
Abstract
Epidermal hydration is maintained by the epidermal lipid barrier, of which ceramide (Cer) is the major constituent. We examined the dietary effect of royal jelly (RJ) on epidermal hydration in aged mice. Altered Cer metabolism was further determined by measuring epidermal levels of individual Cer, glucosylceramide (GC), and sphingomyelin (SM) species, and of Cer-metabolizing enzymes. Aged C57BL/6J mice were fed a control diet (group AGED) or diets with 1% RJ harvested from two different areas (groups AGED+RJ1:AGED+RJ2) for 16 weeks. Aged C57BL/6J mice with no dietary intervention (the control group: group C) represented the onset of aging. In group AGED, epidermal levels of hydration, Cer1/2/5/6/7, GC-A/B/C/D, SM1/2/3, and β-glucocerebrosidase (GCase) protein, an enzyme of GC hydrolysis for Cer generation, were lower than in group C; these levels, as well as those of Cer3/4 and acidic sphingomyelinase (aSMase) protein, an enzyme of SM hydrolysis for Cer generation, were higher in group AGED+RJ1 than in group AGED. Despite increases in GC-B, SM1/2/3, and serine palmitoyltransferase2 protein, an enzyme of de novo Cer synthesis, in group AGED+RJ2 to levels higher than in group AGED, epidermal levels of hydration, Cer1-7, GC-A/C/D, GCase, and aSMase proteins were similar in these two groups. Expression of GCase and aSMase mRNAs, and of Cer synthase3 and ceramidase proteins, enzymes of de novo Cer synthesis and degradation, did not differ among groups. Dietary RJ1 improved epidermal hydration by enhancing Cer metabolism with increased levels of all Cer, GC, and SM species, and of GCase and aSMase proteins.
Introduction
S
Skin dryness in aged skin, 3 –5 coupled with the inconvenience of frequently applying moisturizer, has led to the development of alternative approaches using dietary and natural substances. 6 –8 Royal jelly (RJ), a yellowish material secreted from the hypopharyngeal and mandibular glands of worker-caste (nurse) honeybees, has been reported to have beneficial effects on skin dryness. 9 –11 Topical application of (E)-10-hydroxy-2-decenoic acid (10-HDA), the major bioactive substance of RJ, enhances epidermal hydration by restoring skin barrier function in UV-irradiated human subjects. 9 Furthermore, our previous studies indicate that dietary supplementation with RJ improves epidermal hydration by upregulating the generation of Cer from glucosylceramide (GC) and sphingomyelin (SM), two major precursor lipids, 12,13 in UV-irradiated hairless mice. 10,11 RJ has a beneficial effect on skin protection, based mainly on the observation of skin dryness in extrinsic aging. However, there is limited research evaluating the effects of RJ on skin dryness due to intrinsic aging. In this study, we examined the dietary effect of RJ on epidermal hydration in aged C57BL/6J mice, an animal model of intrinsic aging. 14,15 To partially delineate a possible mechanism for alteration of Cer metabolism, epidermal levels of individual Cer, GC, and SM species, as well as protein expression of Cer-metabolizing enzymes involved in de novo Cer synthesis, GC and SM hydrolysis, and Cer degradation processes, were further determined.
Materials and Methods
Materials
Lipid standards for Cer2 (860524), Cer3 (860724), Cer5 (860823), Cer6 (860924), GC-B (860549), SM1 (860592), and SM2 (860584) were purchased from Avanti Polar Lipids, Inc. (Alabaster, AL, USA). Antibodies against serine palmitoyltransferase 2 (SPT2; SC27500), ceramide synthase 3 (CerS3; SC55962), acidic sphingomyelinase (aSMase; SC9817), ceramidase (CDase; SC28486), and β-actin (SC1616) were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). An antibody against β-glucocerebrosidase (GCase; ab92997) was purchased from Abcam (Cambridge, MA, USA). High-performance thin layer chromatography (HPTLC) plates (Silica gel 60) were obtained from Merck (Darmstadt, Germany). All other chemicals and solvents used were of the highest analytical grade.
