
Abstract
This study examined the beneficial effects of the n-butanol fraction and rutin extracted from tartary buckwheat (TB) on learning and memory deficits in a mouse model of amyloid β (Aβ)-induced Alzheimer's disease (AD). Learning and memory were assessed using the T-maze, object recognition, and Morris water maze tests. Animals administered Aβ showed impaired cognition and memory, which were alleviated by oral administration of an n-butanol fraction and rutin extracted from TB. Similarly, Aβ-induced increases in nitric oxide formation and lipid peroxidation in the brain, liver, and kidneys were attenuated by treatment with n-butanol fraction and rutin from TB in addition to antioxidant effects observed in control (nonAβ-treated) animals. The results of the present study suggest that the n-butanol fraction and rutin extracted from TB are protective against and have possible therapeutic applications for the treatment of AD.
Introduction
A
A major hallmark of AD is the accumulation of neurotoxic amyloid β (Aβ) peptides and their deposition in extracellular plaques. Recent studies indicate that the extracellular accumulation of soluble, high-molecular-weight Aβ oligomers is largely responsible for AD-associated dementia and memory deficits in the Tg2576 mouse model of AD. 3,4 Additionally, Aβ disturbs normal synaptic function and cognitive capabilities 5 with progressive functional disruption accompanied by a loss of neurotrophins (NTs), including nerve growth factor, brain-derived neurotrophic factor (BDNF), NT3, and NT4/5. 6 –8 Aβ-induced disruption of kinases such as extracellular signal-regulated kinases (ERKs) is a critical step in the onset of memory deficit development. 9 The Aβ oligomer inhibits the activation of ERK and thereby decreases active cyclic adenosine monophosphate (cAMP) response element-binding protein (CREB). These findings suggest that a malfunction in cAMP signaling may play an important role in the pathological changes characterizing this disease. Major efforts from both academia and industry are therefore presently focused on developing novel pharmacologic strategies that could delay the initiation and/or slow the oligomerization and aggregation of amyloid peptides. Previous studies have suggested that some polyphenolic compounds could reduce brain amyloid neuropathy and improve cognitive function by promoting nonamyloidogenic α-secretase activity. 10,11
Tartary buckwheat (Fagopyrum tataricum Gaertn., hereafter TB), belonging to the Polygonaceae family, is grown and used in the mountainous regions of southwest China (Sichuan), northern India, Bhutan, and Nepal. 12 TB grain is an important functional food. It contains proteins with a high biological value, a balanced amino acid composition, relatively abundant crude fiber, and vitamins B1, B2, and B6, and contains more rutin than common buckwheat. 13 Buckwheat has numerous bioactive properties, including the ability to increase lactic acid bacteria in rat intestines, treat allergic inflammation, lower serum glucose levels, suppress gallstone formation, reduce cholesterol levels, inhibit protease activity, and promote scavenging of radicals. 14 –16 Rutin is recognized to possess antioxidative, antihypertensive, and anti-inflammatory activities, and buckwheat is the only cereal known to contain rutin in its seeds. 17 –19 TB contains 30–150 times more rutin than the seeds of common buckwheat. 20
Little is known about the protective effects of TB and its active compound, rutin, against AD or their potential to alleviate the cognitive impairment associated with AD. We extracted several fractions from TB and found the n-butanol (n-BuOH) fraction to be a promising active fraction through a series of preliminary tests. In an earlier study, the n-BuOH fraction was found to exhibit a strong antioxidant effect. 15 In this study, we investigated the protective activities of the n-BuOH fraction and rutin extracted from TB against impairments in cognition and memory in an in vivo model of Aβ-induced AD.
Materials and Methods
Plant materials
TB was obtained from the Highland Agriculture Research Center, National Institute of Crop Science, Pyeongchang, Republic of Korea. A voucher specimen of TB (No. LEE 2010-02) was deposited at the Herbarium of Department of Integrative Plant Science, Chung-Ang University, Anseong, Republic of Korea.
Instruments and reagents
Fast atom bombardment mass spectrometry (FAB-MS) was performed with a JMS-600W mass spectrometer (JEOL, Tokyo, Japan). 1 H- and 13 C-nuclear magnetic resonance (NMR) spectra were recorded using a Bruker AVANCE 500 NMR spectrometer (Bruker, Rheinstetten, Germany) in dimethyl sulfoxide (DMSO)-d 6. Aβ 25-35 and malondialdehyde (MDA) were obtained from Sigma Aldrich (Saint Louis, MO, USA). DMSO and sodium chloride (NaCl) were purchased from Bio Basics, Inc. (Markham, Ontario, Canada). Thiobarbituric acid (TBA) was provided by Lancaster Synthesis (Ward Hill, MA, USA). n-Hexane, methylene chloride (MC), ethyl acetate (EtOAc), n-BuOH, and phosphoric acid were acquired from Samchun Pure Chemical Co. (Pyeongtaek, Korea).
Preparation of fractions and rutin from TB
TB (3 kg) was dried and finely powdered, then extracted with methanol (MeOH) for 3 h (4 L×8) under reflux at 65–75°C. The solvent was evaporated in vacuo to obtain the MeOH extract (147.2 g). This extract was suspended in distilled water and partitioned with n-hexane (14.9 g), MC (9.7 g), EtOAc (3.0 g), and n-BuOH (10.3 g), successively. 21 Rutin was detected and isolated from the EtOAc and n-BuOH fractions through recrystallization with MeOH (Fig. 1).
