
Abstract
The proliferation and migration of vascular smooth muscle cells (VSMCs) in blood vessels are important in the pathogenesis of vascular disorders such as atherosclerosis and restenosis. Piperine, a major component of black pepper, has antioxidant, anticancer, and anti-inflammatory activity. However, the antiatherosclerotic effects of piperine have not been investigated. In this study, the effects of piperine on platelet-derived growth factor (PDGF)-BB-induced proliferation and migration of VSMCs were investigated. The antiproliferative effects of piperine were determined using MTT assays, cell counting, real-time polymerase chain reaction, and western blots. Our results showed that piperine significantly attenuated the proliferation of VSMCs by increasing the expression of p27kip1, regulating the mRNA expression of cell cycle enzymes (cyclin D, cyclin E, and PCNA), and decreasing the phosphorylation of extracellular signal-regulated kinase (ERK)1/2 in a noncytotoxic concentration-dependent manner (30–100 μM). Moreover, we examined the effects of piperine on the migration of PDGF-BB-stimulated VSMCs, as determined by the Boyden chamber assay, H2DCFDA staining, and western blots. Our results showed that 100 μM piperine decreased cell migration, the production of reactive oxygen species (ROS), and phosphorylation of the p38 mitogen-activated protein kinase (MAPK). Taken together, our results suggest that piperine inhibits PDGF-BB-induced proliferation and the migration of VSMCs by inducing cell cycle arrest and suppressing MAPK phosphorylation and ROS. These findings suggest that piperine may be beneficial for the treatment of vascular-related disorders and diseases.
Introduction
I
The proliferation and migration of VSMCs play central roles in vascular pathogenesis. 4,5 Arterial dysfunction is characterized by extensive cellular signaling related to abnormal proliferation and migration of VSMCs, which can be stimulated by numerous growth factors, including platelet-derived growth factor (PDGF). 6,7 Recently, studies have reported that PDGF-BB contributes to intracellular signaling through mitogen-activated protein kinases (MAPKs) in various cells. 8,9 VSMC migration and proliferation are also influenced by activated MAPKs, such as extracellular signal-regulated kinase (ERK)1/2 and p38 MAPK. 10,11 Moreover, ROS are involved in the proliferation and migration of PDGF-BB-stimulated VSMCs. 12 Inhibitors of MAPK signaling pathways have been shown to modulate neointimal formation in the balloon-injured arterial vessels of rats. 13 Cell cycle progression is regulated by enzymes such as cyclin E, cyclin A, cyclin B1, and proliferating cell nuclear antigen (PCNA); p27kip1 has been reported to inhibit the G0/G1 phases of the cell cycle by regulating some of these cell cycle-related enzymes. 14,15
Piperine is a nitrogen-rich substance responsible for the pungency of black pepper (Piper nigrum L.) and other Piper species. It mainly exhibits in vitro and in vivo anti-inflammatory, antioxidant, and antiadipogenic activity. 16 –19 However, the effects of piperine on the aberrant growth and migration of PDGF-BB-stimulated VSMCs through the scavenging of ROS have not been investigated.
In this study, we assessed whether piperine inhibits the cell growth and cell migration of PDGF-BB-stimulated VSMCs to determine whether it exhibited the antiatherosclerotic activity. We demonstrate that piperine regulated the cell cycle by increasing p27kip1 expression and inhibiting cell migration signaling through the ERK1/2 and p38 pathways in PDGF-BB-stimulated VSMCs.
Materials and Methods
Reagents and chemicals
Piperine (Fig. 1A) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were obtained from Sigma (St. Louis, MO, USA). Cell culture materials were purchased from Gibco BRL (Invitrogen, Grand Island, NY, USA). PDGF-BB was obtained from R&D Systems (Minneapolis, MN, USA). Total p38 (T-p38) and phospho-p38 (p-p38) antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). 4,6-Diamidino-2-phenylindole (DAPI) and antibodies against glyceraldehyde-3-phosphate dehydrogenase (GAPDH), ERK1/2 (T-ERK1/2), phospho-ERK1/2 (p-ERK1/2), and p27kip1 were obtained from Santa Cruz Biotechnology (Santa Cruz, MA, USA). The fluorescent dye 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA) was purchased from Invitrogen, and all other chemicals were obtained from Sigma.

Effects of piperine on the platelet-derived growth factor (PDGF)-BB-induced proliferation of vascular smooth muscle cells (VSMCs).
Animal care and cultivation of rat primary VSMCs
All animal experiments were approved and performed in accordance with the guidelines issued by the animal subjects committee of Dongguk University (No. 2014-07108). VSMCs were enzymatically isolated from male Sprague-Dawley rats (6 weeks old; weight, 190 g; n=4) as previously described. 20 VSMCs were grown in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin, and 200 mM glutamine at 37°C in a humidified 95% air/5% CO2 mixture (v/v).
VSMC viability test and cell proliferation assay
Cell viability was assessed using an MTT assay and direct cell counting as previously described. 21 Cell growth rates were determined by the MTT assay, with absorption read at 570 nm. Cell morphologies and numbers were observed and recorded using an inverted microscope (IX71; Olympus, Tokyo, Japan) equipped with a CCD camera.
