
Abstract
Butein and luteolin are members of the flavonoid family, which displays a variety of biological activities. In this study, we demonstrated that butein and luteolin exert anti-inflammatory activities in RAW264.7 macrophages by inducing heme oxygenase-1 (HO-1) expression. Butein and luteolin dose-dependently attenuated inducible nitric oxide synthase (iNOS) expression, leading to the suppression of iNOS-derived nitric oxide (NO) production. The inhibitory effect of butein on NO production was greater than that of luteolin. Consistent with this finding, butein also showed higher inhibitory effects on lipopolysaccharide (LPS)-induced translocation of nuclear factor κB (NFκB) and NFκB reporter gene activity in macrophages than luteolin. Furthermore, the expression of HO-1 was dose-dependently induced by butein and luteolin treatments in macrophages. Additionally, the anti-inflammatory activities of butein and luteolin involved the induction of HO-1 expression, as confirmed by the zinc protoporphyrin (ZnPP) treatment (HO-1 selective inhibitor) and HO-1 small interfering (si)RNA system. ZnPP-mediated downregulation and siRNA-mediated knockdown of HO-1 significantly abolished the inhibitory effects of butein and luteolin on the production of NO in LPS-induced macrophages. Consequently, butein and luteolin were shown to be effective HO-1 inducers capable of inhibiting macrophage-derived proinflammatory mechanisms. These findings indicate that butein and luteolin are potential therapeutic agents for the treatment of inflammatory diseases.
Introduction
I
Heme oxygenase-1 (HO-1), a rate-limiting enzyme in heme catabolism, degrades heme to generate carbon monoxide (CO), free iron, and biliverdin, which is subsequently catabolized into bilirubin by biliverdin reductase. HO-1 and its enzymatic by-products provide a host defense mechanism that can protect the body against endotoxin and oxidative injury. 4 Furthermore, it is well known that HO-1 expression or administration of CO inhibits the expression of the proinflammatory iNOS in activated macrophages through the inactivation of NFκB. 5 Therefore, HO-1 has been considered as a potential therapeutic target for inflammatory disease. 6
Flavonoids have shown health-promoting, disease-preventing, and chemopreventive activities. 7 Flavonoids are classified into several groups, such as flavonols, flavones, and chalcones, according to their structural differences. 8 Flavonoids differing by the type and number of substitution patterns show different biological activities. Butein (2′,4′,3,4-tetrahydroxychalcone) is a chalcone compound that belongs to the flavonoid family and exhibits a basic structure of two benzene rings linked by the α,β-unsaturated carbonyl group and a double bond. 9 Because of the specific chemical structure of chalcones, butein acts as a powerful radical scavenger and inducer of the phase II detoxifying enzymes, which are involved in protection against chronic diseases. 10 Butein was reported to induce HO-1 expression through the nuclear factor erythroid 2-related factor 2 (Nrf2) pathway in human dental pulp cells and primary hepatocytes and decreased LPS-stimulated inflammatory response in RAW264.7 cells. 11
Luteolin (3′4′,5,7-hydroxyl-flavone) belongs to the flavone group of flavonoids, which consists of two benzene rings (A and B) linked through a heterocyclic pyrane ring (C), and usually occurs in glycosylated forms in yellow-green plants, such as celery, green peppers, and olive oil. 12 Several studies have reported that luteolin has antimutagenic, antioxidant, and anti-inflammatory activities. 13 Luteolin was also reported to inhibit LPS-induced inflammatory response through NFκB/AP-1/PI3K-Akt signaling cascades in RAW264.7 cells. 14
Even though there have been several studies about inhibitory effects of butein and luteolin on LPS-stimulated inflammatory response in RAW264.7 cells, no reports have found that the anti-inflammatory effects of the two compounds are related to HO-1 expression. Moreover, to date, there have been no published studies about the comparison of anti-inflammatory activities between butein and luteolin, two flavonoids without and with C ring structures, respectively. Therefore, this study aimed to compare inhibitory effects of butein and luteolin on LPS-stimulated nitric oxide (NO) production through expression of HO-1 and to investigate their underlying molecular mechanisms in RAW264.7 macrophage cells.
