
Abstract
The immunomodulatory effects of a dietary supplement of purple sweet potato extract (PSPE) in LP-BM5 murine leukemia virus (MuLV)-induced immune-deficient mice were investigated. Mice were divided into six groups: normal control, infected control (LP-BM5 MuLV infection), positive control (LP-BM5 MuLV infection+dietary supplement of red ginseng 300 mg/kg), purple sweet potato water extract (PSPWE) (LP-BM5 MuLV infection+dietary supplement of PSPE 300 mg/kg), PSP10EE (LP-BM5 MuLV infection+dietary supplement of 10% ethanol PSPE 300 mg/kg), and PSP80EE (LP-BM5 MuLV infection+dietary supplement of 80% ethanol PSPE 300 mg/kg). Dietary supplementation began on the day of LP-BM5 MuLV infection and continued for 12 weeks. Dietary supplementation of PSPE inhibited LP-BM5 MuLV-induced splenomegaly and lymphadenopathy and attenuated the suppression of T- and B-cell proliferation and T helper 1/T helper 2 cytokine imbalance in LP-BM5 MuLV-infected mice. Dietary supplement of PSPE increased the activity of the antioxidant enzymes, superoxide dismutase and glutathione peroxidase. The data suggest that PSPE may ameliorate immune dysfunction due to LP-BM5 MuLV infection by modulating antioxidant defense systems.
Introduction
T
Murine AIDS is induced by LP-BM5 murine leukemia virus (MuLV). The murine infection is similar to human AIDS and is characterized by immune dysfunction. 3 Infection of LP-BM5 MuLV leads to profound splenomegaly, lymphadenopathy syndrome, and hypergammaglobulinemia. 4 Cytokine dysregulation resulting from LP-BM5 MuLV infection leads to disturbed balance between T helper 1 (Th1)-type cytokines, including interleukin (IL)-2 and interferon-gamma (IFN-γ), and T helper 2 (Th2)-type cytokines, such as IL-4, IL-10, and IL-13, and increased levels of tumor necrosis factor-alpha (TNF-α). Mice infected with LP-BM5 MuLV display attenuated CD4+ T helper cell function, which contributes to the decreased Th1-type cytokine production and increased Th2-type cytokine production. 5 –7
Overproduction of TNF-α caused by HIV-induced activated macrophages can provoke excessive formation of reactive oxygen species (ROS) and oxidative stress. 8 –10 ROS play important roles in the replication of HIV through the expression of nuclear factor-kappa B (NF-κB) during AIDS. Thus, TNF-α-mediated HIV replication can be inhibited by antioxidants. 11,12
Anthocyanins, which are antioxidant flavonoids, are natural red, violet, and blue pigments found in many fruits and vegetables. There is increasing evidence that purple sweet potato (Ipomoea batatas L.) is an efficient free radical scavenger that can help the antioxidant defense system because of its constituent anthocyanins. 13 –15 Physiological effects of purple sweet potato include antioxidant, 13,14 antiobesity, 13 antitumor, 14 and anti-inflammation. 15 Purple sweet potato decreases the overproduction of TNF-α in lipopolysaccharide (LPS)-induced acute inflammation. 15
The collective data indicate that purple sweet potato might be immunomodulatory in LP-BM5 MuLV-induced murine AIDS. The purpose of this study was to investigate the immunomodulatory effect of purple sweet potato extract (PSPE) in an LP-BM5 MuLV-induced immune-deficient mouse model.
Materials and Methods
Plant material and extract preparation
Purple sweet potato powder (Mu-an, Korea) was extracted by boiling in water at 100°C, with 10% or 80% ethanol at 10°C for 4 h in a reflux apparatus. Extracts were filtered, concentrated in vacuo, and dried with a lyophilizer. Yields of purple sweet potato water extract (PSPWE), purple sweet potato 10% ethanol extract (PSP10EE), and purple sweet potato 80% ethanol extract (PSP80EE) were 15%, 15%, and 16%, respectively. Each extract was stored in a tight and light-protected container at −20°C until used.
Determination of anthocyanin content
Quantitative determination of anthocyanin in the extracts was performed as previously detailed. 16 Each extract (20 mg) was extracted with 50 mL ethanol containing 15% HCl (1.5 M) and filtered. The absorbance was measured using a spectrophotometer (Agilent Technologies, Mississagua, ON, Canada) at a wavelength of 520 nm. The anthocyanin concentration in PSPWE, PSP10EE, and PSP80EE was 0.18±0.01 mg/g, 0.20±0.02 mg/g, and 0.18±0.02 mg/g, respectively.