RNA extraction and real-time quantitative reverse transcription–polymerase chain reaction
Total RNA was isolated from VSMCs using the TRI-reagent (MRC, Cincinnati, OH, USA) according to the manufacturer's instructions. Briefly, cDNA was synthesized using 1 μg of total RNA and the Improm II reverse transcription system with oligodeoxythymidine (oligo-dT) primers (Promega, Madison, WI, USA). mRNA expression was quantified using the Fast Start Essential DNA Green Master mix (Roche Diagnostics, Mannheim, Germany) and a LightCycler 96 (Roche Diagnostics). The following primers were used for SYBR Green real-time reverse transcription–polymerase chain reaction (RT-PCR): cyclin E, sense primer, 5′-ATG TCC AAG TGG CCT ACG TC-3′ and antisense primer, 5′-TCT GCA TCA ACT CCA ACG AG-3′; cyclin A, sense primer, 5′-GCT TTT AGT GCC GCT GTC TC-3′ and antisense primer, 5′-AGT GAT GTC TGG CTG CCT CT-3′; PCNA, sense primer, 5′-TCA CAA AAG CCA CTC CAC TG-3′ and antisense primer, 5′-CAT CTC AGA AGC GAT CGT CA-3′; cyclin B1, sense primer, 5′-CTG CTC AGG AGA CCA TGT A-3′ and antisense primer, 5′-CTA CGG AGG AAG TGC AGA GG-3′; and GAPDH, sense primer, 5′-TGG AGT CTA CTG GCG TCT T-3′ and antisense primer, 5′-TGT CAT ATT TCT CGT GGT TCA-3′. The following real-time quantitative RT-PCR conditions were used for cyclin E, cyclin A, PCNA, cyclin B1, and GAPDH amplification: initial denaturation at 95°C for 10 min, and 40 cycles of denaturation at 94°C for 10 sec, annealing at 60°C for 10 sec, and extension at 72°C for 30 sec. Transcripts were quantified using LightCycler 96 software, and all assays were performed in triplicate.
Western blot analysis
Equal amounts of cell lysate (20 μg of protein) were separated using 12% acrylamide gels and electroblotted onto polyvinylidenedifluoride (PVDF) membranes (PALL, New Port Richey, FL, USA). Membranes were blocked with 5% bovine serum albumin in Tris-buffered saline (TBS) at room temperature for 1 h and then incubated overnight at 4°C with p-ERK1/2 and p-p38 antibodies, as well as T-ERK1/2 and T-p38 antibodies (all at 1:1000 dilution). The membranes were then washed with TBS containing 1% Tween-20 and incubated with the IgG secondary antibody conjugated to horseradish peroxidase. Protein expression levels were analyzed using an enhanced chemiluminescence plus kit (Amersham Pharmacia Biotech, Buckinghamshire, United Kingdom). Developed protein bands were visualized and quantified using ImageJ software.
Scratch wound healing assay
The scratch wound healing assay was conducted as previously described. 21 VSMCs (2×104 cells) were seeded in six-well plates and incubated for 24 h. After this, the cells were incubated with a serum-deficient medium for 24 h and then were either left untreated or treated with mitomycin C (MCC) for 1 h. Subsequently, cells in all wells were stimulated for 24 h with PDGF-BB in the presence of different concentrations of piperine (0, 10, 30, and 100 μM). A transverse scratch wound on VSMC monolayers was made using a sterilized 200-μL tip. The transverse scratch wounds were then examined and compared to those of untreated VSMCs. Cell numbers in the scratch area were counted and recorded using an inverted microscope (IX71; Olympus), and the data were analyzed using ImageJ software.
Detection of intracellular ROS
Intracellular ROS were assessed using the H2DCFDA (a fluorescent dye) assay, as recommended by the manufacturer (Invitrogen). Briefly, VSMCs were pretreated in the presence or absence of 100 μM piperine for 30 min and then stimulated with PDGF-BB (10 ng/mL) for 10 min at 37°C. VSMCs were then washed twice with phosphate-buffered saline and then stained with 5 μM H2DCFDA for 30 min at 37°C. Finally, the cells were fixed in 4% paraformaldehyde for 30 min at room temperature and then were immediately analyzed under a fluorescence microscope (BX50; Olympus). Nuclei were counterstained with DAPI, and the proportion of H2DCFDA-positive cells to total cells (stained by DAPI) was determined in three fields.
Cell migration assay
The PDGF-BB-mediated VSMC migration assay was performed using a Boyden chamber, as previously described, with slight modifications. 20 The lower chamber was loaded in the presence or absence of 100 μM piperine with PDGF-BB. The upper chamber was filled with 50 μL of VSMCs (1×106 cells/mL), and a membrane coated with type I collagen was placed between the lower and upper chambers. After incubation for 90 min, nonmigrating VSMCs were removed from the upper chamber according to the manufacturer's instructions (Corning, Tewksbury, MA, USA). Migrant cells were fixed in 4% paraformaldehyde and stained with 0.5% Toluidine blue. Migrating VSMCs were observed and quantified using an optical microscope (CKX41; Olympus) equipped with a CCD digital camera (DP71; Olympus).
Statistical analysis
The results are expressed as the mean±standard error of at least three independent experiments (n≥3). The statistical significances of the difference between the two experimental groups were determined by using Student's t-test. A one-way analysis of variance with Tukey's post test was used for multiple comparisons (GraphPad Prism ver. 4.00 for Windows, San Diego, CA, USA), and P-values<.05 were considered statistically significant.