RJ preparation
Fresh RJ1 and RJ2 were harvested from the Cheorwon (RJ1) and Pocheon (RJ2) areas in the Republic of Korea in June. Voucher specimens (RJ1: KHUKSY-HPOP001, RJ2: KHUKSY-HOOC001) were deposited in the herbarium at the Graduate School of East-West Medical Science, Kyung Hee University (Yongin, Republic of Korea).
Animals and diet
Female C57BL/6J mice aged 6 months (24 weeks) and weighing 25–30 g were purchased from Japan SLC (Haruno, Japan). In this study, intrinsic aging was limited to middle age before menopause, when alterations in aging-related metabolic and signaling pathways are not affected by gender. 1,2 Because aggression in male mice causes skin wounds when they are group housed, we used 6-month-old aged female C57BL/6J mice (murine counterparts of 30-year-old human subjects). 16 After a 1-week adaptation period, mice were assigned to one of three groups of seven mice each: an intrinsically aged group (group AGED) fed a control diet, and groups AGED+RJ1 and AGED+RJ2 fed a diet supplemented with 1.0% RJ1 or RJ2, respectively, for 16 weeks. Since the typical upper age limit for mature C57BL/6J mice is∼6 months and past this age, mice might exhibit some age-related changes, 16 6-month-old aged female C57BL/6J mice with no dietary intervention (control group: group C) (n=5) were used to represent the onset of age-related skin changes. Compositions of experimental diets are shown in Table 1.
Aged female C57BL/6J mice were fed a control diet (group AGED) or diets supplemented with 1% royal jelly (RJ) harvested from area 1 (group AGED+RJ1) or area 2 (AGED+RJ2) for 16 weeks. Aged female C57BL/6J mice with no dietary intervention (control group: group C) represented the onset of aging.
Diet composition was based on the AIN-93G diet for mice. 18
AIN-93 vitamin mix 310025 (Dyets, Inc., Bethlehem, PA, USA).
AIN-93G salt mix 210025 (Dyets, Inc., Bethlehem, PA, USA).
Mice were group housed with two or three mice per cage in plastic cages under controlled temperature (22–24°C), humidity (55–60%), and light (lights on from 07:00 to 19:00). Mice were given ad libitum access to food and water. Weight gain and food intake were monitored weekly. Animal care and handling protocols were approved by the Animal Care and Use Review Committee of Kyung Hee University (KHUASP-12-010). At the end of week 16, mice were sacrificed by cervical dislocation, and epidermal strips of the dorsal area were removed as described previously. 17
Assessment of epidermal hydration
Hair from the dorsum of the mice was shaved and depilated with sodium thioglycollate (Niclean cream; Ildong Pharmaceutical Co., Seoul, Korea). Under standardized conditions of 22–24°C and 55–60% humidity, epidermal hydration was measured on the dorsal skin surface using a Corneometer (model MPA-5; Courage+Khazaka Electronic GmbH, Cologne, Germany). 17 The value of skin hydration recorded for each mouse was the average of at least five determinations after equilibrium was attained. The temperature and humidity of the probe were also recorded. Data are expressed as capacitance in arbitrary units.
Analysis of individual Cer, GC, and SM species
Epidermal strips were homogenized, and the protein concentration was measured in epidermal homogenates as described previously. 17 The rest of the epidermal homogenate was extracted with chloroform (CHCl3):methanol (MeOH) (2:1, v/v) to obtain total lipids, and individual Cer, GC, and SM species were fractionated by HPTLC with solvent systems using a Linomat 5 autosampler (CAMAG, Muttenz, Switzerland) as described previously. 18 Fractions containing Cer, GC, and SM species, which had comigrated with the respective standards, were treated with cupric acetate–phosphoric acid, heated to 160°C for 15 min, and scanned using a TLC III scanner (CAMAG). 18 For identification of individual Cer, GC, and SM fractions, Cer1-7, GC-A/B/C/D, and SM1-3 isolated from mouse epidermis (kindly provided by Dr. Y. Uchida, Department of Dermatology, University of California-San Francisco) were further used. Levels of Cer, GC, and SM species were quantified using calibration curves generated with various concentrations of each respective standard and were expressed as μg/μg protein.