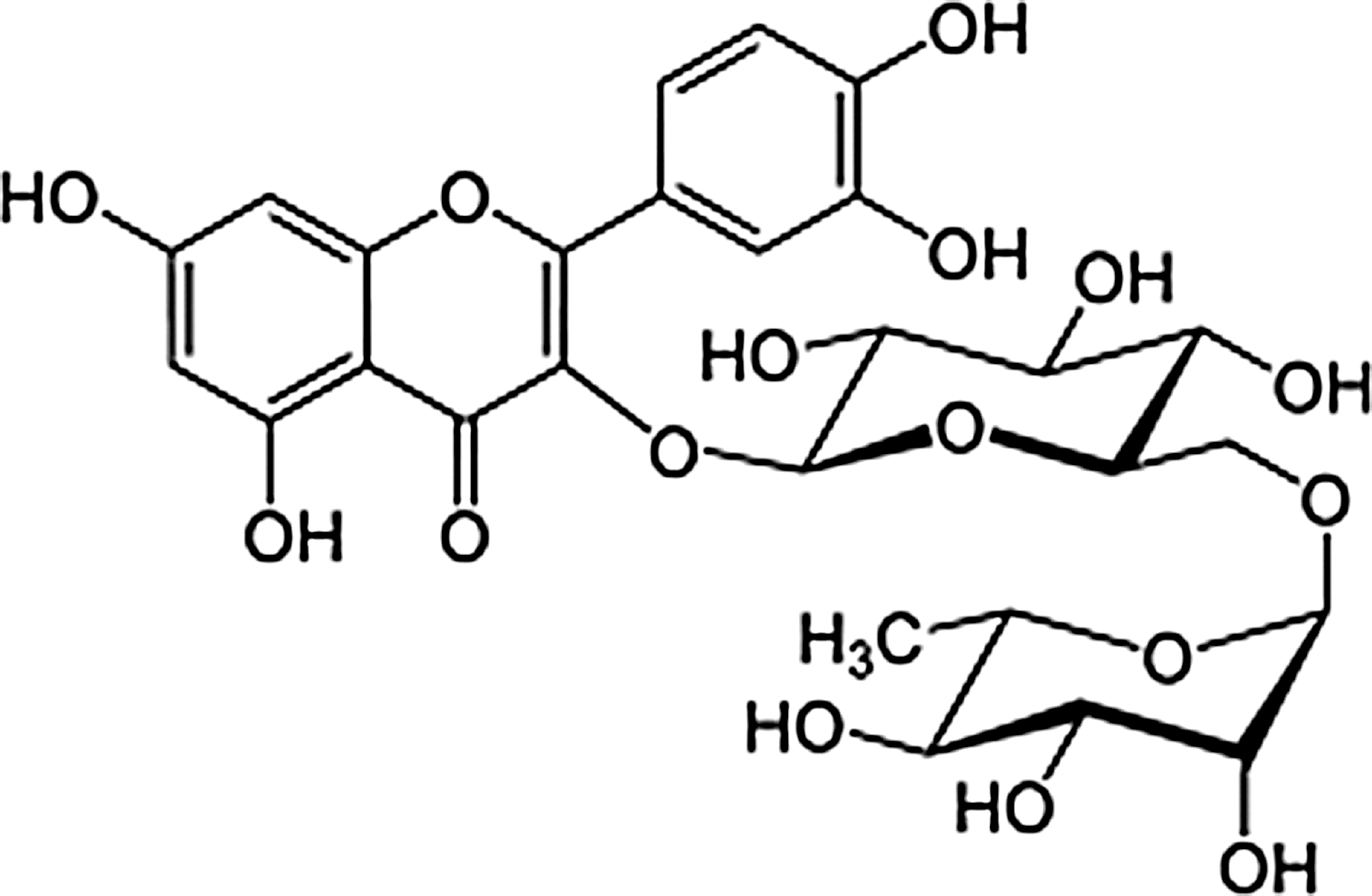
Structure of rutin from tartary buckwheat (TB).
Rutin: FAB-MS: m/z 611 [M+H]+ (Fig. 2); 1 H-NMR (500 MHz, DMSO-d 6): δ 1.00 (3H, d, J=6.0 Hz, Rha-CH3), 3.07–3.69 (12H, m, J=2.0 Hz, sugar H), 4.38 (1H, s, Rha H-1), 5.34 (1H, d, J=7.5 Hz, Glc H-1), 6.19 (1H, d, J=1.2 Hz, H-6), 6.38 (1H, d, J=1.2 Hz, H-8), 6.84 (1H, d, J=8.5 Hz, H-5′), 7.54 (1H, d, J=2.5 Hz, H-2′), 7.55 (1H, dd, J=2.5, 8.5 Hz, H-6′), 12.58 (1H, s, 5-OH); 13C-NMR (125 MHz, DMSO-d 6): δ 156.5 (C-2), 133.2 (C-3), 177.3 (C-4), 161.1 (C-5), 98.6 (C-6), 164.1 (C-7), 93.5 (C-8), 156.5 (C-9), 103.9 (C-10), 121.5 (C-1′), 115.2 (C-2′), 133.2 (C-3′), 148.4 (C-4′), 116.2 (C-5′), 121.1 (C-6′), 101.1 (Glc C-1), 74.0 (Glc C-2), 76.4 (Glc C-3), 70.5 (Glc C-4), 75.9 (Glc C-5), 67.0 (Glc C-6), 100.7 (Rha C-1), 70.3 (Rha C-2), 70.0 (Rha C-3), 72.0 (Rha C-4), 68.2 (Rha C-5), 17.7 (Rha C-6).
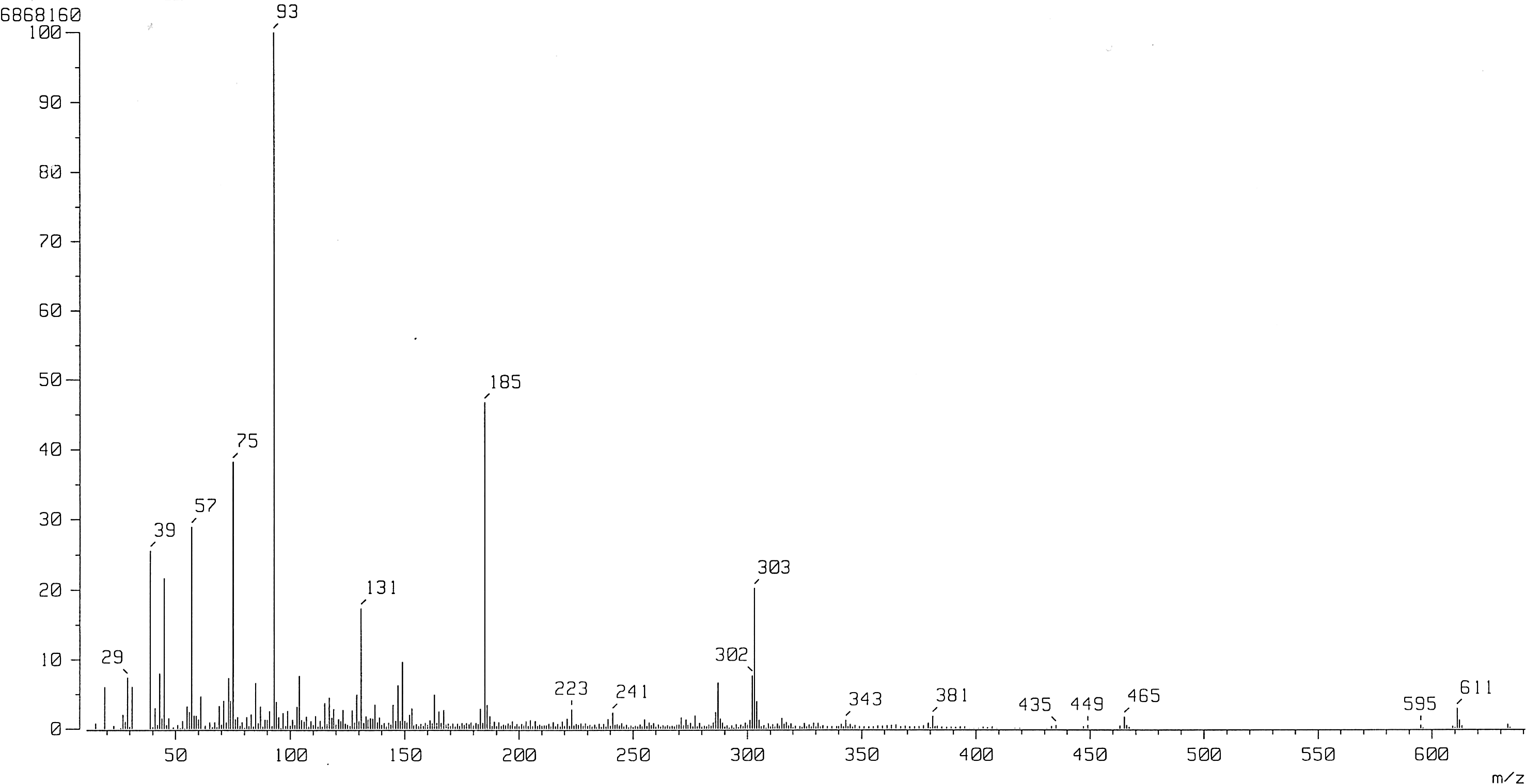
MS spectrum of rutin from TB.