Experimental mice and treatment
The protocol was approved by the Institutional Animal Care and Use Review Committee of Kyung Hee University (KHUASP[SE]-14-003). Female C57BL/6 mice (4 weeks old, 16±1 g) were purchased from Daehan Biolink (Eumseong, Korea). Mice were maintained at 24°C±1°C and 50–60% relative humidity in climate-controlled quarters with a 12-h light/12-h dark cycle. The mice were acclimatized for 6 days before the experiment, fed standard pellet chow, and provided with fresh water ad libitum.
Forty-eight mice were randomized into six groups (n=8 per group): normal control, infection control (LP-BM5 MuLV infection), positive control (LP-BM5 MuLV infection+dietary supplement of red ginseng 300 mg/kg), PSPWE (LP-BM5 MuLV infection+dietary supplement of PSPWE 300 mg/kg), PSP10EE (LP-BM5 MuLV infection+dietary supplement of PSP10EE 300 mg/kg), and PSP80EE (LP-BM5 MuLV infection+dietary supplement of PSP80EE 300 mg/kg). Dietary supplementation began at the day of LP-BM5 MuLV infection and was continued for 12 weeks. All mice were fed experiment diets based on the AIN93G diet.
Infection with LP-BM5 MuLV
LP-BM5 MuLV (generously donated from NIAID, Bethesda, MD, USA) was administered intraperitoneally to mice in a 100 μL minimum essential medium with an ecotropic titer (XC) of 4.5 log10 plaque forming units/mL, which induced disease with a time course comparable with that previously published. 17 When murine AIDS developed by 12 weeks after infection, all mice were killed by cervical dislocation. Spleens of all the mice were dissected and subjected to primary splenocyte culture, and blood samples were collected for assay of antioxidant enzyme activity.
ELISA for cytokines in splenocytes
The culture response of primary splenocytes was assayed as described previously. 18 Spleens were gently teased with a 40 mesh strainer in the RPMI 1640 medium containing 10% fetal bovine serum, 2 mM glutamine, and 1×105 U/L of penicillin and streptomycin. Splenocytes were prepared by lysing the red blood cells with a red blood cell lysing buffer (Sigma-Aldrich, St. Louis, MO, USA) at 37°C for 3 min. Splenocytes were washed twice with the same medium, plated in 96-well tissue culture dishes (1×106 cells/well), and stimulated with concanavalin A (Con A, 5 μg/mL; Sigma-Aldrich) to determine their production of IL-2, IL-4, and IL-10 after a 24-h incubation, IFN-γ after a 72-h incubation at 37°C (5% CO2 incubator), and with LPS (5 μg/mL; GibcoBRL, Grand Island, NY, USA) to induce TNF-α production for the 24-h incubation. After incubation, the supernatants were collected and analyzed. The levels of cytokines were measured using DuoSet ELISA Development Kits (R&D Systems, Minneapolis, MN, USA).
Splenic T- and B-cell proliferation
Splenic T- and B-cell proliferation was determined by EZ-CyTox (Daeil Lab Service, Seoul, Korea). The splenocytes (1×106 cells/well) were plated in 96-well tissue culture dishes and were treated with Con A (5 μg/mL) for T-cell proliferation and LPS (5 μg/mL) for B-cell proliferation for 48 h at 37°C in a 5% CO2 incubator. After incubation, EZ-CyTox 10 μL was added to each well and incubated for 4 h. The absorbance was measured at 340 nm using a VersaMax ELISA microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Assays of antioxidant enzymes
Serum was collected from whole blood by centrifugation at 14,000 g at 4°C for 20 min. The activity of serum superoxide dismutase (SOD) was measured using a WST-1 [2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2.4-disulfophenyl)-2H-tetrazolium, monosodium salt]-based SOD Assay Kit (Dojindo, Kumamoto, Japan) according to the manufacturer's protocol. The sample solution (20 μL) was mixed with 200 μL of WST-1 and dispensed in wells of a 96-well plate. Each well received 20 μL of xanthine oxidase and incubated at 37°C for 20 min. Absorbance was measured at 450 nm. The serum catalase activity was measured using a Catalase Assay Kit (BioVision, Milpitas, CA, USA). Catalase was reacted with hydrogen peroxide (H2O2) to produce water and oxygen. Unconverted H2O2 was reacted with OxiRed™ probe and absorbance was measured at 570 nm. The glutathione peroxidase (GPx) activity was measured according to the manufacturer's protocol (BioVision) with absorbance measured at 340 nm.
Statistical analysis
Results are expressed as mean±standard deviation. The significance of treatment effects was assessed by Duncan's multiple range tests after one-way ANOVA using SPSS PASW Statistic 20.0 for Windows (SPSS, Chicago, IL, USA). Statistical significance was considered at the P<.05 level.