Western blotting
Protein extracts (35 μg/each) from the epidermis were separated electrophoretically on 8–10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then blotted on nitrocellulose membranes, as described previously. 18 Membranes were incubated with primary antibodies against SPT2 (70 kDa), CerS3 (50 kDa), GCase (65 kDa), aSMase (60 kDa), CDase (55 kDa), and β-actin (42 kDa), followed by incubation with immunoglobulin G-horseradish peroxidase secondary antibodies. Immunoreactive proteins were visualized using the ECL detection system (GE Healthcare, Buckinghamshire, United Kingdom), and band density was quantified by densitometry.
Quantitative reverse transcription–polymerase chain reaction of GCase and aSMase mRNAs
Total RNA was extracted from the epidermis using the TRIzol reagent (Gibco, New York, NY, USA). The isolated RNA was quantified and electrophoresed in 1.2% agarose gels to assess the sample quality, as described previously. 19
Reverse transcription of isolated total RNA (5 μg) was performed using a SuperScript III first-strand synthesis system (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Quantitative real-time PCR was performed in 384-well plates using the Thermal Cycler Dice Real Time System TP850 (Takara Bio, Inc., Shiga, Japan). Each 25 μL reaction mixture consisted of 12.5 μL of SYBR® Premix Ex Taq (Takara Bio, Inc.), 2 μL of diluted cDNA, and 0.5 μL of 10 μM of forward and reverse primers for GCase, aSMase, or GAPDH. The following primer sets were used: GCase, 5′-GTG ACT TCT CCA TCC GTG TCT-3′ (forward), 5′-CGT AGG TTC ATT CTC CGC TGT-3′ (reverse) 20 ; aSMase, 5′-CAC GTG GAT GAG TTT GAG GT-3′ (forward), 5′-AGA GCT CCC AGA GTA GTT AC-3′ (reverse) 21 ; and GAPDH, 5′-AAT GGT GAA GGT CGG TGT GA-3′ (forward), 5′-CTG GAA GAT GGT GAT GGG C-3′) (reverse). 22 Thermal cycling was initiated at 95°C for 10 s to activate the polymerase, followed by 40 cycles of 95° for 5 s, 60°C for 30 s, and 72°C for 30 s.
Statistical analysis
Data are expressed as mean±standard error of the mean. All data were analyzed by one-way analysis of variance using SPSS for windows (SPSS 18.0; SPSS, Inc., Chicago, IL, USA), and differences among groups were determined by Tukey's honestly significant difference post hoc test for unequal sample size. Differences with P<.05 were considered significant.
Results
Epidermal hydration
Weight gain and food intake of groups AGED+RJ1 and AGED+RJ2 did not differ from those of groups C or AGED (Table 2). As there were no differences in weight gain and food intake, the food efficiency ratio did not differ among groups.
Aged female C57BL/6J mice were fed a control diet (group AGED) or diets supplemented with 1% RJ harvested from area 1 (group AGED+RJ1) or area 2 (AGED+RJ2) for 16 weeks. Aged female C57BL/6J mice with no dietary intervention (control group: group C) represented the onset of aging.
Values are mean±SEM (group C: n=5; groups AGED, AGED+RJ1, and AGED+RJ2: n=7). aSuperscripts with different letters in the same row indicate significant difference (P<.05) using one-way ANOVA and Tukey's honestly significant difference (HSD) post hoc test for unequal sample size.
Food efficiency ratio (FER)=weight gain (g)/food intake (g).
SEM, standard error of the mean; ANOVA, analysis of variance.
Assessment of epidermal hydration in group AGED revealed that intrinsic aging caused skin dryness; epidermal hydration (water content) in group AGED was lower than that in group C (Fig. 1). Epidermal hydration was higher in group AGED+RJ1 than in group AGED and was increased to a level higher than in group C. However, epidermal hydration of group AGED+RJ2 was similar to that of group AGED and modestly lower than that in group C, but much lower than that in group AGED+RJ1. These results suggest that RJ2 had no beneficial effects on epidermal hydration. Dietary supplementation of RJ1 improved epidermal hydration to a level higher than that in group C.