Materials and Methods
Chemicals
Luteolin, LPS, 3-(4,5-dimethyl thiazol 2-yl)-2,5-diphenyltetrazolium (MTT), dimethyl sulfoxide (DMSO), Griess reagent, and other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA). Butein was obtained from Toronto Research Chemicals (Downsview, ON, Canada), Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), and penicillin–streptomycin were purchased from Invitrogen (Carlsbad, CA, USA). Antibodies against iNOS, β-actin, HO-1, NFκB p65, NFκB p50, proliferating cell nuclear antigen (PCNA), and peroxidase-conjugated secondary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). ECL™ detection reagents were purchased from GE Healthcare (Buckinghamshire, United Kingdom). Zinc protoporphyrin (ZnPP) was obtained from Porphyrin Products (Logan, UT, USA). Lipofectamine 2000 was obtained from Invitrogen and the Dual-Glo Luciferase assay kit was obtained from Promega (Madison, WI, USA). All other chemicals were of the purest grade available.
Cell culture
The murine macrophage cell line RAW264.7 was obtained from the Korean Cell Line Bank (Seoul, Korea) and maintained in DMEM supplemented with 10% heat-inactivated FBS, 100 U/mL penicillin, and 100 mg/mL streptomycin in a humidified atmosphere of 5% CO2 at 37°C. Cells grown to 80% confluence were treated with LPS (1 μg/mL) with or without different concentrations of butein and luteolin (0.5, 1.0, 2.5, and 5.0 μM) for 18 h. Butein and luteolin were dissolved in DMSO and the final concentration of DMSO was adjusted to less than 0.1% for all treatments.
Cytotoxicity assay
The cytotoxic effects of butein and luteolin on RAW264.7 cells were estimated by measuring lactate dehydrogenase (LDH) levels released from the cytosol of damaged cells. The cells were cultured in wells of 48-well plates at a density of 2.0×105 cells per well. After 6 h, the cells were treated with different concentrations (0.5, 1.0, 2.5, and 5.0 μM) of butein and luteolin with LPS (1 μg/mL) for 18 h. Butein and luteolin were dissolved in DMSO and the final concentration of DMSO was adjusted to less than 0.1% for all treatments. The concentration of LDH in the medium was determined using an LDH cytotoxicity detection kit (Roche Applied Science, IN, USA) according to the manufacturer's instructions.
Nitrite assay
NO production was determined by measuring the amount of nitrite, a relatively stable NO oxidation product. Briefly, RAW264.7 cells (1×105 cells/100 μL) were seeded in 96-well plates and incubated for 6 h. Cells were treated with LPS (1 μg/mL) with or without different concentrations of butein and luteolin (0.5–5.0 μM) for 18 h. Nitrite accumulation in the supernatant was determined by Griess reaction. One hundred microliters of culture supernatant was mixed with an equal volume of Griess reagent and incubated for 10 min at 37°C (5% CO2). Absorbance was read at 550 nm using an ELISA plate reader. NO concentration was determined using dilutions of sodium nitrite as standards.
Western blot analysis
Protein expression levels were examined by western blot analysis. Whole-cell extracts were obtained after treatment by harvesting cells, washing them with ice-cold phosphate-buffered saline, and lysing in a Pro-Prep™ sample buffer (iNtRON Biotech, Seongnam, Korea). To obtain nuclear and cytoplasmic lysates, the cell pellets were lysed using NE-PER® nuclear and cytoplasmic extraction reagents (Pierce Biotechnology, Inc., Rockford, IL, USA) according to the manufacturer's guidelines. The protein concentration of each sample was quantified using the Take3™ Multivolume Plate (Biotek Instruments GmbH, Bad Friedrichshall, Germany). Equal amounts of proteins were denatured and separated on sodium dodecyl sulfate–polyacrylamide gels, and then transferred onto nitrocellulose membranes. Membranes were blocked with 5% nonfat milk in Tris-buffered saline and Tween 20 (TBST) for 1 h and incubated overnight at 4°C with antibodies against iNOS, HO-1, β-actin, p65, p50, and PCNA. After washing with the TBST buffer, the horseradish peroxidase-labeled IgG was added at room temperature for 1 h. Protein bands were visualized on X-ray film activated by chemiluminescence using the supersignal west pico chemiluminescent substrate (Thermo scientific, Tewksbury, MA, USA).