Results and Discussion
Effects of PSPE on organ weights of LP-BM5 MuLV infection-induced mice
The pathogenesis of LP-BM5 MuLV is similar to the human HIV infection. Therefore, LP-BM5 MuLV infection-induced mice have been widely used to study the pathogenesis of HIV/AIDS and to evaluate the immunomodulatory effects of various interventions in animal models. 3 Presently, the immunomodulatory effect of PSPE was evaluated in LP-BM5 MuLV-induced immune-deficient mice. Infection of LP-BM5 MuLV causes pathological changes that include splenomegaly, lymphadenopathy, and fatty liver. Spleen and lymph nodes are peripheral lymphoid organs; infection or autoimmune disorders enlarge these organs. 19 Weights of body, spleen, lymph node, and liver in LP-BM5-infected mice are increased significantly compared to uninfected controls. 20
Weight gains, food efficiency ratio (FER), and organ weights were measured for pathological observation (Table 1). Ginseng was used as the positive control because ginseng is linked with improved human immunity to infection in Korea. 21 Both the weight gain and FER of LP-BM5 MuLV infection-induced mice were significantly higher compared with the normal control. No significant difference among the LP-BM5 MuLV infection-induced groups was evident. Spleen, lymph node, and liver weights of the infection control group (1.70±0.51 g, 1.03±0.29 g, and 1.85±0.37 g, respectively) were significantly higher than those of the normal control (0.09±0.01 g, 0.08±0.01 g, and 0.98±0.13 g, same respective order). This indicated that LP-BM5 MuLV infection had progressed to murine AIDS. The positive control, PSPWE, PSP10EE, and PSP80EE groups displayed significant lower organ weights. The data suggest that dietary supplementation of PSPE protects against an increase in spleen, lymph node, and liver weight induced by LP-BM5 MuLV infection.
Values are mean±SD of eight mice. Different superscript letters show a significant difference at P<.05 as determined by Duncan's multiple range test.
Normal control, no treatment group; infection control, LP-BM5 MuLV infection group; positive control, LP-BM5 MuLV infection with dietary supplement of red ginseng 300 mg/kg; PSPWE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato water extract 300 mg/kg; PSP10EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 10% ethanol extract 300 mg/kg; PSP80EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 80% ethanol extract 300 mg/kg.
Weight gain (g/12 week)=final body weight (g)−initial body weight (g).
FER=weight gain (g)/food intake calories (kcal)×100.
FER, food efficiency ratio; MuLV, murine leukemia virus; PSPWE, purple sweet potato water extract; SD, standard deviation.
Effects of PSPE on T- and B-cell proliferation of LP-BM5 MuLV infection-induced mice
AIDS progression due to HIV infection is characterized by the progressive depletions of the level of cellular CD4+ T helper cells and B cells. However, the detailed molecular mechanisms have not yet been completely elucidated. HIV infection may increase cell apoptosis. HIV-infected apoptosis of the CD4+ T helper cells and B cells causes immune dysfunction, which is important in AIDS immunopathogenesis. 22 –25 Polyclonal B cells are activated during HIV replication, which induces hypergammaglobulinemia and B-cell function loss of response to specific pathogens. 22 Numbers of CD4+ T cells, CD8+ T cells, and CD19 B cells in both early and chronic HIV-infected individuals are reportedly significantly lower compared with uninfected controls. 23 Presently, T- and B-cell proliferation from mitogen-stimulated splenocytes was significantly decreased in the infection control group (% of normal control: T cells, 56.58±1.91; B cells, 68.90±8.11). Dietary supplementation with red ginseng, PSPWE, PSP10EE, and PSP80EE significantly increased T- and B-cell proliferation. There was no marked difference in T- and B-cell proliferation among the positive control, PSPWE, PSP10EE, and PSP80EE groups. Compared with the normal control group, the positive control and PSPWE groups did not show statistical significant difference in B-cell proliferation (Fig. 1). Dietary supplementation with purple sweet potato can induce a significant increase of Con A-induced proliferation of splenocytes and thymocytes compared with the control group in immunized chickens. 26 The prior and present data establish the immunomodulatory effect of purple sweet potato due to inhibition of apoptosis during LP-BM5 MuLV infection.