Effect of intrinsic aging and dietary royal jelly (RJ) supplementation on epidermal hydration of groups. Aged female C57BL/6J mice were fed a control diet (group AGED) or a diet with 1% RJ harvested from area 1 (group AGED+RJ1) or area 2 (group AGED+RJ2) for 16 weeks. Aged female C57BL/6J mice with no dietary intervention (control group: group C) represented the onset of aging. Epidermal hydration was measured on the dorsal skin surface at the end of week 16 using a Corneometer. Data are mean±standard error of the mean (SEM, group C: n=5; groups AGED, AGED+RJ1, and AGED+RJ2: n=7). abcSuperscripts with different letters indicate significant difference (P<.05) using one-way analysis of variance (ANOVA) and Tukey's honestly significant difference (HSD) post hoc test for unequal sample size.
Epidermal levels of Cer species
Total Cer in group AGED was lower than that in group C (Table 3). Total Cer in group AGED+RJ1 was higher than that in group AGED and similar to that in group C. However, total Cer in group AGED+RJ2 was similar to that in group AGED.
Individual species of Cer1-7, GC-A/B/C/D, or SM1-3 were fractionated by high-performance thin layer chromatography (HPTLC): Cer and GC species were first fractionated using CHCl3:MeOH:water (40:10:1, v/v/v) to 2 cm and again to 5 cm, followed by fractionation with CHCl3:MeOH:acetic acid (47:2:05, v/v/v) to 8.5 cm and finally with n-hexane:diethyl ether:acetic acid (30:15:0.5, v/v/v) to the top; SM species were fractionated using CHCl3:MeOH:acetic acid: water (25:15:4:2, v/v/v).
Aged female C57BL/6J mice were fed a control diet (group AGED) or diets supplemented with 1% RJ harvested from area 1 (group AGED+RJ1) or area 2 (AGED+RJ2) for 16 weeks. Aged female C57BL/6J mice with no dietary intervention (control group: group C) represented the onset of aging.
Data are mean±SEM (group C: n=5; groups AGED, AGED+RJ1, and AGED+RJ2: n=7). abcSuperscripts with different letters in the same row indicate significant difference (P<.05) using one-way ANOVA and Tukey's HSD post hoc test for unequal sample size.
Based on structural moieties of amide-linked nonhydroxy acid, α-hydroxy acid, ω-hydroxy acid, or ester-linked FAs on sphingoid bases, the Cer in the murine epidermis comprised a heterogeneous family of at least seven species (Cer 1-7) with different polarities. 10,13 Further analysis of individual Cer species revealed that Cer1, the least polar Cer species, 23 and the more polar Cer species such as Cer 2-7 (Cer7 is the most polar Cer among Cer1-7) were separated in the epidermis of mice, as reported previously. 10,13 Levels of Cer2, the major Cer species in the epidermis, 24 Cer1, and Cer5/6/7 were significantly lower in group AGED than in group C. In group AGED+RJ1, levels of Cer1, Cer2, and Cer 5/6/7 were significantly higher than in group AGED and similar to those in group C, which reflected the increase in total Cer. Consistent with the epidermal hydration and total Cer levels, epidermal levels of Cer1, Cer2, and Cer5/6/7 in group AGED+RJ2 were similar to those in group AGED. Although levels of Cer3/4 was similar between groups C and AGED, the level in group AGED+RJ1 (but not in group AGED+RJ2) was higher than that in groups C and AGED.
Epidermal levels of GC and SM species
Since Cer species are generated mostly from GC and SM, 12,13,24 altered levels of GC and SM are related to Cer metabolism. Total GC in group AGED was significantly lower than that in group C (Table 3). In contrast, total GC in group AGED+RJ1 was higher than that in group AGED and was similar to that in group C. Further fractionation into GC-A (the least polar GC) and more polar GC species such as GC-B, GC-C, and GC-D 12 revealed that epidermal levels of these four GC species in group AGED were significantly lower than those in group C, whereas the levels of GC-A, GC-C, and GC-D in group AGED+RJ1 were similar to or higher than those in group C. However, epidermal levels of total GC and of GC-A, GC-C, and GC-D species in group AGED+RJ2 were similar to those in group AGED. Epidermal levels of GC-B in groups AGED+RJ1 and AGED+RJ2 were higher than those in group AGED, but lower than those in group C.
SM was separated into three major subfractions (SM1/SM2/SM3, of which SM3 is the most polar). 13 Total SM, including all three SM species in group AGED, were significantly lower than those in group C (Table 3). Notably, these levels in group AGED+RJ2 were higher than those in group AGED and similar to or higher than those in group C, whereas these levels in group AGED+RJ1 were higher than those in group AGED, but lower than in group AGED+RJ2.