Total RNA isolation and real-time PCR
Total RNA was purified from treated cells using TRIzol RNA isolation reagent (Invitrogen) according to the manufacturer's protocol. cDNA was obtained by reverse transcription of total RNA (0.5–1.0 μg) using the High Capacity cDNA Reverse Transcription Kit (Invitrogen). Quantitative real-time PCR of iNOS was performed using an IQ5 real-time PCR detection system (Bio-Rad, Richmond, CA, USA) by IQ SYBR Green PCR Supermix (Bio-Rad). The forward and reverse primers for iNOS were 5′-ACCATGGAGCATCCCAAGTA-3′ and 5′-CCATGTACCAACCATTGAAGG-3′, respectively. The forward and reverse primers for glyceraldehydes-3-phosphate dehydrogenase (GAPDH) were 5′-AATGTATCCGTTGTGGATCTGA-3′ and 5′-GCTTCACCACCTTCTTGATGT-3′, respectively. The expression levels relative to control were estimated by calculating the ΔΔCt (ΔCt,iNOS−ΔCt,GAPDH) and subsequently using the 2−ΔΔCt method. To confirm HO-1 mRNA expression, the obtained cDNA was analyzed by real-time PCR using a StepOnePlus real-time PCR (Applied Biosystem, Carlsbad, CA, USA) and TaqMan probe-based gene expression analysis (Applied Biosystem). Quantification of gene transcripts for HO-1 (NM_010442.2) and GAPDH (NM_008084.2) was completed using gene-specific primers. Primer sequences are available upon request. Results are expressed as mean±standard deviation after normalizing to the expression of the GAPDH gene using the 2−ΔΔCt method.
Transient transfection and luciferase assay
RAW264.7 macrophages were transfected using the Lipofectamine 2000 and pNFκB-Luc reporter plasmids, according to the manufacturer's instructions. The RAW264.7 macrophages were seeded in a black 96-well plate at a density of 1.5×105 cells per well. Twenty-four hours after the transfection, cells were treated with butein and luteolin with LPS (1 μg/mL). Following 24 h of stimulation, cells were lysed and the luciferase activity was determined using the Promega luciferase assay system and a luminometer (Panomics, Fremont, CA, USA).
HO-1 small interfering RNA transfection
Transient transfections were performed using the Lipofectamine 2000 reagent. HO-1 and stealth RNAi™ small interfering (si)RNA were purchased from Invitrogen. Briefly, 5.0×105 cells were seeded in six-well plates and grown overnight, followed by transfection with 10 nM HO-1 siRNA and 10 nM stealth RNAi using the transfection reagent for 12 h according to the manufacturer's instructions. To confirm the efficacy of HO-1 siRNA, western blotting assay was conducted using the HO-1 antibody.
Statistical analysis
Each experiment was performed in triplicate and repeated thrice. The results are reported as mean±standard error. Statistical comparisons were determined by one-way analysis of variance using SAS version 8.1 (SAS Institute, Cary, NC, USA), followed by Duncan's multiple comparison test. A P-value of<.05 was considered significant.
Results
Effects of butein and luteolin on NO production and iNOS expression in LPS-stimulated RAW264.7 macrophages
The cytotoxicity of butein and luteolin on RAW264.7 cells was evaluated using the LDH assay. The LDH released from the cells indicates a loss of membrane integrity and is a direct measurement of cell death. As shown in Figure 1C, no cytotoxic effects on RAW264.7 cells were observed at the tested concentrations up to 5 μM. Hence, it was concluded that the inhibition of inflammation was not due to butein and luteolin cytotoxicity. Cell exposure to LPS, a bacterial endotoxin, is known to cause the production of iNOS-derived NO that leads to the initiation and amplification of inflammation. To estimate the inhibitory effects of butein and luteolin on the production of iNOS-derived NO in LPS (1 μg/mL)-stimulated RAW264.7 cells, cells were treated with various concentrations (0.5, 1, 2.5, and 5 μM) of butein or luteolin for 18 h. LPS induced a five-fold induction of NO production compared with the native control (Fig. 2A). However, butein and luteolin significantly suppressed LPS-induced production of NO in a dose-dependent manner. Western blot and real-time PCR analysis were also performed to determine whether the inhibitory effects of butein and luteolin on NO production were correlated with the modulation of iNOS expression. As shown in Figure 2B and C, iNOS mRNA and protein levels were dramatically decreased by either butein or luteolin treatment compared with that of LPS, which was in concordance with the decreased NO production. Luteolin at 5 μM showed only a slight inhibitory activity toward iNOS-derived NO production in LPS-stimulated cells, whereas butein strongly inhibited iNOS-derived NO production at the same concentration.