Animals and experimental protocols
Male ICR mice (5 weeks of age; Orient, Inc., Seongnam, Korea) weighing 25–28 g were housed in plastic cages with access to food and water ad libitum and maintained in a controlled environment (20°C±2°C, humidity 50%±10%, 12-h light/12-h dark cycle). The research protocol guiding the care and handling of laboratory animals used in this study was reviewed and approved by the Pusan National University-Institutional Animal Care and Use Committee (PNU-IACUC). The mice were divided into five groups with five individuals housed in each cage. Treatment groups were defined as follows: saline injected=0.9% NaCl solution and orally administered water; scrambled Aβ 25-35=Aβ 25-35 mice, injected=Aβ 25-35 and orally administered water; B100 mice, injected=Aβ 25-35 injection and orally administered 100 mg/kg/day of the n-BuOH fraction; B200 mice, injected=Aβ 25-35 injection and orally administered 200 mg/kg/day of the n-BuOH fraction; rutin-treated mice, injected=Aβ 25-35 injection and orally administered 100 mg/kg/day of rutin. All oral administrations were performed for 14 days using a sonde. At the start of the study, no significant differences were observed in body weight between the groups. The timeline of behavioral procedures is presented in Figure 3.
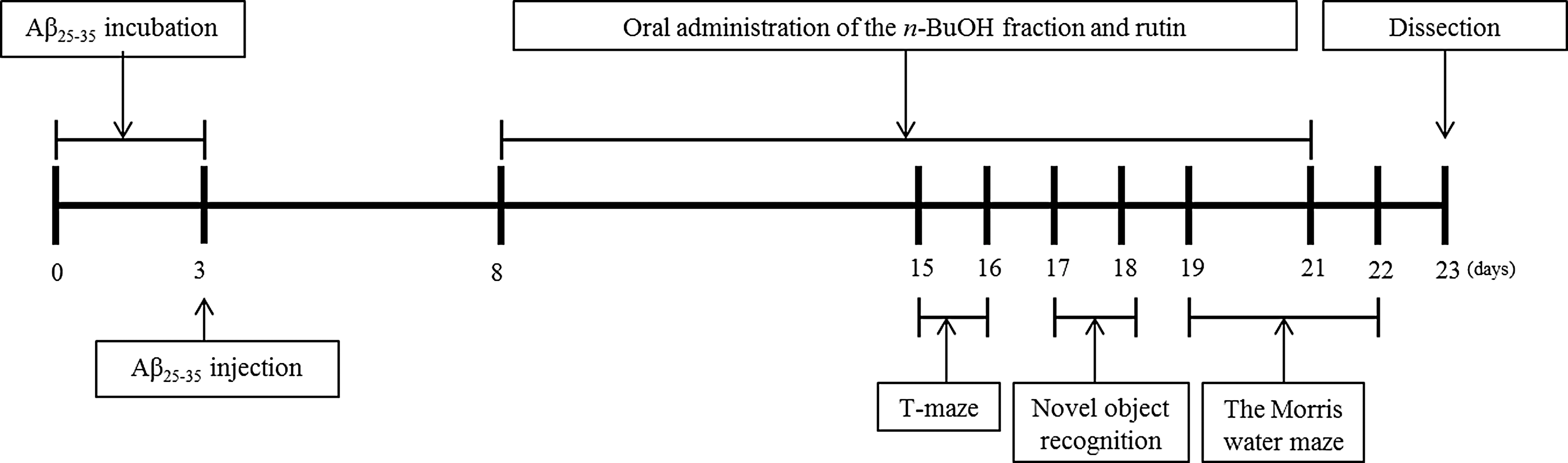
Behavioral experiment schedule for mice injected with Aβ 25-35.
Aβ25-35-infused mouse model
Aβ 25-35 was aggregated according to the procedure outlined by Maurice et al. 22 Briefly, the Aβ 25-35 peptide was dissolved and diluted in sterile distilled water to the concentration of 1 mg/mL, aliquoted into tubes, and incubated at 37°C for 3 days to achieve aggregation before injection. Aggregated Aβ 25-35 in water was injected into mice according to the procedure established by Laursen and Belknap. 23 The mice were lightly anesthetized with ether, and sterile distilled water containing Aβ 25-35 was injected directly into the third ventricle, 0.25 mm posterior to the bregma and at a depth of 2.4 mm (anteroposterior, −0.25 mm; mediolateral, 0 mm; dorsal ventral, −2.4 mm relative to the bregma). All injections were administered with a 10-μL Hamilton microsyringe, fitted with a 26-gauge needle, inserted 2.2 mm beneath the surface of the brain. The animals were injected with a 5-μL volume containing 5 nmol of Aβ 25-35 aggregate in sterile distilled water or the same volume of vehicle without Aβ 25-35 in each cerebral lateral ventricle at a rate of 1 μL/min. The needle was left in the injection site for 1 min.
T-maze test
The T-maze test was conducted according to the procedure established by Montgomery. 24 The T-shaped maze apparatus consisted of walls constructed from black boards (length of start and goal stems, 50 cm; width, 13 cm; height, 20 cm) that had been glued to a square black bottom. The maze consisted of a start box, a left arm, a right arm, and a door to separate the two sides. Mice were placed at the start box, and the number of touches and exploration times in the right arm of the T-maze were recorded during a 10-min period (training session). The mice were then placed back into the same apparatus 24 h after the training session. They were allowed to explore the right and left sides of the maze freely for 10 min, and the number of touches and exploration times were recorded (test session). Space perception was calculated as the percentage of the number of entries into the left or right arm.
Novel object recognition test
The object recognition test 25 was performed in a square, black open-field apparatus (40×30×20 cm). Two identical objects (plastic bottles) were placed at fixed distances within the square field. The mice were then placed at the center of the square field, and the number of touches of each object was recorded during a 10-min period (training session). The mice were placed back into the same field 24 h after the training session, but this time one of the objects used during the training session was replaced with a novel object (another plastic bottle). Mice were allowed to search freely for 10 min, and the number of touches was recorded (test session). Object cognitive ability, a percentage of the amount of time spent exploring any one of the two original objects (training session) or the novel object (test session), was used as a measure of cognitive function.