Protein and mRNA expressions of Cer-metabolizing enzymes in the epidermis
Cer is synthesized de novo by enzymatic condensation of serine and palmitoyl-Co A by SPT and subsequent N-acylation by CerS. 24 The newly synthesized Cer is modified at the 1-hydroxy position to either GC by GC synthase or to SM by SM synthase. Cer1-7 are generated primarily from GC hydrolysis by GCase. 12,24 Cer2 or Cer5 is generated, in part, from SM hydrolysis by SMase. 13,24 Ultimately, Cer undergoes degradation by CDase into sphingosine and FAs. 24
In the epidermis of group AGED, protein expression of GCase was significantly lower than that in group C, whereas protein expression of aSMase, which plays a greater role in maintaining the epidermal lipid barrier than other SMases, 24 –26 was similar to that in group C (Fig. 2). In group AGED+RJ1, protein expression of GCase and aSMase was higher than in group AGED. There were no apparent alterations in protein expression of SPT2, the SPT subunit of which mRNA expression is altered by UV irradiation and is required for recovery of the disrupted epidermal barrier, 27,28 in groups C, AGED, and AGED+RJ1. Notably, in group AGED+RJ2, SPT2 protein expression was higher than in group AGED, but protein expression of GCase was similar to that in group AGED and that of aSMase was modestly lower than that in group AGED+RJ1. Protein expression of CDase and CerS3, the CerS isozyme responsible for maintaining the epidermal lipid barrier with long chain fatty acylation, 29,30 did not differ among groups.