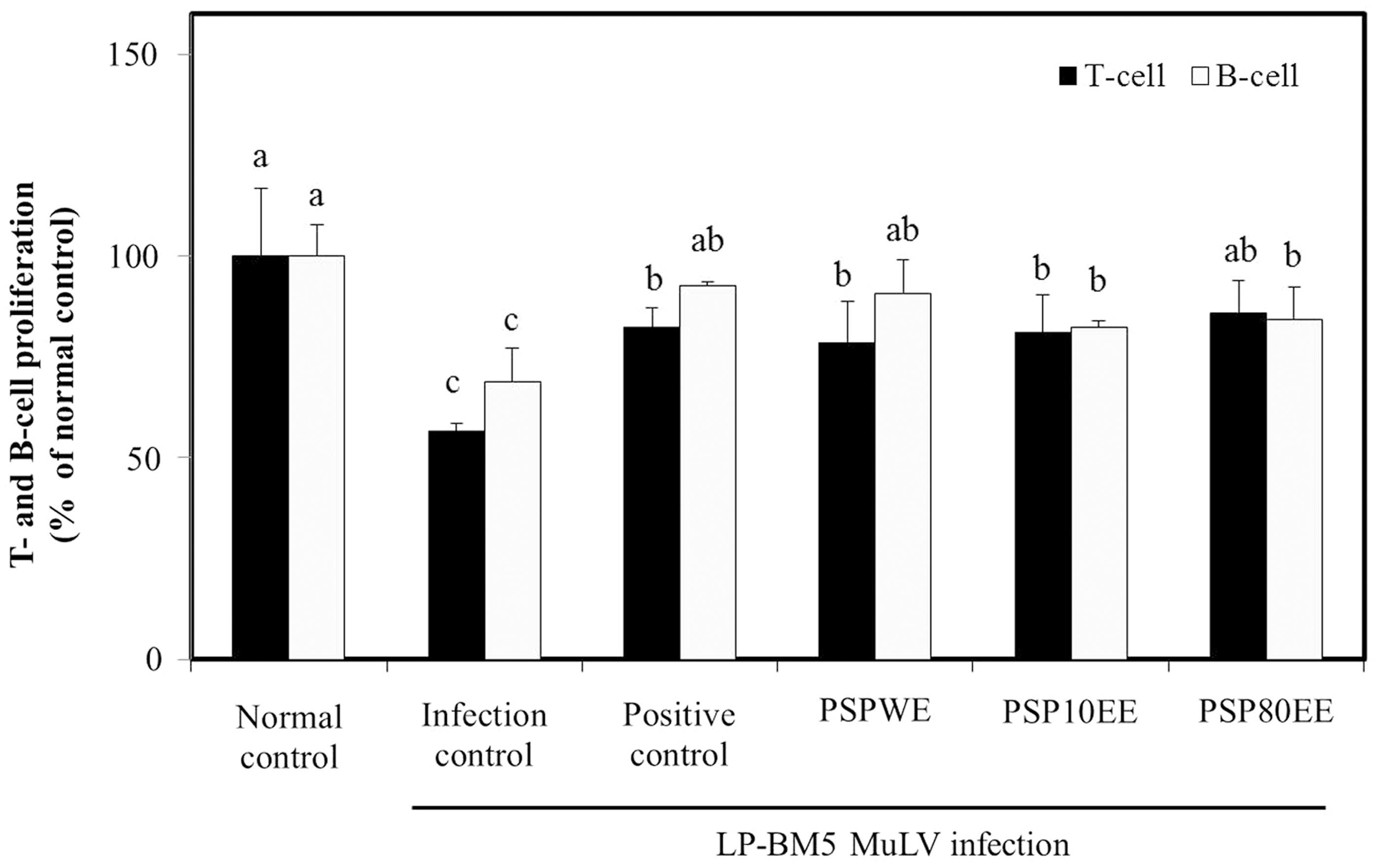
Effects of PSPE on T- and B-cell proliferation from mitogen-stimulated splenocytes prepared from C57BL/6 mice with or without LP-BM5 MuLV infection. Normal control, no treatment group; infection control, LP-BM5 MuLV infection group; positive control, LP-BM5 MuLV infection with dietary supplement of red ginseng 300 mg/kg; PSPWE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato water extract 300 mg/kg; PSP10EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 10% ethanol extract 300 mg/kg; PSP80EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 80% ethanol extract 300 mg/kg. Values are mean±SD of eight mice. Different letters show a significant difference at P<.05 as determined by Duncan's multiple range test. MuLV, murine leukemia virus; PSPE, purple sweet potato extract; PSPWE, purple sweet potato water extract; SD, standard deviation.