Chemical structures of butein
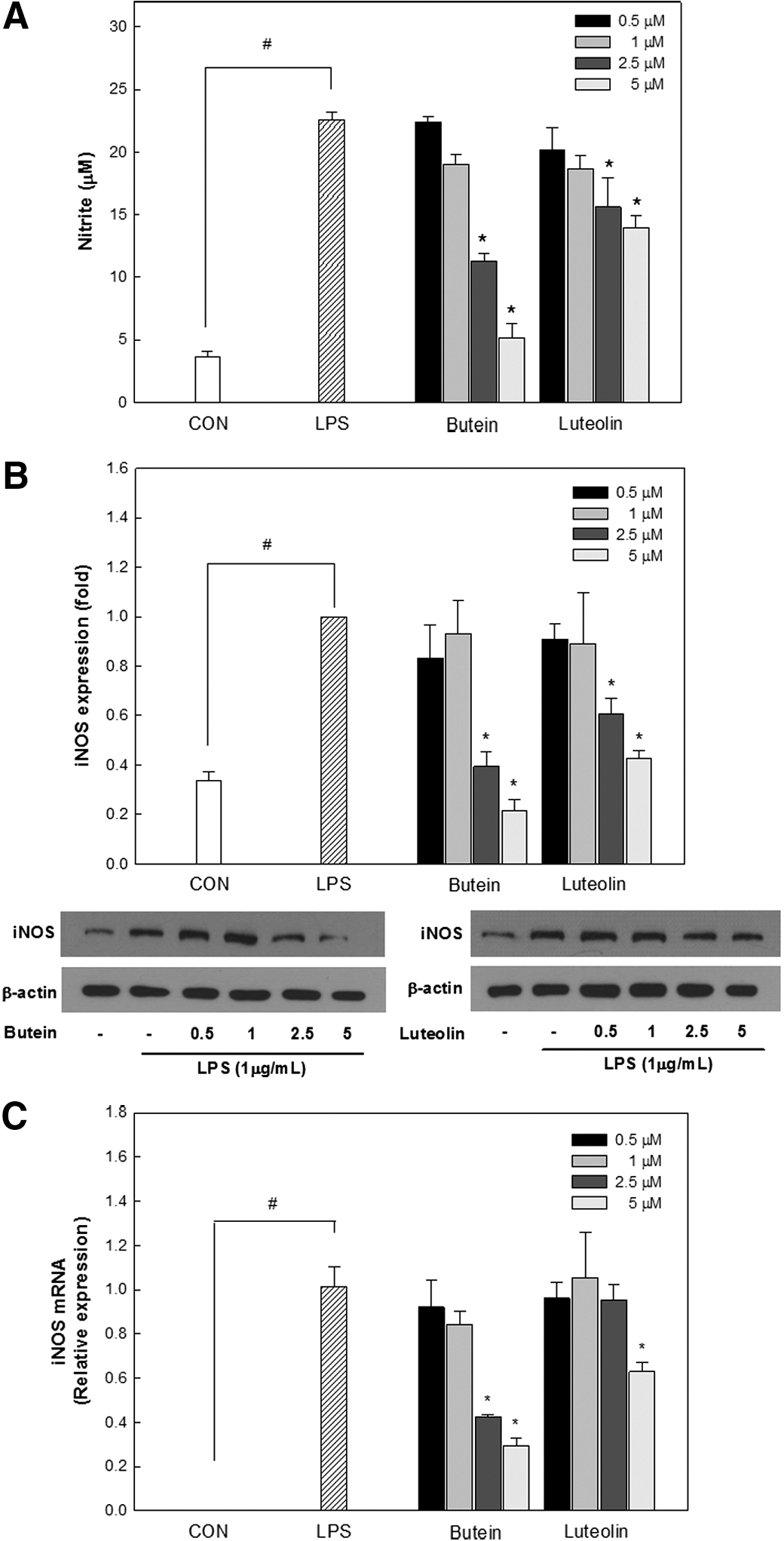
Inhibitory effects of butein and luteolin on LPS-induced nitric oxide (NO) production
Effects of butein and luteolin on nuclear translocation and activation of NFκB in LPS-stimulated RAW264.7 macrophages
Activation of NFκB is thought to play a key role in the LPS-stimulated release of NO production. Therefore, we examined the effects of butein and luteolin on LPS-stimulated NFκB translocation to the nucleus. The p65 and p50 proteins, NFκB subunits, were strongly expressed in the nuclear fraction after stimulation with 1 μg/mL LPS for 1 h (Fig. 3A). The LPS-induced increase in p65 and p50 NFκB levels in the nucleus was significantly inhibited by 5 μM of butein. In contrast, luteolin at the same concentration did not affect this response, suggesting that the anti-inflammatory action of luteolin was weaker than that of butein. To measure the effects of butein and luteolin on the activity of an NFκB-responsive promoter, we examined the activity of the luciferase reporter gene driven by the NFκB-binding consensus concatemer (pNFκB-Luc). As shown in Figure 3B, a significant increase in luciferase activity was observed after stimulation with LPS, suggesting that NFκB was activated by LPS. However, the NFκB luciferase activity was markedly reduced by butein and luteolin at 5 μM. In accordance with NFκB p65 and p50 protein expression in the nucleus, the effect of luteolin on the NFκB promoter activity was also weaker compared with that of butein.