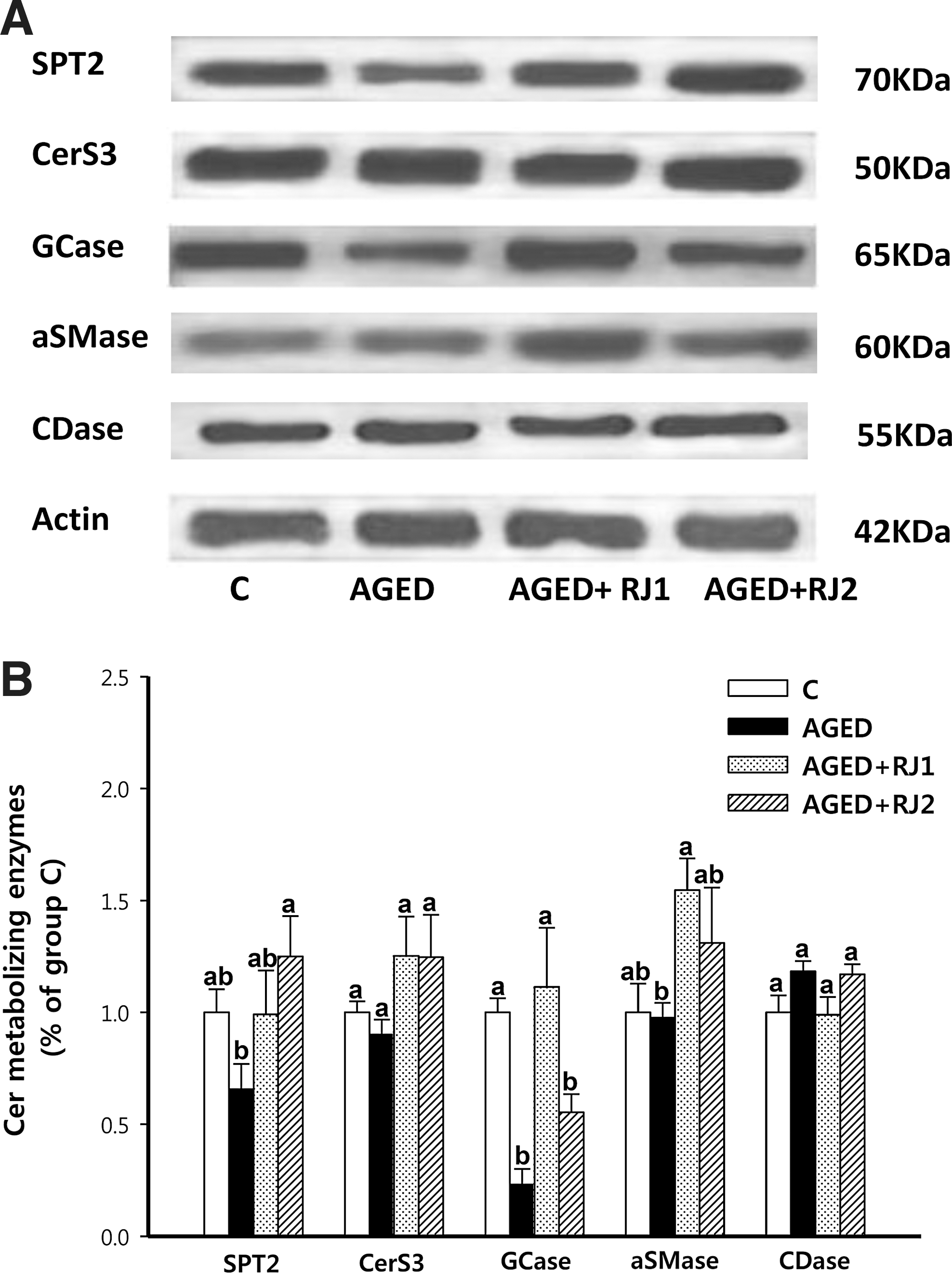
Effect of intrinsic aging and dietary RJ supplementation on protein expression of ceramide-metabolizing enzymes in the epidermis of groups. Aged female C57BL/6J mice were fed a control diet (group AGED) or a diet with 1% RJ harvested from area 1 (group AGED+RJ1) or area 2 (group AGED+RJ2) for 16 weeks. Aged female C57BL/6J mice with no dietary intervention (control group: group C) represented the onset of aging.
There were no alterations in the mRNA levels of GCase among any of these groups despite the significant difference in GCase protein levels among groups C, AGED, AGED+RJ1, and AGED+RJ2 (Fig. 3). Furthermore, the mRNA levels of aSMase did not differ among groups. Whether the translation of GCase and aSMase mRNAs or the stability of these two proteins is altered by either intrinsic aging or dietary supplementation with RJ requires further investigation.
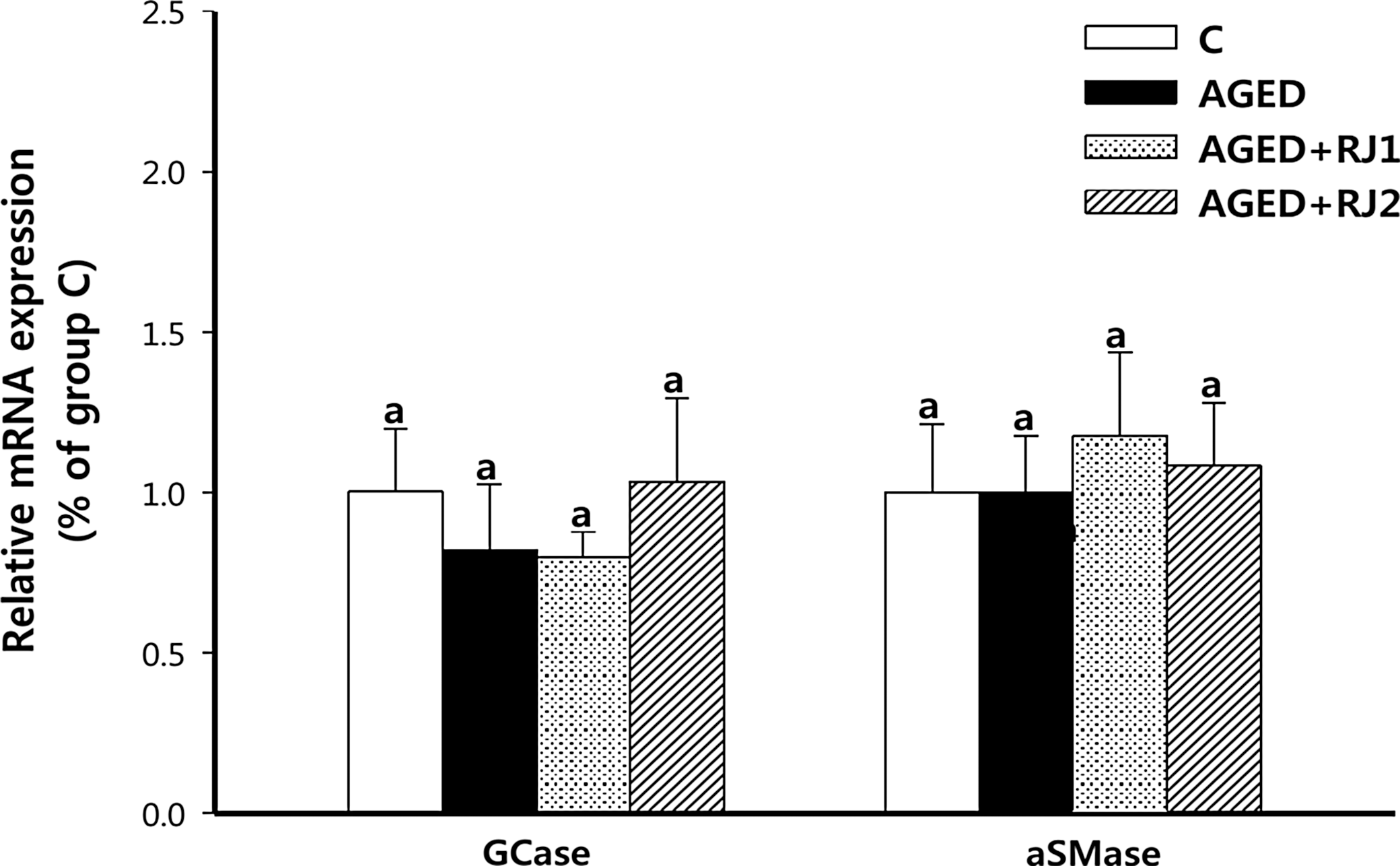
Effect of intrinsic aging and dietary RJ supplementation on mRNA expression of GCase and aSMase in the epidermis of groups. Aged female C57BL/6J mice were fed a control diet (group AGED) or a diet with 1% RJ harvested from area 1 (group AGED+RJ1) or area 2 (group AGED+RJ2) for 16 weeks. Aged female C57BL/6J mice with no dietary intervention (control group: group C) represented the onset of aging. Total RNA of the epidermis was isolated, and quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was performed for GCase and aSMase