Effects of PSPE on Th1-type and Th2-type cytokine production of LP-BM5 MuLV infection-induced mice
T helper cells are important to the immune system response, particularly in the adaptive immune system. These cells are crucial in identifying pathogens in the body and signaling other cells for the immune response and defense by releasing T-cell cytokines. 24 Surfaces of mature T helper cells have protein and are referred to as CD4+ helper T cells. CD4+ helper T cells are found in two different cell types: Th1 and Th2. Th1 cells produce Th1-type cytokines (IL-2 and IFN-γ), while Th2 cells produce Th2-type cytokines (IL-4, IL-10, and IL-13). Th1 cytokines stimulate phagocytosis and destruction of pathogens by the activation of macrophage. Th2 cytokines stimulate the production of immunoglobulins by the activation of B cells and the degradation of mast cells. 27
HIV infection induces the loss of function of CD4+ helper T cells, which can lead to declines in the Th1 cell activity and increased Th2 activity. This imbalanced cytokine production may contribute to AIDS immunopathogenesis. 28 Presently, IL-2 production from ConA-induced splenocytes in the infection control group (7.14±2.02 pg/mL) was significantly lower than the normal control group (649.05±45.72 pg/mL) (Fig. 2). IL-2 is necessary for the proliferation and differentiation of CD4+ helper T cells. HIV/AIDS patients display declines in T-cell proliferation and IL-2 production. 29 IL-2 production in the PSPWE, PSP10EE, and PSP80EE groups was 67.64±26.53 pg/mL, 76.07±19.22 pg/mL, and 43.33±27.64 pg/mL, respectively, which was significantly higher compared with the infection control group. No significant difference was evident among the normal, positive control, PSPWE, and PSP10EE groups. In addition, IFN-γ production from ConA-induced splenocytes of the infection control group (381±117.38 pg/mL) was significantly less than the normal control group (1312.75±112.29 pg/mL). However, dietary supplementation with PSPWE induced a significant increase in IFN-γ production (795.50±48.08 pg/mL), which was not significantly different compared to the positive control group (907.50±90.51 pg/mL). The values for the PSP10EE group (603.25±64.7 pg/mL) and PSP80EE group (522.50±115.38 pg/mL) were not significantly different compared with the infection control (Fig. 2). IFN-γ enhances the function of macrophages and promotes the differentiation of naive T helper cells into CD4+ helper T cells. Suppressed IFN-γ production inhibits the antiviral activity in the immunoregulatory processes. 30 LP-BM5 MuLV infection results in a significant decrease of IL-2 and IFN-γ production when compared with the control group. 31 Presently, dietary supplementation with purple sweet potato diminished the reduction of Th1-type cytokines induced by LP-BM5 MuLV infection. 31