The Morris water maze test
The Morris water maze test was conducted according to the procedure established by Morris, 26 with slight modifications. The apparatus consisted of a dark, plastic circular pool, 80 cm in diameter, surrounded by a 40-cm-high wall and randomly divided into quadrants. White poster color was added to the pool water to make it opaque, and the water temperature was maintained at 22°C±1°C. A platform, 8 cm in diameter, was placed 1 cm below the water surface in the middle of one quadrant. The position of the platform was not changed during the training session. Four posters on the walls of the apparatus provided visual cues for navigation. Three training trials per day were conducted over 3 days. In the training trials, the mice were randomly placed in the water facing the pool wall and allowed to swim for a maximum of 60 s. The latency time required to find the platform was recorded. Mice that found the platform were allowed to rest there for 15 s. If a mouse could not find the platform within 60 s, it was guided by an experimenter to the platform and allowed to rest there for 15 s. A probe trial of the Morris water maze test was performed 1 day after the 3 days of training were completed. In the primary test, the experiment was performed in a manner identical to the training sessions. However, in the secondary test, the trial was performed without the platform. The mice were placed in the pool and they swam for 60 s looking for the platform, with the time spent in the position previously occupied by the platform recorded. In the tertiary test, the water was transparent and the number of attempts needed to reach the platform, which was visible 1 cm above the surface of the water, was counted. Occupancy of the target quadrant (%) was calculated as the percentage of time spent in the target quadrant during a 60-s trial.
Measurement of lipid peroxidation
MDA levels were measured using the method described by Ohkawa et al. 27 After completion of behavioral observations, the mice were anesthetized using ether. The brain, liver, and kidneys were immediately removed and placed on ice. The dissected tissue was homogenized in saline solution and mixed with 1% phosphoric acid and 0.67% TBA solution. After boiling for 45 min, the solution mixture was cooled in an ice bath and 2 mL of n-BuOH was added before centrifugation at 3000 rpm for 10 min. The absorbance values of the supernatant were measured at 535 and 520 nm. The level of lipid peroxidation was calculated using an MDA standard curve.
Nitric oxide scavenging activity
The nitric oxide (NO) concentration in tissues was determined using the method described by Schmidt et al.
28
An aliquot (150 μL) of the supernatant from the lipid peroxidation procedure was mixed with 130 μL of distilled water and a 20-μL sample of the dilution was added to the same amount of phosphoric acid and 0.1%
Statistical analysis
The results are expressed as mean±SD. The statistical significance was determined using a one-way ANOVA, followed by Duncan's post hoc test and Dunnett's test. Comparisons with P<.05 were deemed significant.
Results
Identification of rutin from TB
The B-ring aromatic protons exhibited a typical ABX system at δ 7.54 (d, J=2.5 Hz) for H-2′, δ 6.84 (d, J=8.5 Hz) for H-5′, and δ 7.55 (dd, J=2.5, 8.5 Hz) for H-6′ of rutin by 1 H-NMR assignments. The FAB-MS spectrum consisted of the protonated molecular ion [M+H]+ at m/z 611 for rutin (Fig. 2). 29
T-maze test
The data regarding the effect of buckwheat on new route cognition in all groups are shown in Figure 4a. The noninjected group was better at distinguishing between the familiar and the newly opened arm than the scrambled Aβ 25-35 group. However, mice treated with B100, B200, and rutin showed a greater preference for the newly opened arm of the T-maze. The B200-administered group, in particular, showed greater spatial cognition of the newly opened arm. These results demonstrate that oral administration of TB is protective against Aβ 25-35-induced impairment of spatial cognition in mice.

Spatial perceptive ability scores
Novel object recognition task
No significant differences were found between the treatment groups in cognitive recognition of the familiar object 24 h after the training session (Fig. 4b). However, Aβ 25-35-injected mice exhibited lower cognitive ability, as assessed by the novel object recognition task, in comparison with the noninjected group. Animals administered B100, B200, and rutin exhibited a significantly greater number of touches and more time spent around the novel object compared with the familiar object than the scrambled Aβ 25-35 group in the test session. In particular, rutin-treated animals showed relatively high recognition of the novel object compared with the scrambled Aβ 25-35 group, indicating that TB administration improved the recognition ability of mice impaired by Aβ 25-35 infusion.
The Morris water maze test
The results of the Morris water maze test are shown in Figures 5, 6a, 6b and Table 1. In the training session, all groups learned to reach the platform and showed a progressive reduction in the time required to reach the platform with increased training. The scrambled Aβ 25-35 group required the most time to reach the platform during the final test (Fig. 5). Moreover, the latencies of noninjected animals and B100-, B200-, and rutin-treated groups were all shorter than those recorded in the scrambled Aβ 25-35 group during the 3 days of repeat training. The shortest time to reach the platform was observed in the B200-treated group during the final test. Figure 6a shows the occupancy time of each experimental group in the zone containing the cue poster when the platform was removed. The scrambled Aβ 25-35 group spent less time in the platform quadrant than the noninjected group, while the B100-, B200-, and rutin-treated groups spent the longest time in the platform quadrant. These data suggest that the administration of B100, B200, and rutin improved the ability of mice to remember the position of the platform. The results of this exercise and visual recognition ability are presented in Figure 6b. No significant differences were seen between the experimental groups for any of the measures with a visible platform. This result supports the idea that TB protects against cognitive impairment induced by Aβ 25-35, but suggests that this role is not associated with exercise or visual ability.