mRNAs. The signal intensities from qRT-PCR analysis were quantified and normalized, first to the corresponding values for the GAPDH internal controls and then to that of group C. Data are mean±SEM (n=5). aSuperscripts with different alphabet letters indicate significant difference (P<.05) using one-way ANOVA and Tukey's HSD post hoc test.
Discussion
Skin dryness paralleled decreased levels of total Cer, GC, and SM in group AGED. Decreased levels of Cer species in group AGED might be related to alterations in de novo Cer synthesis, GC and SM hydrolysis, and/or Cer degradation. Reduced CerS and SMase activities in 15- to 18-month-old (60 to 72 week old) hairless mice 31 and enhanced CDase activity in 41- to 84-year-old healthy humans (average age 59.5) 32 have been reported previously. However, unlike the more elderly mice or humans, 31,32 no altered expressions of SPT, CerS3, aSMase, and CDase proteins were observed in the 24-week-old (group C) to 40-week-old mice (group AGED) (murine counterparts of 30- to 38-year-old human subjects). 16 On the other hand, the decreased levels of total Cer, including Cer1, Cer2, and Cer5/6/7, paralleled the reduced levels of all GC species (GC-A/B/C/D) with low expression of GCase protein. These data, together with a previous report demonstrating that decreased GCase activity alone is able to account for epidermal barrier defects in aged mice, 33 suggest that GC hydrolysis is the primary metabolic target pathway for decreased Cer levels in middle-aged mice.