Results
Effects of piperine on PDGF-BB-induced proliferation of VSMCs
The MTT assay was used to determine the inhibitory effects of piperine on PDGF-BB-induced VSMC proliferation. As shown in Figure 1, the proliferation rate of untreated cells did not change, whereas that of cells treated with PDGF-BB (10 ng/mL) significantly increased by 153.0%. However, cotreatment with 10, 30, and 100 μM piperine decreased the proliferation of PDGF-BB-stimulated VSMCs by 44.3%, 57.6%, and 138.0%, respectively. Furthermore, similar dose-dependent inhibitory effects of piperine were observed by cell counting. The treatment with PDGF-BB increased cell counts to 3.58×104 cells/mL, whereas the number of cells in cultures that were cotreated with 10, 30, and 100 μM piperine decreased to 2.58×104, 1.33×104, and 0.91×104 cells/mL, respectively (Fig. 1C). Although piperine reduced the proliferation of PDGF-BB-stimulated VSMCs, the cell morphologies remained unchanged under all conditions (Fig. 1D).
Effects of piperine on the PDGF-BB-stimulated cell cycle in VSMCs
We investigated whether piperine regulates the cell cycle in PDGF-BB-stimulated VSMCs. To determine if piperine regulates the cell cycle, we quantified the mRNA levels of the cell cycle factors cyclin E, cyclin A, PCNA, and cyclin B1 by real-time PCR using specific primers. As shown in Figure 2A, cells treated with PDGF-BB (10 ng/mL) showed the following increases in mRNA expression: cyclin E, 56.4±6.5-fold; cyclin A, 1.14±0.1-fold; PCNA, 18.4±2.3-fold; and cyclin B1, 36.4±4.6-fold to the expression of cells cultured in the absence of PDGF-BB and piperine. In contrast, low concentrations of piperine (10 μM) resulted in a lower increase in mRNA expression as follows: cyclin E, 8.3±2.2-fold; cyclin A, 5.19±0.1-fold; PCNA, 8.53±1.83-fold; and cyclin B1, 25.6±3.8-fold to the expression in cells cultured in the absence of PDGF-BB and piperine. When cells were cotreated with 100 μM piperine, an even lower increase in the mRNA expression of cell cycle enzymes was observed as follows: cyclin E, 0.7±0.0-fold; cyclin A, 1.15±0.0-fold; PCNA, 6.5±0.1-fold; and cyclin B1, 1.8±0.0-fold to the expression in cells cultured in the absence of PDGF-BB and piperine (Fig. 2A). In particular, western blot analysis showed that the expression of p27kip1 in PDGF-BB-stimulated VSMCs increased by 37.3%±2.0%, 53.6%±3.6%, and 63.3%±3.7% after treatment with 10, 30, or 100 μM piperine, respectively, whereas expression in the PDGF-BB (10 ng/mL)-treated group significantly decreased by 48.0%±6.4%, relative to that in the untreated group (Fig. 2B).
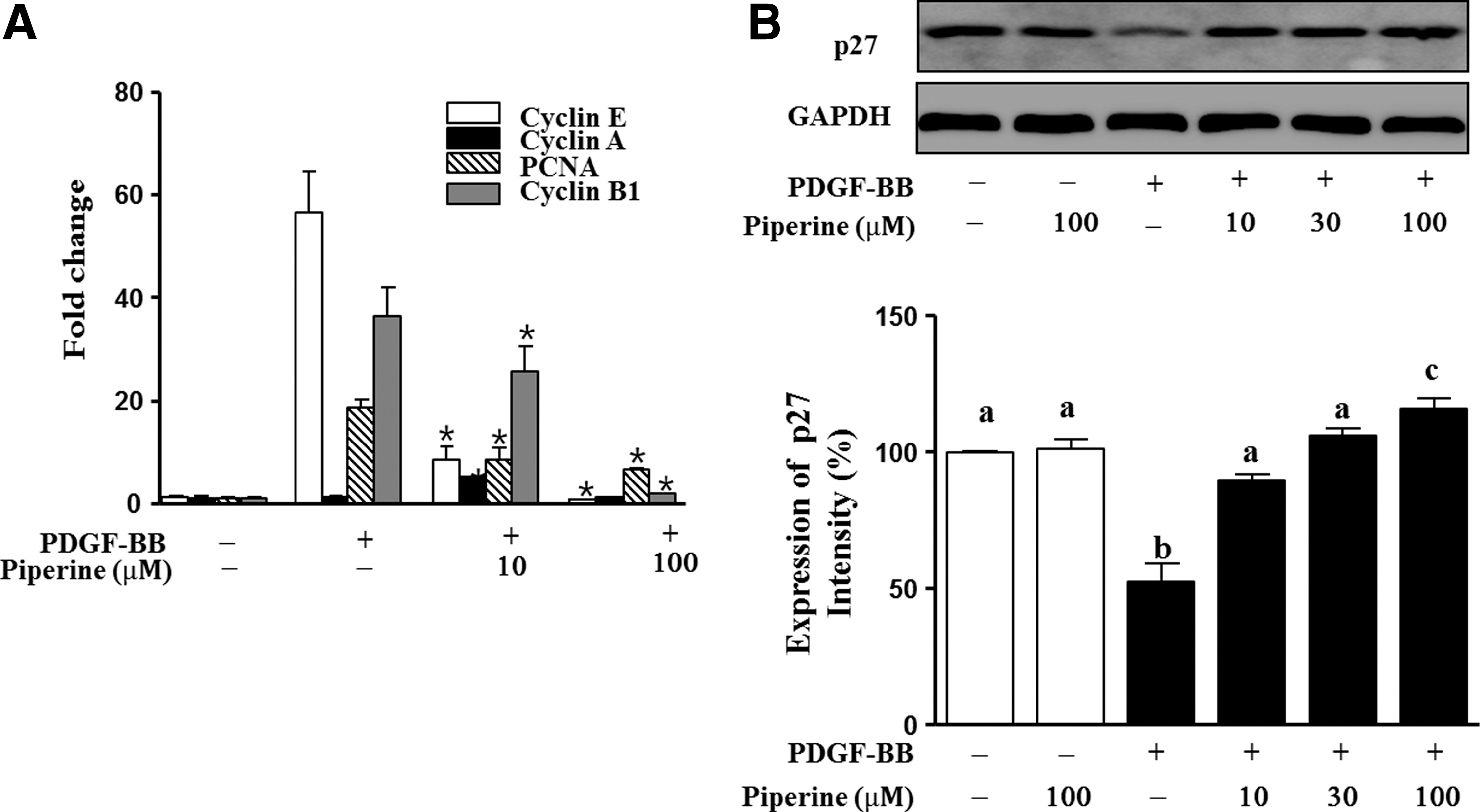
Effects of piperine on the expression of cell cycle enzymes in PDGF-BB-stimulated VSMCs.