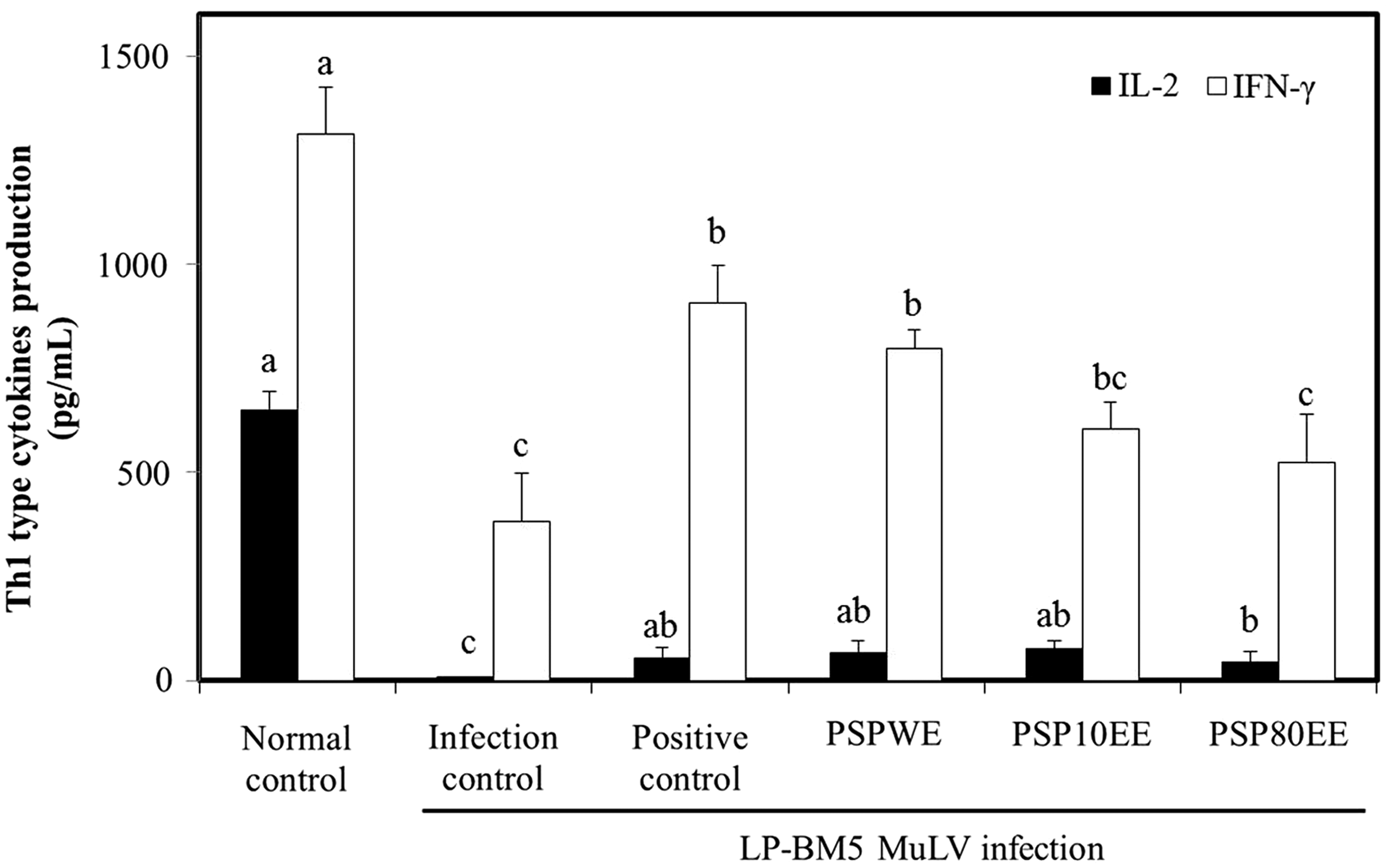
Effects of PSPE on Th1-type cytokine production from mitogen-stimulated splenocytes prepared from C57BL/6 mice with or without LP-BM5 MuLV infection. Normal control, no treatment group; infection control, LP-BM5 MuLV infection group; positive control, LP-BM5 MuLV infection with dietary supplement of red ginseng 300 mg/kg; PSPWE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato water extract 300 mg/kg; PSP10EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 10% ethanol extract 300 mg/kg; PSP80EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 80% ethanol extract 300 mg/kg. Values are mean±SD of eight mice. Different letters show a significant difference at P<.05 as determined by Duncan's multiple range test. Th1, T helper 1.
The production of Th2-type cytokines (IL-4 and IL-10) from ConA-induced splenocytes is presented in Figure 3. IL-4 and IL-10 secretions were significantly higher with LP-BM5 MuLV infection (IL-4: 48.86±10.58 pg/mL, IL-10: 131.67±4.95 pg/mL) compared to the normal control (IL-4: 8.05±2.64 pg/mL, IL-10: 26.06±8.23 pg/mL) (P<.05). IL-4 suppresses HIV replication in human monocyte-derived macrophages, and IL-4 and IL-10 are produced in abnormally high levels in LP-BM5 MuLV infection-induced mice. 32 Dietary supplementation of red ginseng, PSPWE, PSP10EE, and PSP80EE leads to significant differences compared with the infection control group. There were no significant differences among the positive control, PSPWE, PSP10EE, and PSP80EE groups. Similar to our observations, high-purity anthocyanins obtained from chokeberries produce a dose-dependent reduction in IL-10 production in LPS-stimulated monocytes. 33 The present data indicate that concentrated anthocyanin obtained from PSPE attenuates the imbalance of the Th1/Th2 cytokine production and, therefore, is immunomodulatory.

Effects of PSPE on Th2-type cytokine production from mitogen-stimulated splenocytes prepared from C57BL/6 mice with or without LP-BM5 MuLV infection. Normal control, no treatment group; infection control, LP-BM5 MuLV infection group; positive control, LP-BM5 MuLV infection with dietary supplement of red ginseng 300 mg/kg; PSPWE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato water extract 300 mg/kg; PSP10EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 10% ethanol extract 300 mg/kg; PSP80EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 80% ethanol extract 300 mg/kg. Values are mean±SD of eight mice. Different letters show a significant difference at P<.05 as determined by Duncan's multiple range test. Th2, T helper 2.