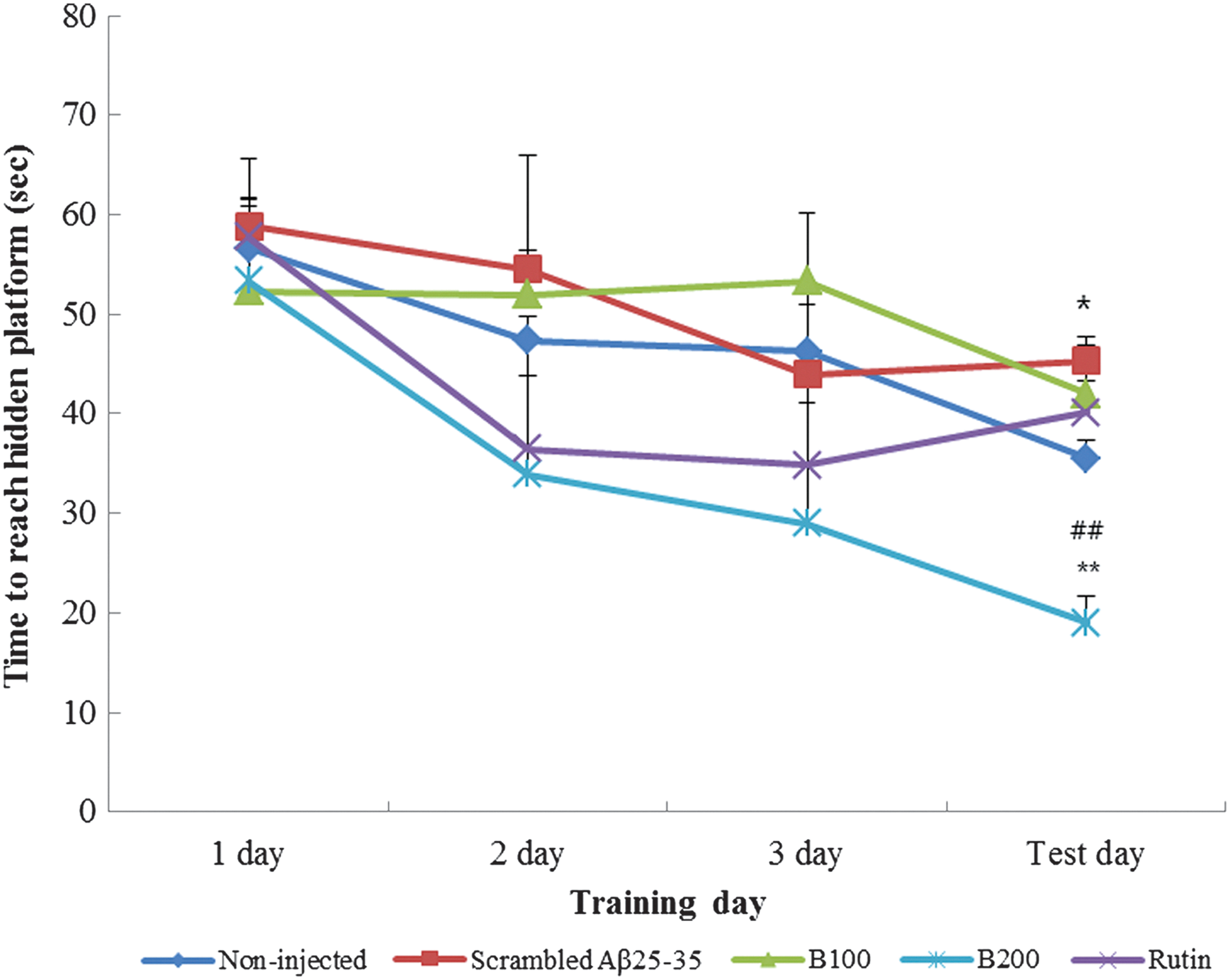
Spatial learning in the Morris water maze test. Values are reported as the mean±SD. *P<.05 showed a significant difference compared with the noninjected group, **P<.01 showed a significant difference compared with the noninjected group, and ##
P<.01 showed a significant difference compared with the scrambled Aβ
25-35 group. Color images available online at
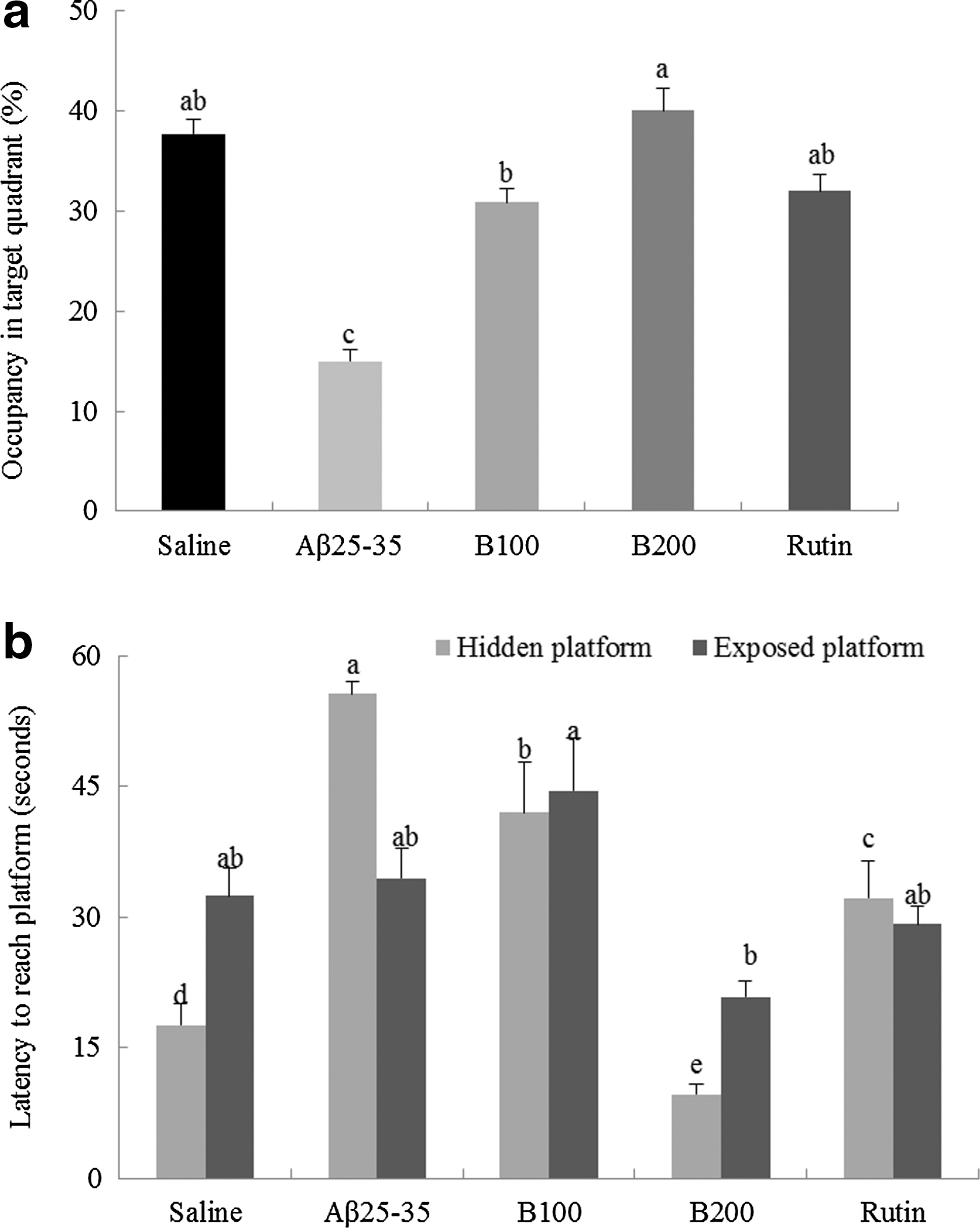
Time spent in the target quadrant
Values are reported as the mean±SD.
Scores marked with the letters a–c are significantly different (P<.05).
Mean values in the row of final test with different superscripts are significantly different as determined by one-way ANOVA, followed by Duncan's post hoc test (P<.05).