Piperine inhibited scratch wound healing in PDGF-BB-induced VSMCs
A scratch wound healing assay was used to determine whether piperine could inhibit the migration or proliferation of PDGF-BB-stimulated VSMCs. As shown in Figure 3, the cell numbers in the wound healing areas of cultures treated by piperine decreased dose dependently in the absence of MCC; 10, 30, and 100 μM piperine-treated cultures had 7.1±0.8, 15.7±1.4, and 21.75±1.5 cells per field less than that in PDGF-BB-treated cultures, respectively (Fig. 3A). When the cells were pretreated with MCC for 1 h, the cell numbers in the wound healing area were reduced only in cultures treated with 100 μM piperine (Fig. 3B).
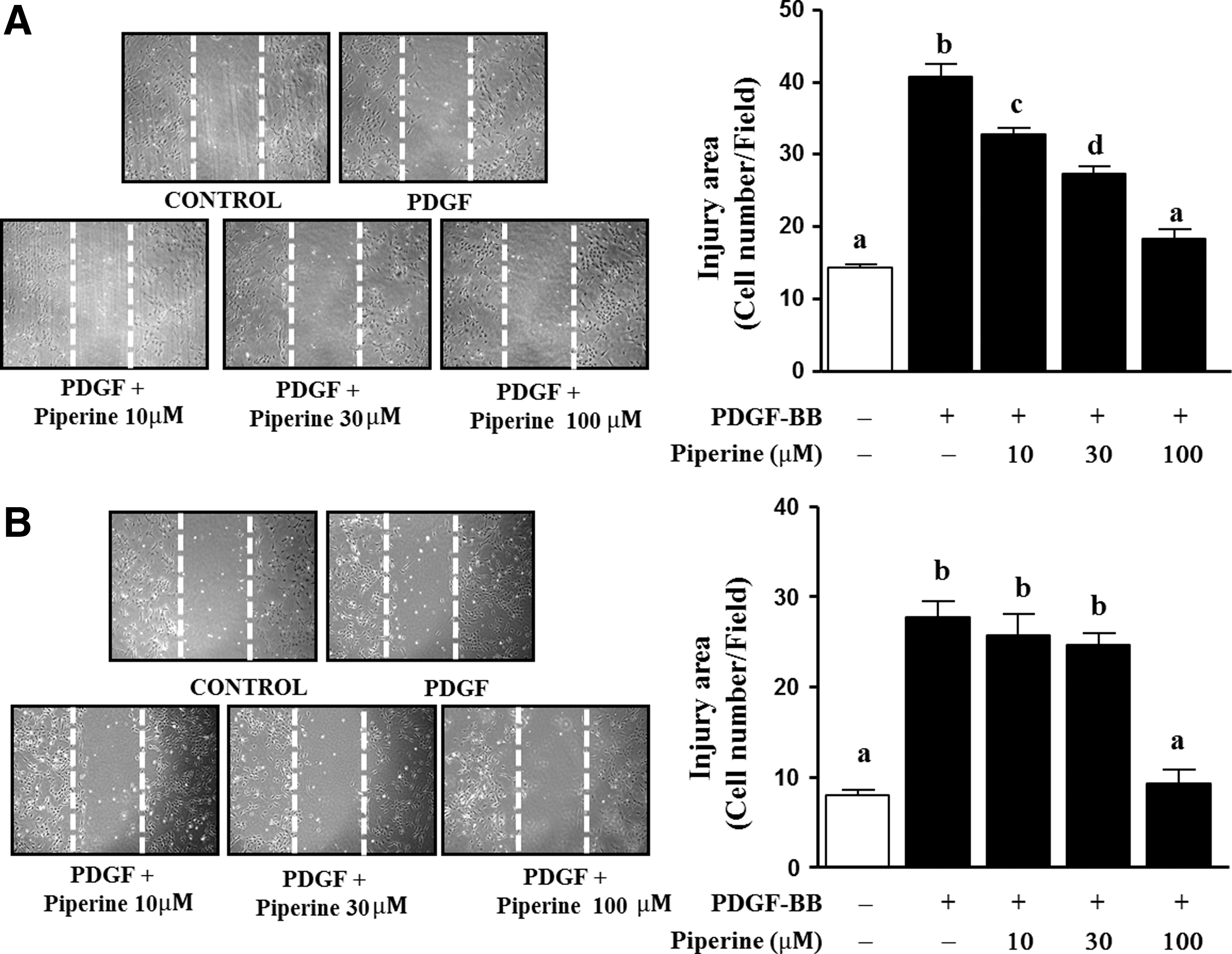
Effects of piperine on scratch wound healing in PDGF-BB-stimulated VSMCs. VSMCs were cotreated with piperine (10, 30, or 100 μM) and PDGF-BB (10 ng/mL) for 24 h. The scratch wound healing assay was performed, as described in the Materials and Methods section. The mitomycin C untreated