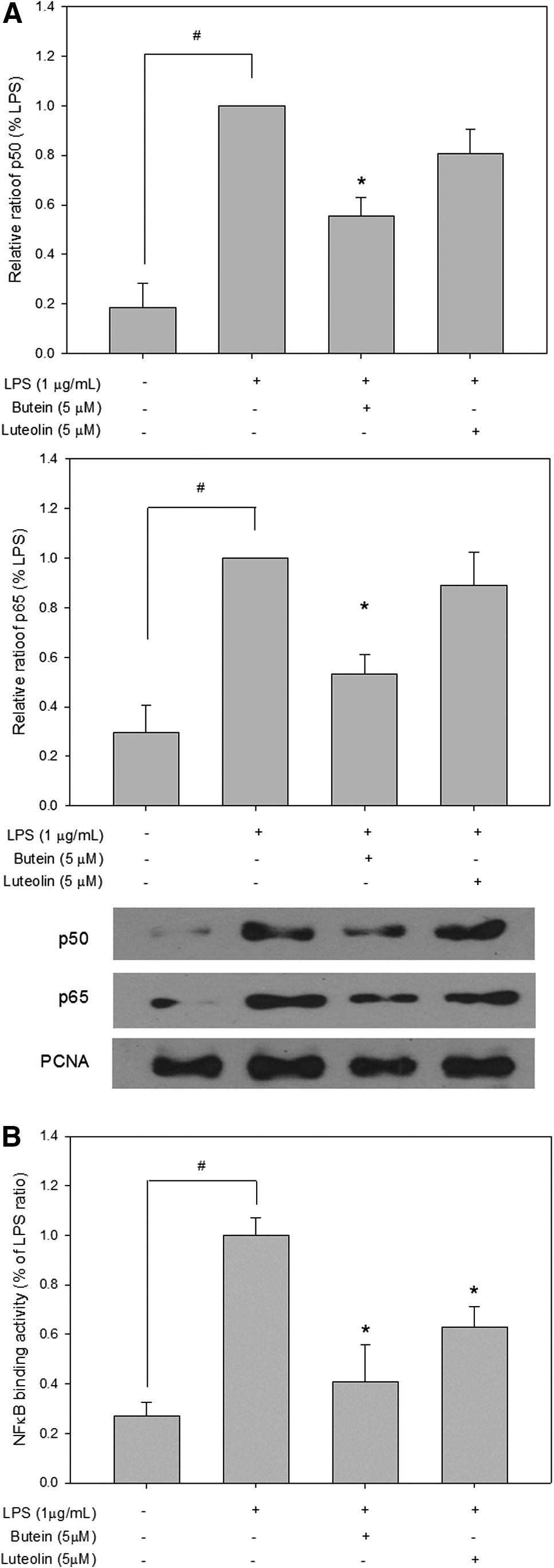
Inhibitory effects of butein and luteolin on LPS-induced nuclear factor κB (NFκB) translation
Induction of HO-1 expression by butein and luteolin in RAW264.7 macrophages
HO-1 plays a critical role in regulating biological responses, including oxidative injury and inflammation. Induction of HO-1 enables the anti-inflammatory function in macrophages. 15 To examine the effects of butein and luteolin on HO-1 expression, RAW264.7 cells were treated with butein and luteolin at 0.5–5 μM for 24 h, and then HO-1 protein and mRNA levels were measured by western blot analysis and real-time PCR, respectively. As shown in Figure 4, butein and luteolin prominently increased HO-1 protein and mRNA levels in a dose-dependent manner.