Measurement of lipid peroxidation
The inhibitory effects of B100, B200, and rutin against lipid peroxidation induced by Aβ 25-35 are shown in Figure 7a–c. MDA levels, an important index of lipid peroxidation, in the brains of the noninjected group were 66.41 nmol/mg protein, while the scrambled Aβ 25-35 group, which received Aβ 25-35 injections and oral treatment with water, exhibited increased MDA levels (92.87 nmol/mg protein). In comparison, the MDA values in the B100, B200, and rutin groups were 42.97, 68.07, and 36.3 nmol/mg protein, respectively, demonstrating that TB administration can significantly inhibit the formation of MDA in the brain. MDA concentration in the liver of the scrambled Aβ 25-35 group was 44.75 nmol/mg protein, which was 2×higher than the noninjected group. A significant decrease was also observed in the levels of MDA in the liver tissue of the B100, B200, and rutin groups with values of 18.95, 34.84, and 33.29 nmol/mg protein measured, respectively. Furthermore, MDA levels in the kidneys of the scrambled Aβ 25-35 group were 68.99 nmol/mg protein, a much higher value compared with the levels measured in the noninjected group. This value was decreased in B100, B200, and rutin groups. Overall, our results indicate that administration of TB markedly lowers MDA levels in the brain, liver, and kidney.
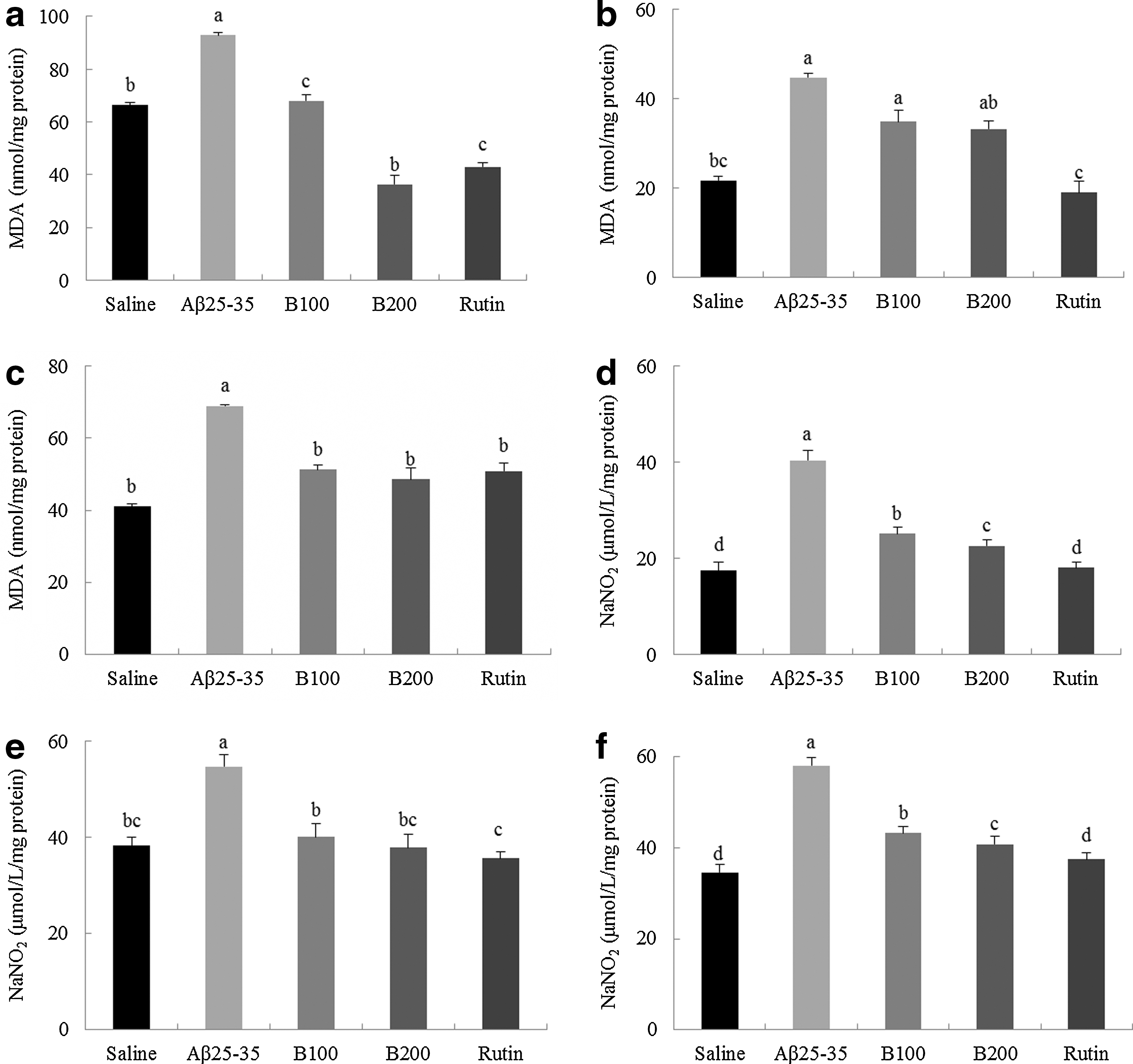
The protective activities of the n-BuOH fraction and rutin from TB on Aβ
25-35-induced lipid peroxidation and nitric oxide formation in the brain
NO scavenging activity
NO scavenging activity after the administration of B100, B200, and rutin was measured in the brain, liver, and kidney, as shown in Figure 7d–f. NO levels in the brain tissues of the noninjected group were 17.51 nmol/mg protein compared with 40.41 nmol/mg protein measured in the scrambled Aβ 25-35 group. Interestingly, animals administered rutin, B100, or B200 extract showed a significant reduction in NO production, with NO levels of 18.03, 25.21, and 22.66 nmol/mg protein observed, respectively. Furthermore, elevated NO levels in the liver were observed following the Aβ 25-35 injection. The administration of B100, B200, and rutin significantly lowered NO levels compared with the scrambled Aβ 25-35 group. Last, NO levels in the kidneys of the noninjected group were 34.52 nmol/mg protein, while those of the scrambled Aβ 25-35 group were higher, at 58.04 nmol/mg protein. The kidney NO levels in animals administered B100, B200, and rutin were lower than those measured in the scrambled Aβ 25-35 group: 37.40, 43.19, and 40.81 nmol/mg protein, respectively. Therefore, administering TB strongly inhibited NO generation in the brain, liver, and kidney.