Piperine decreased the phosphorylation of p38 and ERK1/2 in PDGF-BB-stimulated VMSCs
Using western blot analysis, we investigated whether piperine could inhibit phosphorylation of p38 and ERK1/2 in PDGF-BB-induced VSMCs (Fig. 4A). As shown in Figure 4, after treatment with 10, 30, and 100 μM piperine, ERK1/2 phosphorylation decreased in a dose-dependent manner by 13.5%±2.0%, 15.6%±4.7%, and 72.6%±5.1%, respectively, relative of that in cells treated with PDGF-BB (Fig. 4B). In the case of p38, phosphorylation levels were similar following cotreatment with 10 or 30 μM of piperine, whereas with 100 μM piperine, its phosphorylation was significantly decreased by 59%±2.0% that of cells treated with PDGF-BB only (Fig. 4C).
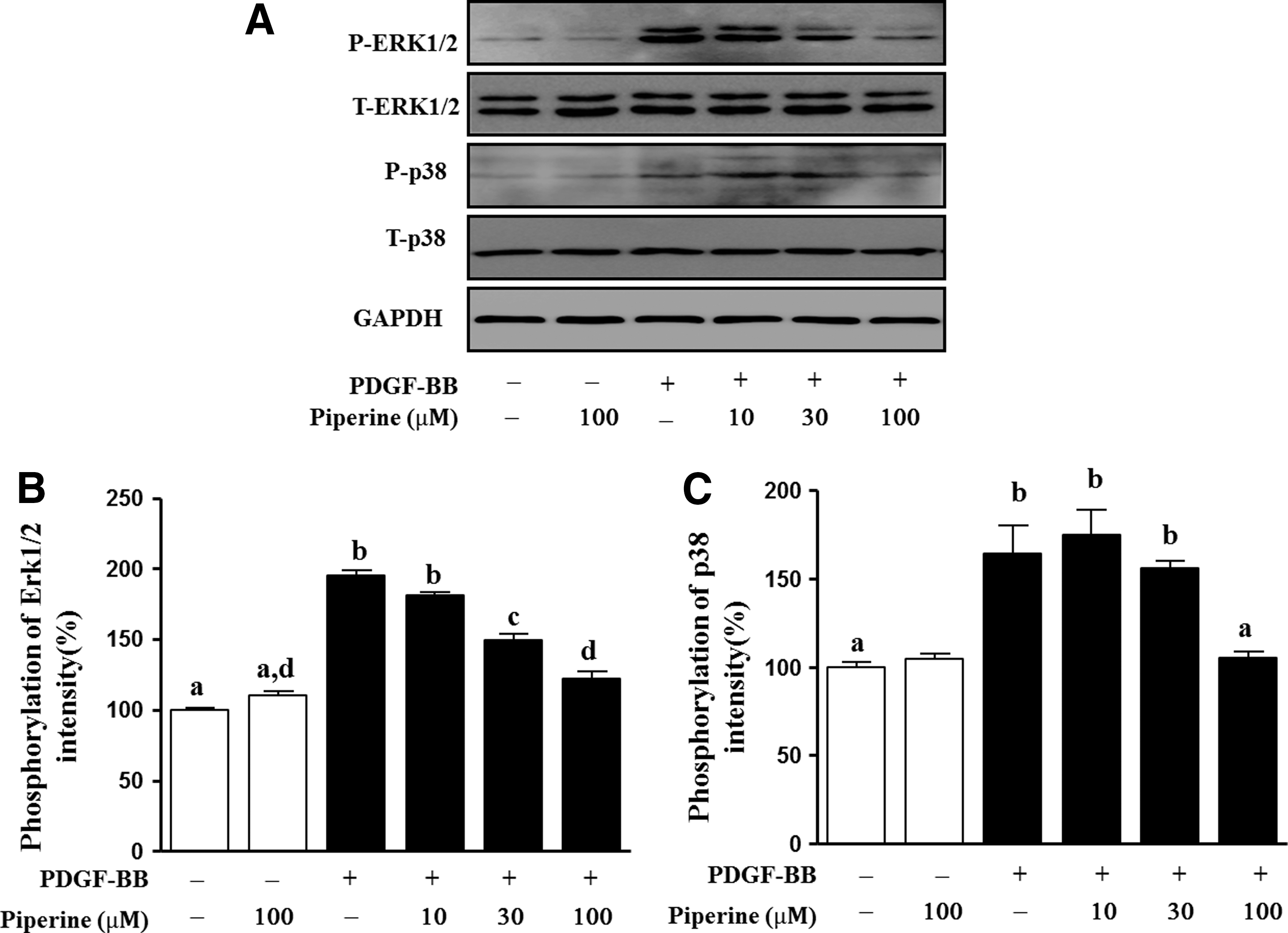
Effects of piperine on the PDGF-BB-stimulated phosphorylation of extracellular signal-regulated kinase (ERK)1/2 and p38 in VSMCs. VSMCs were either pretreated with piperine (10, 30, or 100 μM) for 1 h or were untreated, before stimulation with 10 ng/mL PDGF-BB for 15 min.