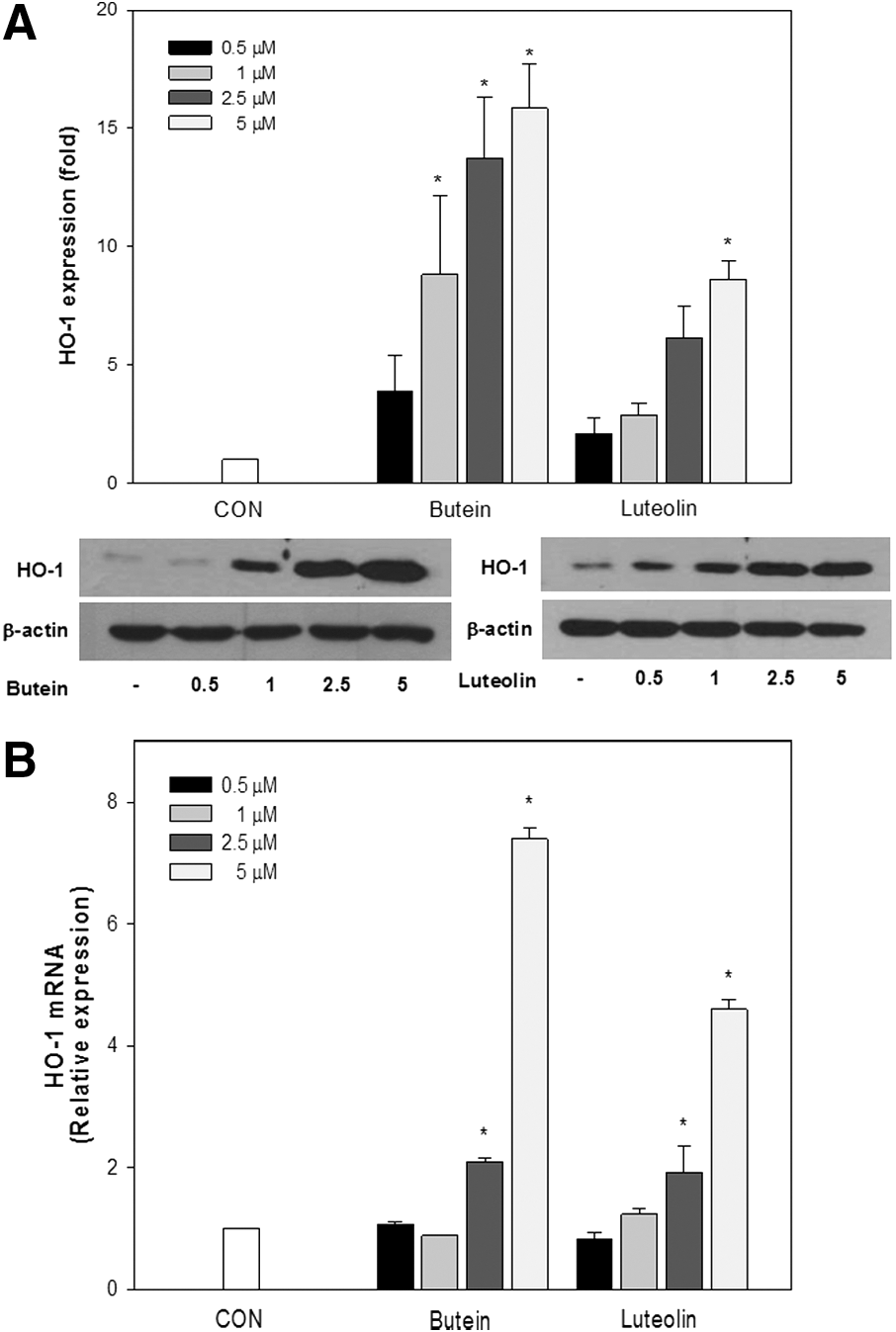
Butein and luteolin-induced heme oxygenase-1 (HO-1) protein
Effects of HO-1 expression on the inhibition of LPS-induced NO production by butein and luteolin
To determine whether HO-1 mediates the inhibitory effects of butein and luteolin, RAW264.7 cells were transfected with HO-1-specific siRNA. Western blot analysis revealed that butein and luteolin-induced HO-1 expression were significantly suppressed following HO-1 knockdown compared with cells transfected with stealth siRNA (Fig. 5A). Inhibitory effects of butein and luteolin on NO production were rescued by HO-1 siRNA transfection (Fig. 5B), suggesting that HO-1 plays a critical role in the suppressive effects of butein and luteolin.