The restoration of total Cer levels in the hydrated epidermis of group AGED+RJ1 could be explained by increased protein expression of GCase. In fact, levels of total Cer, including all Cer species (Cer1-7) and protein expression of GCase in group AGED+RJ1, were similar to or higher than those in group C, which may result in GC depletion. However, levels of total GC and most GC species in group AGED+RJ1 were similar to or higher than those in group C. In addition, levels of total SM, including all SM species, and of aSMase protein expression in group AGED+RJ1 were higher than in group AGED, but there were no alterations in protein expression of SPT, CerS3, and CDase between groups AGED+RJ1 and AGED. These results indicate that dietary RJ1 improved epidermal hydration in middle-aged mice by increasing the generation of all Cer species with enhanced accumulation and hydrolysis of total GC, including all GC species, and that of all SM species to some extent (but not with alteration of de novo Cer synthesis and/or Cer degradation).
RJ consists of proteins, carbohydrates, lipids, mineral salts, and vitamins 34 together with significant amounts of bioactive substances. Among these substances, 10-HDA has been identified as a standardized major bioactive substance; it was present at a concentration of ∼2 mg/g (∼0.2%) in the RJ used in this study. 35,36 Royalisin 37 and apisin 38 have also been identified in RJ, but little is known about their beneficial effects on skin. The synthetic counterpart of 10-HDA has been reported to enhance epidermal hydration in UV-irradiated human subjects by restoring the skin barrier with increased protein expression of filaggrin, 9 degradation of which yields natural moisturizing factor, another constituent that maintains epidermal hydration. 39 On the other hand, ingested RJ alters serum levels of insulin growth factor-1, 40 which indirectly regulates the expression of antiaging- or longevity-related genes. 40,41 Major protein 1 of RJ has been found to enhance mRNA expression of tumor necrosis factor-alpha, the major cytokine involved in increasing aSMase activity, 42 in keratincoytes. 43 Together, these data suggest that synergistic interactions of RJ1 components with or independently from 10-HDA could indirectly promote systemic changes that enhance protein expression and/or activity of target enzymes in Cer and/or filaggrin metabolism. In this study, epidermal hydration in group AGED+RJ1 was far higher than in group C, which might be explained by alterations in filaggrin metabolism, but this remains to be elucidated in future studies.
RJ2, the RJ harvested from Pocheon, did not sufficiently improve epidermal hydration. In addition, epidermal levels of all Cer species, total GC (including most GC species), and GCase and aSMase proteins in group AGED+RJ2 were similar to those in group AGED, despite an increase in all SM species and SPT2 protein expression to levels similar to or higher than those in group C. Although de novo Cer synthesis and SM accumulation may be enhanced to a certain extent, dietary RJ2 did not enhance the accumulation of total GC (including most GC species), the major precursor lipid of Cer generation. 12,25 Moreover, dietary RJ2 did not increase the generation of Cer species from GC and SM, ultimately resulting in no apparent improvement in epidermal hydration in middle-aged mice. The less beneficial effect of dietary RJ2 in this study agrees with our earlier reports: namely that lower Cer and filaggrin levels are found in the epidermis of UV-irradiated 11 or ovariectomized animals fed RJ2 versus RJ1. 44 In our earlier study, 35,36 when 10-HDA was analyzed by high-performance liquid chromatography using a symmetry C18 column (250×4.6 mm i.d., S-4 μm, 80 Å; YMC Co., Ltd., Kyoto, Japan), which was run at a flow rate of 1.0 mL/min in a linear gradient of acetonitrile (A) and water (B) (0–20 min, 30–100% A; 20–25 min, 100% A) and detected at 210 nm, the 10-HDA content of RJ2 (2.01±0.008 μg/g, 0.202%) was less than that of RJ1 (2.114±0.0065 μg/g, 0.211%). These findings indicate compositional differences in RJ depending on the area of harvest. Although the content of other bioactive substances in RJ1 and RJ2 was not quantified, these data might suggest that the content of other bioactive substances in RJ2 are also likely to be less than that in RJ1. Synergistic interaction of RJ2 seems to cause fewer systemic changes in signaling responses, ultimately resulting in the less beneficial effect on epidermal hydration.
In conclusion, dietary RJ1 improved epidermal hydration by enhancing Cer metabolism with increased levels of all Cer, GC, and SM species, and of GCase and aSMase proteins. RJ1 might therefore be used as a dietary supplementation to improve epidermal hydration of intrinsically aged skin.
Footnotes
Acknowledgment
This work was supported by a grant from the Next-Generation BioGreen 21 Program (NO. PJ0090662013), Rural Development Administration, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.