Effect of piperine on ROS generation in PDGF-BB-stimulated VSMCs
We also examined the inhibitory effects of piperine on ROS generation in PDGF-BB-stimulated VSMCs by using H2DCFDA staining. VSMCs were pretreated with piperine for 1 h and then treated with PDGF-BB (10 ng/mL) for 15 min. As shown in Figure 5A, ROS production increased to 86.9%±3.1% when cells were treated with PDGF-BB, but this increase was inhibited by pretreatment with 100 μM piperine, and ROS levels were restored to approximately baseline values of 103.0%±12.4% found in untreated cells. Lower concentrations of piperine (10 and 30 μM) had no effect on ROS generation (data not shown).
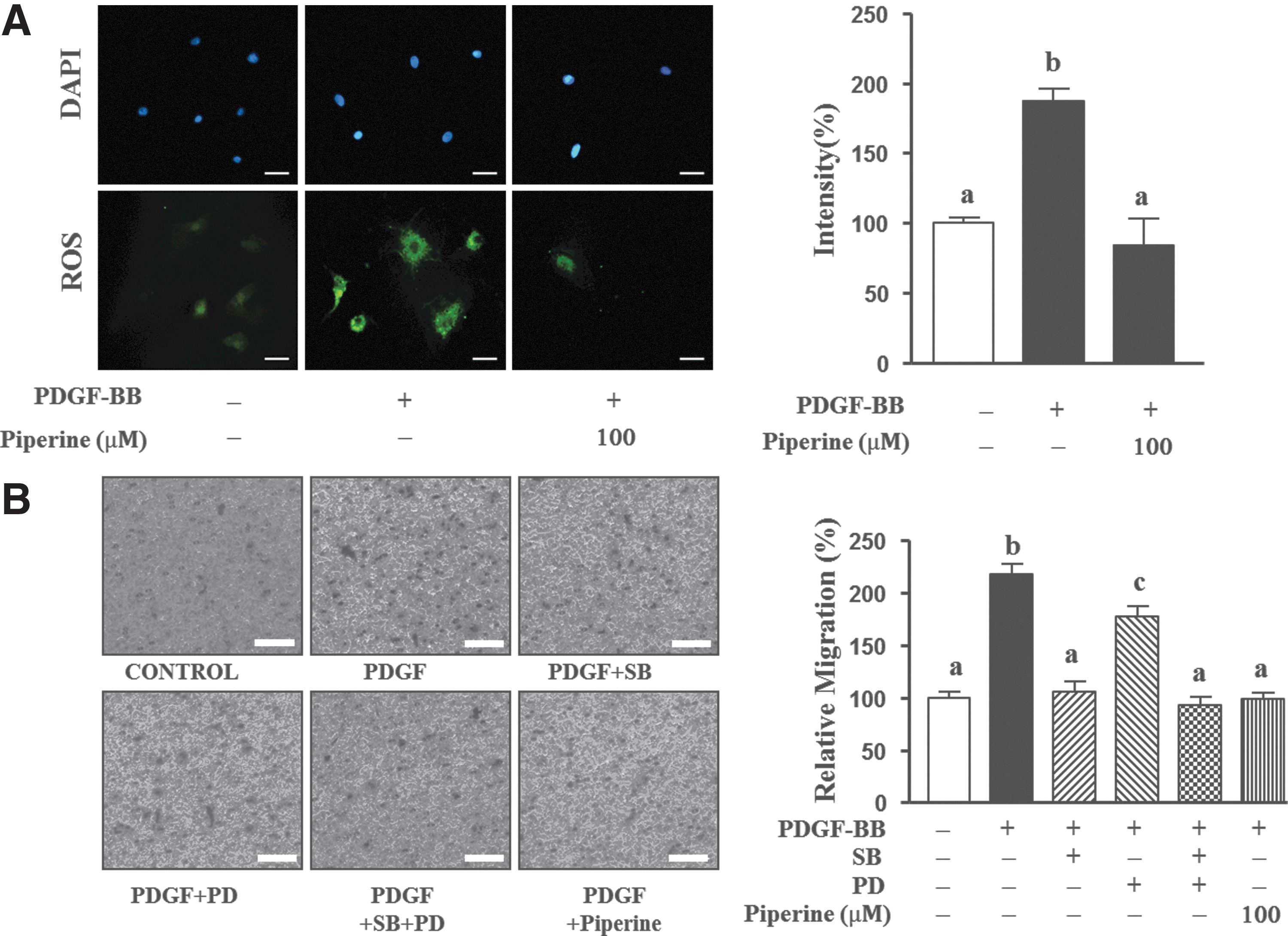
Effects of piperine on PDGF-BB-mediated intracellular reactive oxygen species (ROS) generation and VSMC migration.