Effects of PSPE on TNF-α production of LP-BM5 MuLV infection-induced mice
Following stimulation by macrophages, monocytes, neutrophils, T cells, and NK cells, the proinflammatory cytokine, TNF-α, provokes HIV replication and activation by inducing NF-κB and other proinflammatory cytokines (IL-6 and IL-8), which support the upregulation of HIV replication. 8,34 TNF-α production from LPS-induced splenocytes of the infection control group (775.00±73.54 pg/mL) was significantly elevated approximately fourfold compared to the normal control (179.67±55.77 pg/mL) (P<.05; Fig. 4). Several prior reports described the significant elevation of TNF-α production of LPS-stimulated splenocytes in LP-BM5 MuLV infection-induced murine AIDS. TNF-α production plays an important key role in the development of inflammation, which can influence the progression of organ damage. 34 Presently, dietary supplementation with red ginseng, PSPWE, PSP10EE, and PSP80EE resulted in significant decreases of TNF-α production compared with the infection control group, with no significant difference evident among the positive control, PSPWE, PSP10EE, and PSP80EE groups. Purple sweet potato color may bestow neuroprotection by inhibiting the expressions of proinflammatory cytokines and blocking NF-κB signaling. 15 Anthocyanin reportedly inhibits TNF-α production in LPS-induced cells. 35 The prior and present data suggest that anthocyanin of purple sweet potato have advantageous effects on inflammation both in vitro and in vivo.
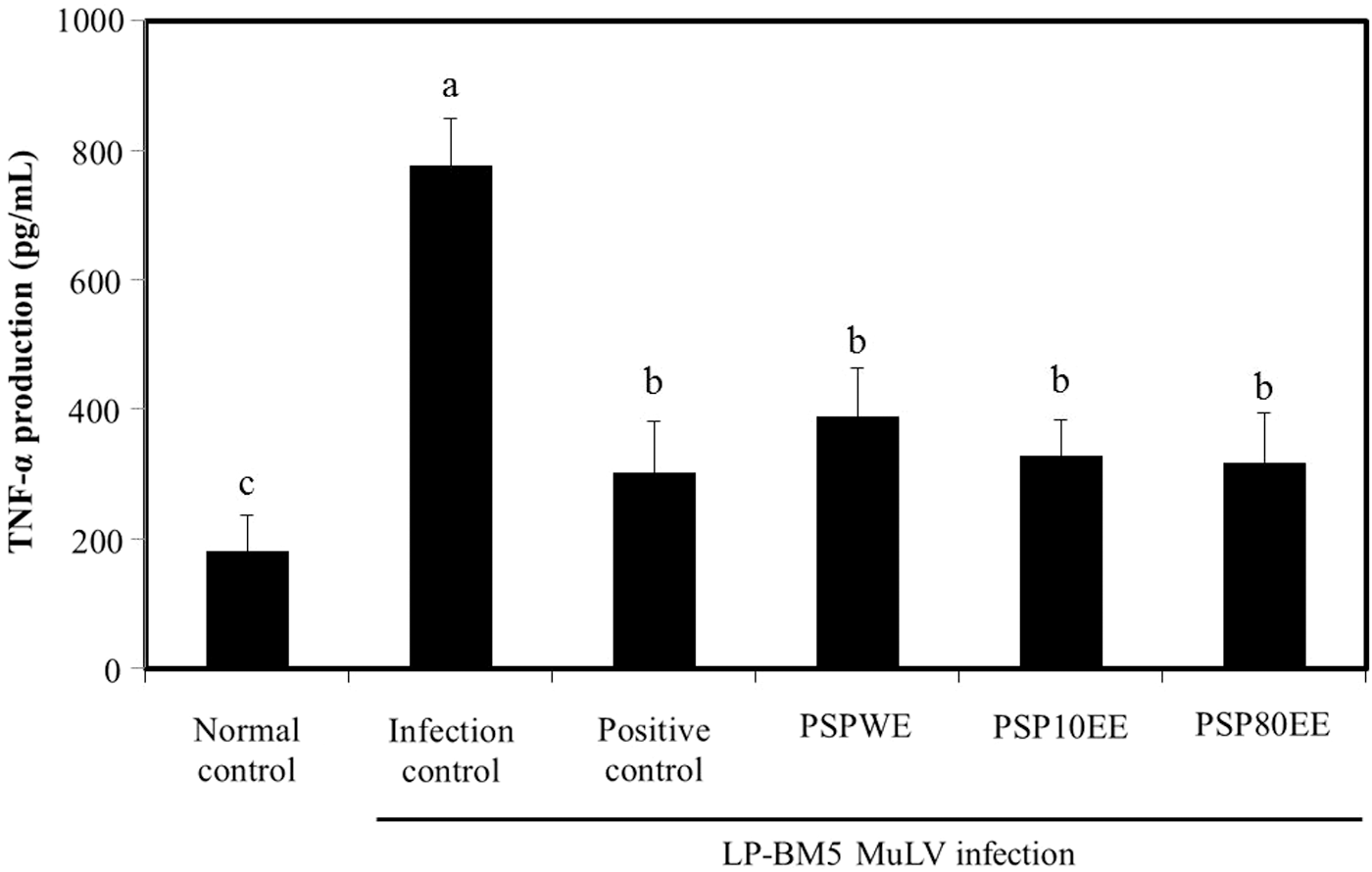
Effects of PSPE on TNF-α production from mitogen-stimulated splenocytes prepared from C57BL/6 mice with or without LP-BM5 MuLV infection. Normal control, no treatment group; infection control, LP-BM5 MuLV infection group; positive control, LP-BM5 MuLV infection with dietary supplement of red ginseng 300 mg/kg; PSPWE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato water extract 300 mg/kg; PSP10EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 10% ethanol extract 300 mg/kg; PSP80EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 80% ethanol extract 300 mg/kg. Values are mean±SD of eight mice. Different letters show a significant difference at P<.05 as determined by Duncan's multiple range test. TNF-α, tumor necrosis factor-alpha.