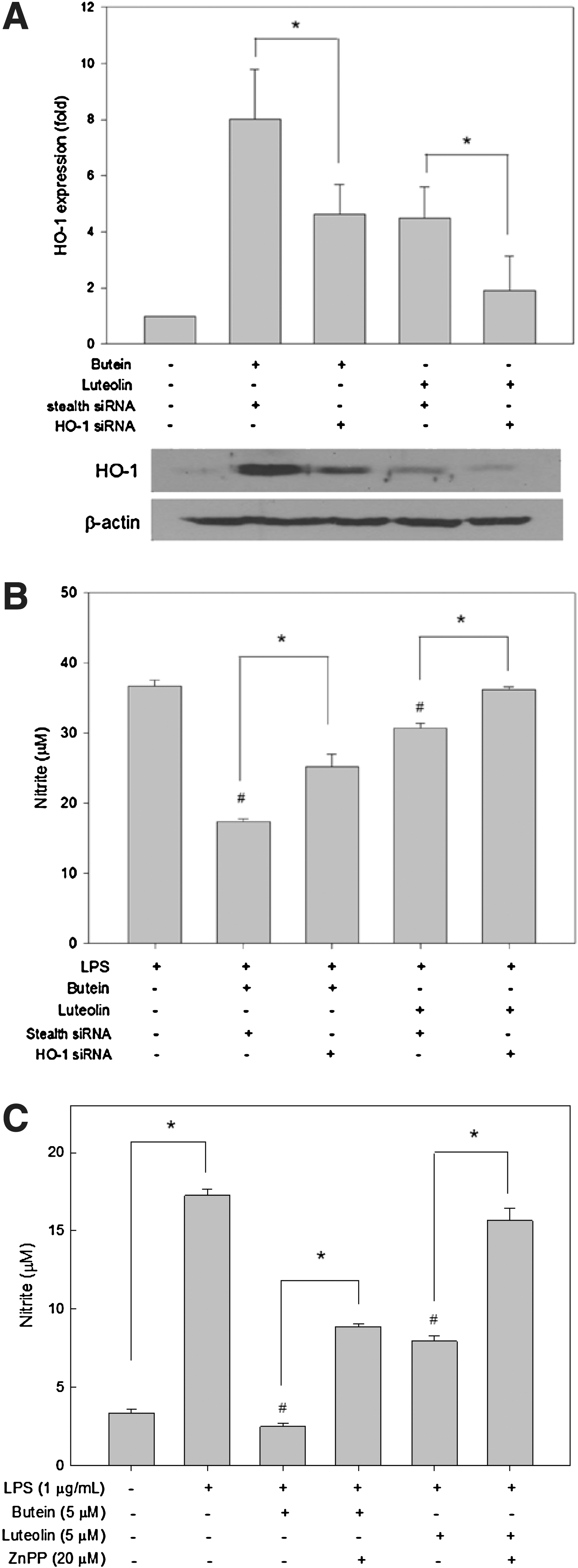
Induction of HO-1 expression by butein and luteolin mediates the suppression of LPS-induced NO production in RAW264.7 cells.
The involvement of HO-1 in the anti-inflammatory activities of butein and luteolin was also confirmed using a selective HO-1 competitive inhibitor, ZnPP. Consistent with the results obtained using the HO-1 siRNA system, ZnPP treatment also reversed the inhibitory effects of butein and luteolin on LPS-stimulated NO production (Fig. 5C). These data suggest that the HO-1 enzyme could play a key role in the anti-inflammatory effects of butein and luteolin in macrophages.
Discussion
Flavonoids generally share a common structural feature, which consists of two aromatic carbon rings, constituting benzopyran (A and C rings) and a benzene (B ring). It is well known that the degree of the double bond at C2–C3 in the C ring and the substituted number of hydroxyl groups on the A and B rings are important for flavonoid biological activities, including antioxidant, anti-inflammatory, and anticancer actions. 16 Although many studies investigated the biological activities of different flavonoids, differing by the type and number of substitution patterns, the anti-inflammatory mechanism of flavonoids with or without C ring structures remains unclear. Therefore, this study was designed to investigate whether butein and luteolin, two flavonoids without and with C ring structures, respectively, could decrease LPS-induced inflammatory response in RAW264.7 macrophages and explore the possible molecular mechanism involved in this anti-inflammatory action.