Effects of piperine on the PDGF-BB-mediated migration of VSMCs
To confirm the observed inhibitory effects of piperine on VSMC migration, we performed a Boyden chamber assay. As shown in Figure 5B, migration of VSMCs stimulated by PDGF-BB for 90 min increased by 117.6%±11.4% of that of cells not treated with PDGF-BB, whereas this migration was attenuated by 111.1%±6.4% of that of the untreated control in the 100 μM piperine-treated cells. PDGF-induced migration of VSMCs reduced by 111.8%±12.4% in 30 μM SB20358-treated cells, by 39.7%±22.2% in PD98059-treated cells, and by 125.0%±20.8% in the cells cotreated with SB30358 and PD98059.
Discussion
Vascular disease insidiously progresses throughout life, is considered incurable, and is associated with high mortality. 22 Statins, HMG-CoA reductase inhibitors, are cholesterol-lowering drugs that have been used to treat atherosclerosis, as the metabolic dysfunction associated with a high-fat diet is involved in the development of atherosclerosis. However, the first stage of restenosis requires treatment with an antiplatelet drug because statins are ineffective in the progressed hyperplasia of VSMCs from the lesion of blood vessels. 23 Thus, dietary approaches are an important component of the strategies used to prevent vascular disorders and may be useful for improving patient survival and for slowing the progress of restenosis. Black pepper (P. nigrum L.) is a commonly used spice and is used in traditional medicine in Asian countries. 16,24 Furthermore, it has been used in Korean traditional medicine for the treatment of digestive disorders such as gastric ulcers and indigestion. 25 Piperine, the major component of black pepper, has well-known metabolic effects in vitro and in vivo. 17 –19 Although piperine exhibits a wide range of bioactivities in diverse cells, its effects on PDGF-BB-induced proliferation and migration of VSMCs have not been investigated. In this study, we demonstrated that piperine inhibited the proliferation, p27kip1 activation, and expression of representative cell cycle enzymes, such as cyclin E, cyclin A, cyclin B1, and PCNA, in PDGF-BB-stimulated VSMCs. Moreover, we demonstrated that the inhibitory effects of piperine on PDGF-BB-stimulated VSMC migration are exerted through downregulation of the phosphorylation of ERK1/2 and p38.
Abnormal proliferation of PDGF-BB-stimulated VSMCs increases the risk of vascular disorders, and it is critical for the progression of restenosis and atherosclerosis. 5 In the present study, we first examined the effects of piperine on PDGF-BB-induced proliferation in VSMCs. Based on MTT assays and cell counting, treatment of cells with piperine at a maximum concentration of 100 μM for 24 h did not result in any cytotoxicity. Notably, the proliferation of PDGF-BB-stimulated VSMCs was significantly reduced by piperine in a dose-dependent manner.
We also determined whether piperine could induce cell cycle arrest in PDGF-BB-stimulated VSMCs. Cell cycle progression is dependent on enzymes such as cyclins. 14 We observed that piperine significantly decreased the expression of key cell cycle enzymes such as cyclin E, cyclin A, PCNA, and cyclin B1.
In general, p27kip1, a cyclin-dependent kinase inhibitor, plays an important role in arresting the cell cycle. 15 Expression of p27kip1 has been reported to negatively regulate the proliferation rate of PDGF-BB-stimulated VSMCs. 26 We also observed that PDGF-BB decreased p27kip1 expression in VSMCs, whereas cotreatment with piperine (10–100 μM) significantly increased p27kip1 expression in a dose-dependent manner. These results suggest that piperine exerts its antiproliferative effects by regulating the expression of p27kip1, which inhibits the proliferation of PDGF-BB-stimulated VSMCs by decreasing the expression of cyclins such as cyclin E, cyclin A, cyclin B1, and PCNA.
Furthermore, the antimigratory effects of piperine were determined using a scratch wound healing assay and the Boyden chamber assay. The migration of PDGF-BB-stimulated VSMCs was significantly reduced by 100 μM piperine. These results suggested that piperine exerted antiatherosclerotic effects by regulating the migration of VSMCs.
