflecting an inhibition of the PI3K pathway. Furthermore, the combination of F1 or F2 with a PI3K inhibitor shows no additional increase in the anticancer activity, indicating that the fractions and the PI3K inhibitor exert their effect through the same pathway. Collectively, our data show that the cell cycle arrest and the induction of apoptosis by either fraction may be mediated through the MAPK/Erk and the PI3K/Akt pathway. This is consistent with previous reports showing that caryophyllene oxide, a main component of F1, induces apoptosis through the suppression of the PI3K/Akt/mTOR pathway. 46
In conclusion, our data provide the first evidence that the DCOE fractions (F1 and F2) strongly inhibit cell proliferation and induce apoptosis in colon cancer cells. These effects were mainly mediated through the inhibition of the MAPK/Erk and PI3K/Akt pathways. Further studies are required to isolate and characterize the active component(s) in DCOE fractions, mainly the major compound 2-himachalen-6-ol present in the pentane:diethylether fraction (F2).
Footnotes
Author Disclosure Statement
No competing financial interests exist.