Excessive amounts of NO are released through the activity of its inducible enzyme iNOS that is expressed predominantly in activated macrophages and plays an important role in inflammatory conditions. 17 Thus, the regulation of proinflammatory mediators in LPS-stimulated macrophages is considered as one of the strategies for developing therapeutic agents against various inflammatory diseases. Upregulation of iNOS is mediated by activation of NFκB in cells stimulated with LPS. The activated NFκB complex translocates from the cytoplasm to the nucleus where it can be involved in the transcription of specific proinflammatory genes such as iNOS. 18 In this study, we observed that butein and luteolin decreased NO production as well as the expression of iNOS in LPS-activated macrophages and that this effect was mediated by blocking the NFκB function. The inhibitory effect of luteolin on iNOS-derived NO induction was significantly lesser than that of butein, which lacks the C ring. Consistent with these findings, butein obviously reduced the NFκB p65 and p50 subunit nuclear translocation and NFκB promoter-binding activity, while luteolin modestly lowered NFκB subunit translocation and NFκB promoter-binding activity. Thus, our results suggested that the suppression of NFκB activation may be the mechanism by which butein and luteolin inhibit the induction of iNOS-derived NO in LPS-stimulated RAW264.7 cells.
HO-1 is a cytoprotective enzyme, which catalyzes the degradation of heme to biliverdin, carbon monoxide, and free iron. Its metabolites have been shown to provide protection against the oxidative and inflammatory stimuli. 19 HO-1 gene expression is mainly mediated by transcription factor Nrf2. Nrf2 is basally confined to the cytoplasmic complex through binding to inhibitory protein Kelch-like ECH-associated protein 1 (Keap1). After exposure of cells to several antioxidants, Nrf2 is released from Keap1, resulting in translocation of Nrf2 to the nucleus. Nrf2 heterodimerizes with small Maf proteins in the nucleus and activates the transcription of genes containing the antioxidant response element located in the promoter regions of HO-1. 15,19 Accumulating evidence demonstrates that induction of HO-1 expression plays a significant protective role by inhibiting the production of proinflammatory mediators in LPS-stimulated macrophages and is a potential therapeutic strategy for treating inflammatory diseases. 15,20 In this study, HO-1 was dose-dependently induced by butein and luteolin in RAW264.7 cells. We also found that the effect of butein on HO-1 induction was significantly greater compared with that of luteolin. Studies on structure–activity relationships have shown that the presence of the α,β-unsaturated carbonyl moiety and hydroxyl groups in various phytochemicals appears to be important for the antioxidant and free radical scavenging activities of these compounds. 21 Curcumin, okanin, and phloretin, which possess a chemical structure similar to that of butein, have been reported to induce HO-1 expression. 22,23 Moreover, some compounds presenting α,β-unsaturated carbonyl groups such as chalcones have been shown to be potent HO-1 modulators due to the inactivation of the Nrf2–Keap1 complex and the activation of Nrf2 nuclear accumulation. 24 Therefore, our results suggest that butein with an α,β-unsaturated carbonyl moiety may be a biological sensor of molecules such as the Nrf2–Keap1 complex. We also demonstrated that downregulation of iNOS-derived NO is likely associated with its ability to induce HO-1 expression. Inhibition of HO-1 activity by ZnPP abolished the suppressive effect of butein or luteolin treatment on LPS-stimulated NO production. HO-1 siRNA knockdown also markedly reversed the inhibitory effects of butein and luteolin on NO production in LPS-induced inflammation in a macrophage model. Our data confirmed that butein and luteolin decrease NO production through the modulation of HO-1 expression. Macrophages stimulated with LPS produce several proinflammatory cytokines, such as tumor necrosis factor (TNF-α), interleukin (IL-1β), and IL-6, as well as iNOS through the NFκB signaling pathway. 25 In addition, several previous studies reported that NO can promote TNF-α production in mouse macrophages and TNF-α can induce the release of IL-6. 26 Therefore, it is expected that butein and luteolin might decrease the release of proinflammatory cytokines in LPS-stimulated macrophages because these two compounds suppressed NO production, which can promote cytokine production, accompanied by inhibiting LPS-induced translocation of NFκB and NFκB reporter gene activity in macrophages. These findings suggest that the anti-inflammatory effects of butein and luteolin were at least partially attributable to the induction of HO-1 expression. Although the exact mechanisms involved in HO-1 anti-inflammatory effects were not fully elucidated in this study, we speculated that the HO-1 enzymatic by-product could be evaluated as possible factors.
In conclusion, the above results demonstrated that both butein and luteolin alleviate LPS-induced inflammatory events by suppression of NFκB activation and the induction of HO-1 expression in RAW264.7 macrophages. Moreover, our data show that the butein anti-inflammatory effect is greater than that of luteolin. Although additional studies are required to determine a direct correlation between the compound structures and molecular genes related to the inflammatory mechanism, we suggest that phytochemicals presenting α,β-unsaturated carbonyl groups may be potent natural agents for modulating inflammatory changes in macrophages.
Footnotes
Acknowledgment
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2012R1A1A4A0105273), Republic of Korea.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.