MAPKs are involved in cellular responses required for cell survival, including cell migration, proliferation, and differentiation. 8,27 In VSMCs, these responses are initiated when PDGF-BB induces the activation of downstream signaling molecules, such as by phosphorylation of ERK1/2 and p38. 7 Our data show that piperine reduces the phosphorylation of ERK1/2 in PDGF-BB-stimulated VSMCs in a dose-dependent manner and upregulates p27kip1 expression. However, only a high concentration of piperine (100 μM) reduced the levels of ROS, leading to the inhibition of p38 phosphorylation. These results raised a question about how the phosphorylation of p38 is inhibited in PDGF-BB-stimulated VSMCs. The Boyden chamber assay also revealed that 100 μM piperine reduced the migration of and ROS generation in PDGF-BB-stimulated VSMCs. Interestingly, similar antimigratory effects were found in PDGF-BB-induced VSMCs treated with 100 μM piperine, SB250358 (p38 inhibitor), or both SB250358 and PD98059 (ERK1/2 inhibitor). These results indicate that decreased ROS generation and the resultant inhibition of the migration-related pathways activated by ROS are related to the decrease in the phosphorylation of p38 MAPK. The phosphorylation of p38 is known to be stimulated by ROS, which are involved in modulating the migration of VSMCs. 20 According to a number of studies, antioxidants exert their antiatherosclerotic effects by scavenging ROS, which in turn inhibits the ROS-induced proliferation and abnormal migration of VSMCs. Previous studies have reported that the antioxidant effects of curcumin and resveratrol lead to significant reduction in cell migration and proliferation. 28,29
In conclusion, our results demonstrate that piperine concentration dependently inhibits cell proliferation by inhibiting ERK1/2 phosphorylation and limiting the synthesis of specific cell cycle enzymes, including cyclin D, cyclin B1, cyclin E, and PCNA (Fig. 6A). We also speculate that at a high dose, piperine plays another important role, inhibition of ROS generation, which leads to suppression of the migration and abnormal proliferation of PDGF-BB-stimulated VSMCs. Therefore, our findings suggest that piperine, a component of black pepper, can upregulate the expression of p27kip1, which in turn prevents the abnormal proliferation of VSMCs by inducing cell cycle arrest. Furthermore, we have shown here that piperine suppresses the migration of PDGF-BB-stimulated VSMCs through ROS scavenging. Based on ex vivo data, black pepper can be considered a nutraceutical candidate for the treatment of restenosis (Fig. 6B). However, based on the present data, we cannot specify the exact amount of black pepper required to maintain a sufficient piperine level in the circulating blood. Because our data were obtained in in vitro and ex vivo analyses, further in vivo experiments are required to confirm these results.
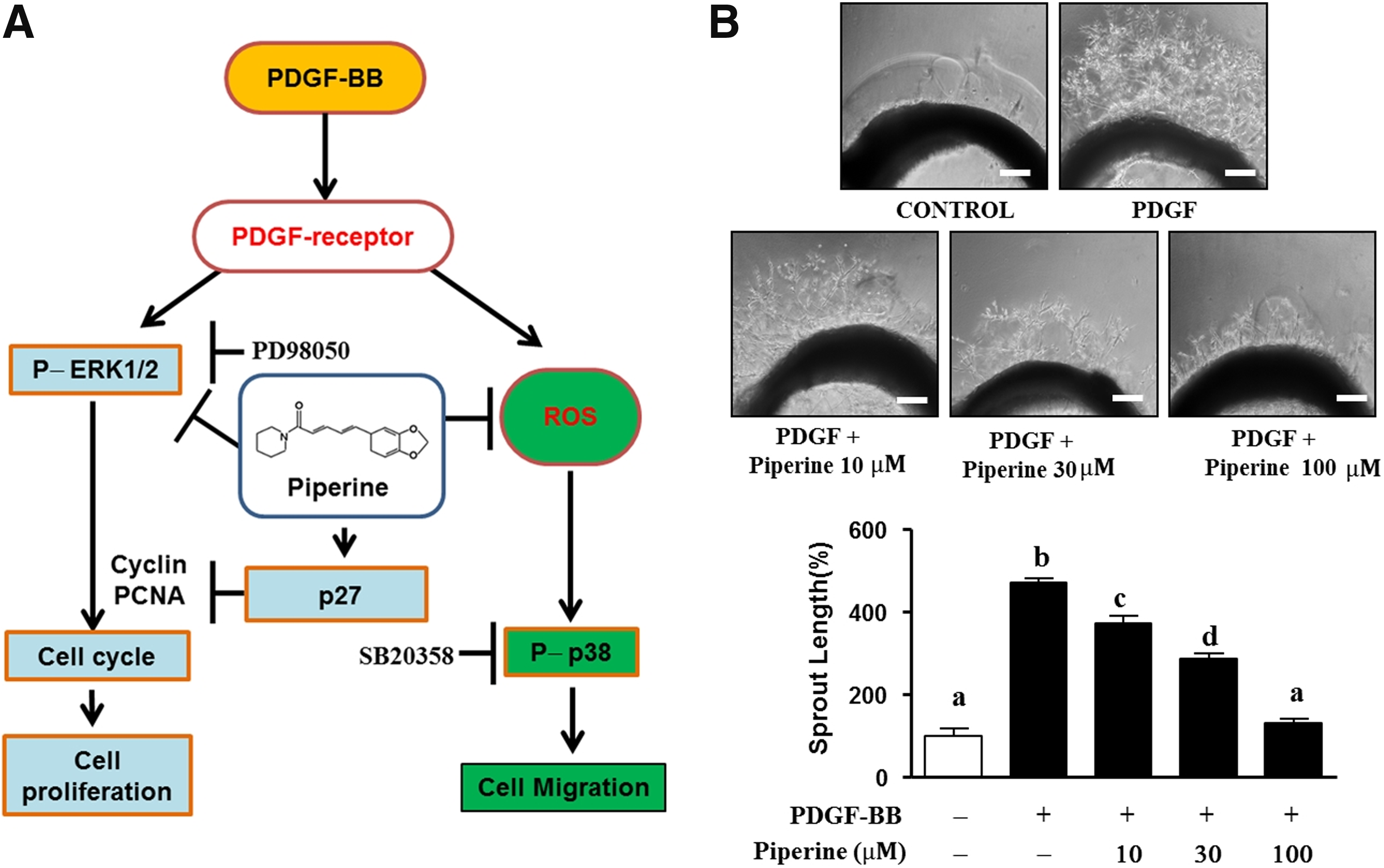
Effects of piperine on the PDGF-BB-related ERK1/2 and p38 pathways in VSMCs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.