Effects of PSPE on antioxidant enzyme activity of LP-BM5 MuLV infection-induced mice
No significant differences in the SOD and catalase activity were found between the normal control and infection control groups (Table 2). However, the GPx activity was significantly lower in the infection control group than in the normal control group. Normal cells have antioxidant systems consisting of enzymatic (SOD, catalase, and GPx) and nonenzymatic antioxidants that act to neutralize oxidative stress. 36 LP-BM5 MuLV infection generates excessive quantities of ROS by overproduction of TNF-α, which leads to an imbalance between ROS production and antioxidant defenses. Accordingly, LP-BM5 MuLV infection aggravates antioxidant systems and induces oxidative stress. 37 Oxidative stress accompanied by the dysregulation of enzyme antioxidants induces the enhanced replication of HIV and HIV-related pathogenesis. 9 –11,38 Thus, there is a high correlation between LP-BM5 MuLV infection-induced murine AIDS progression and the reduction of activities of antioxidant enzymes. For this reason, antioxidant supplementation represents an important therapy for the treatment of HIV/AIDS. Several studies have reported that vitamins A, C, and E, the most important antioxidants known, decrease lipid peroxidation and increase the activity of antioxidant enzymes in HIV-infected patients. 39,40
Values are mean±SD of eight mice. Different superscript letters show a significant difference at P<.05 as determined by Duncan's multiple range test.
Normal control, no treatment group; infection control, LP-BM5 MuLV infection group; positive control, LP-BM5 MuLV infection with dietary supplement of red ginseng 300 mg/kg; PSPWE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato water extract 300 mg/kg; PSP10EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 10% ethanol extract 300 mg/kg; PSP80EE, LP-BM5 MuLV infection with dietary supplement of purple sweet potato 80% ethanol extract 300 mg/kg.
GPx, glutathione peroxidase; NS, not significant; SOD, superoxide dismutase.
Presently, no significant difference in the SOD activity was found among the positive control, PSPWE, PSP10EE, and PSP80EE groups, although all the groups displayed a significantly increased SOD activity compared to the infection control group. Furthermore, the positive control, PSPWE, PSP10EE, and PSP80EE groups displayed statistically significant increases in the GPx activity compared to the infection control group, especially the PSPWE group. Purple sweet potato contains a considerable amount of anthocyanins, which are flavonoids with antioxidant activity. 14 Purple sweet potato color inhibits D-galactose treatment-induced reduction of Cu/Zn-SOD and catalase activity, and the level of malondialdehyde (a major maker of lipid peroxidation). 41 Presently, the anthocyanin content in the PSPWE, PSP10EE, and PSP80EE groups was similar. The immunomodulatory effect and the antioxidant activity of PSPE were similar in LP-BM5 MuLV-induced immune-deficient mice.
The present study investigated the immunomodulatory effect of PSPE in an LP-BM5 MuLV-induced immune-deficient mouse model. Dietary supplementation with PSPE attenuated the decreases in T- and B-cell proliferation, antioxidant enzyme activities, and imbalance of the Th1/Th2 cytokines by LP-BM5 MuLV infection. Purple sweet potato may be useful in the prevention of LP-BM5 MuLV infection-induced oxidative stress and might also play a crucial role in immune systems of HIV-infected individuals.
Footnotes
Acknowledgment
This research was financially supported by the Ministry of Knowledge Economy (MKE) and the Korea Institute for Advancement of Technology (KIAT) through the Research and Development for Regional Industry.
Author Disclosure Statement
No competing financial interests exist.