Discussion
AD is a progressive neurodegenerative disorder diagnosed clinically by progressive deficits in cognition and memory and pathologically by an increase in Aβ and deposition of Aβ plaques in the brain. 30 Increased proteolytic degradation of Aβ precursor protein (APP) is considered among the primary causative molecular mechanisms of the disease process. 31 At excessive levels, Aβ peptides tend to oligomerize, leading to polymerization, conformational change, fibrillation, and deposition of fibrillar Aβ plaques within the brain parenchyma. 32 Aβ 25–35 induces brain inflammation, oxidative stress, activation of proapoptotic caspases, impairment of long-term potentiation, and cell loss in the hippocampus, and, most notably, leads to memory impairments. 33 Fang and Liu reported that Aβ 25-35-treated mice showed neuronal degeneration in the hippocampus and disarray in the arrangement of neurons on histological investigation of pathological damage. Additionally, immunohistochemical and biochemical analyses showed significantly increased expression of acetylcholinesterase, Bax, and Bcl-2 in the hippocampal CA1 region of Aβ 25-35-injected mice. 34
In the present study, we demonstrated the protective effects of the n-BuOH fraction and rutin from TB against cognitive dysfunction in an Aβ 25-35-induced animal model of AD. Aβ 25-35 injection resulted in a moderate, but significant, decrease in the granule cell number in all hippocampal areas from 1 week to 1 month after injection. 35 After 8 days, Aβ 25-36 injection decreased choline acetyltransferase activity in the medial septum, cortex, and hippocampus and decreased the number of choline acetyltransferase-immunoreactive cells in the medial septum. 36 Oxidative stress contributed to the in vivo Aβ-induced toxicity. At 1 week after injection, Aβ 25-35 induced significant oxidative stress in the hippocampus, as reflected by measured increases in lipid peroxidation, protein nitration, and superoxide generation. 37,38 In addition, the levels of procaspase 9, 12, and 3 in the hippocampi of mice 35 were increased at 1 week after Aβ 25-35 injection. Based on these evidences, the behavior experiment with Aβ 25-35 injected mice was scheduled.
The oral doses of fractions and rutin were based on a previous study. We observed that 14 days of daily administration of TB MeOH extract (100 mg/kg) before training resulted in increased memory in an AD model. 39 Furthermore, Puchchakayala and coworker reported that rutin is safe, at repeated intakes, in doses less than 2000 and 100 mg/kg/day, doses found to be more effective than 50 mg/kg dose. 40 On the basis of these studies, we determined the experimental schedule and oral dose. We carried out behavioral assessments using the objective recognition, T-maze, and the Morris water maze tests. Moreover, inhibitory effects on lipid peroxidation, as well as NO scavenging activity, were also investigated.
The administration of the n-BuOH fraction and rutin from TB significantly alleviated Aβ 25-35-induced cognitive impairments, as demonstrated by the object recognition test, T-maze test, and the Morris water maze test. In the novel object recognition test, the exploration times of a previously introduced object and a novel object were measured as indices of memory performance. 41 The results showed that scores in novel object recognition and new route awareness tests were higher in groups who were administered the n-BuOH fraction and rutin from TB than in animals in the scrambled Aβ 25-35 group, suggesting an improvement in cognitive ability. In particular, rutin-treated animals showed the highest index among the treatment groups. The results of the behavioral tests indicate that TB strongly protects against Aβ 25-35-induced impairments of learning and memory function in mice. The T-maze test is used to evaluate the long-term latent learning of mice. 42 Animals administered the n-BuOH fraction and rutin from TB showed higher cognitive ability, manifested by increased recognition of the new route, with the B200-treated animals displaying the highest cognitive recognition ability.
Spatial cognitive ability and long-term memory were assessed using the Morris water maze test. In training trials, the latency times of mice treated with the n-BuOH fraction and rutin from TB were significantly shortened by repeated training for 3 days, while the latency of the mice in the scrambled Aβ 25-35 group was not reduced by training. In mice administered B200, in particular, the latency time required to find the platform was significantly shortened. Compared with the scrambled Aβ 25-35 group, animals treated with the n-BuOH fraction and rutin from TB needed less time to reach the platform and spent more time at the platform position, indicating improvements in memory. However, the time needed to reach the exposed platform was not significantly different between the treatment groups (P<.05). These results suggest that deficits in long-term spatial cognition induced by Aβ 25-35 are not related to swimming or visual ability and thus receive no protective effects from the n-BuOH fraction and rutin from TB. Taken together, our results indicate that the n-BuOH fraction and rutin from TB prevented the impairments in short-term and recognition memory induced by Aβ 25-35. B200 and rutin treatments, in particular, exhibited stronger protective activity against Aβ 25-35-induced impairments in memory and cognition.
Lipid peroxidation, a well-known marker of oxidative stress, is widespread in the brains of AD patients. 43 Indicators of oxidative stress include urinary 8-iso-prostaglandin F2α formation and MDA. MDA is one of a number of breakdown products produced during lipid peroxidation 44 and has traditionally been used as a measure of oxidative stress. The specificity of this compound as a marker of lipid peroxidation is somewhat dubious 45 with a number of other biological processes yielding MDA as an end product. 46 Therefore, levels of 8-iso-prostaglandin F2α , a bioactive isoprostane produced from arachidonic acid through the predominantly nonenzymatic process of membrane lipid and low density lipoprotein oxidation, 47 were also used for indicators of lipid peroxidation. Boyle et al. 48 carried out quantitation of urinary 8-iso-prostaglandin F2α following 6 weeks of rutin supplementation. Although there was no significant change in the level of urinary 8-iso-prostaglandin F2α following 6 weeks of rutin supplementation, a significant positive correlation was found between the urinary e8-iso-prostaglandin F2α and MDA. Although we did not measure the secretion of 8-iso-prostaglandin F2α , a decline in MDA levels by rutin suggests the inhibition of lipid peroxidation.
The injection of Aβ 25-35 into the brains of mice led to notable increases in the MDA content in brain tissue, indicating that Aβ 25-35 injection increases lipid peroxidation. To confirm the effects of the n-BuOH fraction and rutin from TB on Aβ 25-35-induced oxidative stress, we also measured the levels of NO. We found that treatment with n-BuOH fraction and rutin from TB prevented the accumulation of MDA and NO induced by Aβ 25-35. B200 and rutin, in particular, showed a significant decrease in both lipid peroxidation, evidenced by reduced MDA levels, and NO production. These results suggest that the protective effects of B200 and rutin against Aβ 25-35-induced memory impairment are related to the attenuation of oxidative stress. The toxicity of NO increases when it reacts with O2 to generate peroxynitrite, which is a strong oxidant that can decompose to produce •OH and NO. 49 Peroxynitrite is known to produce compounds such as nitrotyrosine. The accumulation of nitrotyrosine correlates with increased levels of cerebral Aβ and the severity of cognitive impairment. NO-mediated oxidative damage was confirmed to contribute to Aβ-induced neuronal toxicity and cognitive deficits, while the overproduction of NO in the brain contributes to the pathological condition of AD. 50 The increased NO synthesis and superoxide generation could result in the formation of peroxynitrite and nitration of protein tyrosine residues, 49 suggesting that rutin might inhibit the nitration of tyrosine.
The age-dependent progression of neurodegeneration is associated with decreased antioxidant capacity and increased oxidative damage to proteins, DNA, and lipids. 51 In animal models, expression of antioxidant enzymes, such as superoxide dismutase (SOD), catalase, and glutathione peroxidase (GSH-Px), as well as lipid peroxidation levels and NO concentration in the blood and brain, can be substantively altered in patients with AD. The brain contains high levels of polyunsaturated fatty acids, which are targets for the initiation of lipid peroxidation and can propagate the formation of reactive lipid species such as MDA, which in turn are capable of modifying proteins. 52 To protect the biological systems from free radical toxicity, several cellular antioxidant defense mechanisms keep the production of reactive oxygen species (ROS) in check, including both enzymatic and nonenzymatic pathways. 53 The primary antioxidant enzymes involved in scavenging superoxide radicals include SOD, catalase, and GSH-Px. These enzymes act together in the metabolic pathway of free radicals, and altered activity of one enzyme without compensatory changes in others may lead to lipid peroxidation. In the brain, decreased antioxidative system capacity has been reported in aging and AD. 54 ROS can be scavenged by endogenous antioxidants, including SOD and GSH-Px. Wang et al. showed that TB regulates serum antioxidant enzymes such as SOD and GSH-Px and inhibits the formation of peroxides and free radicals. These findings indicate that the ability of TB to inhibit lipid peroxidation and NO production may be a result of the elevation in the antioxidative enzyme defense system. 55 Decreased NO production, in particular, may be related to the elevation of antioxidative activities of the n-BuOH fraction and rutin from TB, which compete with oxygen to react with NO and thereby inhibit the generation of nitrite. Phenolic compounds and flavonoids have been associated with antioxidative action in biological systems, acting as scavengers of NO. 56,57
The results of the present study demonstrate, for the first time, that TB can alleviate Aβ 25-35-induced memory deficits in mice. Our findings suggest that the prevention of oxidative damage in the brain by TB may be attributed to the n-BuOH fraction and rutin components. As a therapeutic agent, TB is well tolerated, largely free of adverse effects, and has few adverse drug interactions. 58 Aβ-peptide is produced by a number of different cell types and circulates in blood and cerebrospinal fluid in its soluble form. 59 Senile plaques are defined by the progressive deposition of Aβ in the parenchyma and cerebral microvasculature, which is a characteristic event in the pathogenesis of AD. 60 Increased levels of oxidative stress are associated with amyloid plaques and neuronal degeneration in the brains of AD patients, as well as with their alterations in peripheral tissues or circulating cells. 61 Additionally, the concentration of reactive oxygen species and levels of lipid peroxidation in the cortex, hippocampus, spleen, and astrocytes are known to be significantly elevated in Aβ 25-35-injected mice. This study demonstrated that the induction of oxidative stress by Aβ was also present in peripheral tissues in addition to the brain.
Although multiple mechanisms have been hypothesized for Aβ peptide-induced memory deficits, there is considerable evidence suggesting that Aβ-induced disruption of kinases such as ERK is critical for memory impairment. 9 Aβ could inhibit the activation of ERK and CREB in primary neurons and thereby suppress the downstream expression of postsynaptic N-Methyl-D-aspartate receptor subunits. 62 The critical role of CREB was established in memory, both in long-term consolidation and indirect regulation of short-term memory. 63 Decreased phosphorylation of CREB was observed in postmortem brains of AD patients. Additionally, lower BDNF levels were found in AD, with the magnitude of decrease correlating with the loss of cognitive function. 64 Moghbelinejad et al. reported that rutin could activate the ERK-CREB pathway, and the activation of this cascade leads to increased expression and release of BDNF from the synapse through enhanced CREB activation. 65 Based on this evidence, upregulation of ERK1, CREB, and BDNF expression by rutin treatment may underlie the improvements in memory and defend against Aβ neurotoxicity.
The mechanism of how rutin decreased Aβ accumulation by suppressing the expression of APP and Aβ remains unclear, but several reports have demonstrated that rutin could activate the ERK-CREB pathway and activation of this cascade leads to increased expression and release of BDNF from the synapse through enhanced CREB activation. Thus, it seems that upregulation of ERK1, CREB, and BDNF expression by rutin could improve memory and defend against Aβ neurotoxicity. 65 Rutin showed antiamyloidogenic effects in vitro by reversibly binding to the amyloid fibril structure of Aβ oligomer and monomers.
Several past studies have demonstrated that buckwheat contains antioxidants, such as rutin, tocopherols, and phenolic acids, and can be stored for a long time without apparent chemical changes. 66 Rutin content in buckwheat has been documented to be the highest of all tested plants. In a previous study, rutin was isolated from TB and shown to exhibit beneficial biological activities. 67 Rutin is a flavonol glycoside plant metabolite with antioxidative, anti-inflammatory, and anticarcinogenic effects, which can also reduce the fragility of blood vessels related to hemorrhagic disease and hypertension in humans. 68 In addition, Koda et al. 69 demonstrated the protective effects of rutin against cognitive impairment in normal rats. They reported that rats treated with rutin showed the highest spatial memory ability in the Morris water maze test. In our present study, rutin exerted a protective activity against the impairment of cognitive function and memory in AD mice. Based upon this evidence, we suggest that rutin may not only improve cognitive ability under AD pathological conditions but also under normal conditions as well. Several studies have demonstrated the antioxidative activity of buckwheat hull extract against oxidative stress and rutin against Aβ-induced neurotoxicity. 70,71 Koda et al. 69 reported protective effects of rutin against spatial memory impairment induced by trimethyltin in rats, and Javed et al. 71 reported protective effects of rutin for neuroinflammation in a rat model of sporadic dementia. However, the protective activity of the fraction and rutin from TB under the Aβ-induced AD model has not been clearly studied, and this is the first report on the study that n-butanol fraction and rutin from TB improve cognition and memory impaired by Aβ. In addition, the present study is significant and important because the active fraction from TB and active rutin isolated and identified from TB, that contains higher amounts of rutin than common buckwheat, protected from Aβ-induced AD. TB is a functional food material that contains proteins with high biological value and balanced amino acid composition, relatively abundant crude fiber, vitamins B1, B2, and B6, and higher rutin content than common buckwheat. The seeds of common buckwheat contain rutin at levels between 12.6 and 35.9 mg/100 g, 19 while rutin content in TB seeds is between 800 and 1700 mg/100 g. 20 These findings imply that the protective role of buckwheat may be related to the quantity of rutin present. Accordingly, TB may provide more protection than common buckwheat due to its higher rutin content. The present study demonstrated that the n-BuOH fraction and rutin from TB can protect memory and cognitive function against Aβ 25-35-induced oxidative stress. Based on our research, adding TB to the diet could be particularly beneficial to AD patients.
Footnotes
Acknowledgments
This research was supported by the Chung-Ang University Research Scholarship Grants in 2014. The authors thank the staff and crew of the National Center for Inter-University Research Facilities (Seoul National University) for assistance with the NMR and GC/MS experiments.
Author Disclosure Statement
No competing financial interests exist.