
Abstract
Acetaminophen (APAP) overdose causes severe liver and kidney damage. APAP-induced liver injury (AILI) represents the most frequent cause of drug-induced liver failure. APAP is relatively insoluble and can only be taken orally; however, its prodrug, propacetamol, is water soluble and usually injected directly. In this study, we examined the time-dependent effects of AILI after propacetamol injection in mice. After analyses of alanine aminotransferase and aspartate aminotransferase activities and liver histopathology, we demonstrated that a novel AILI mouse model can be established by single propacetamol injection. Furthermore, we compared the protective and therapeutic effects of galangin with a known liver protective extract, silymarin, and the only clinical agent for treating APAP toxicity, N-acetylcysteine (NAC), at the same dose in the model mice. We observed that galangin and silymarin were more effective than NAC for protecting against AILI. However, only NAC greatly improved both the survival time and rate consequent to a lethal dose of propacetamol. To decipher the hepatic protective mechanism(s) of galangin, galangin pretreatment significantly decreased the hepatic oxidative stress, increased hepatic glutathione level, and decreased hepatic microsomal CYP2E1 levels induced by propacetamol injection. In addition, propacetamol injection also reproduced the probability of APAP-induced kidney injury (AIKI), appearing similar to a clinical APAP overdose. Only galangin pretreatment showed the protective effect of AIKI. Thus, we have established a novel mouse model for AILI and AIKI using a single propacetamol injection. We also demonstrated that galangin provides significant protection against AILI and AIKI in this mouse model.
Introduction
N-
Currently, N-acetylcysteine (NAC) is the only therapeutic agent meeting the clinical criteria for the treatment of APAP toxicity. 8 NAC detoxifies primarily by converting NAC into cysteine, then synthesizing GSH in liver cells to help the metabolism of NAPQI. NAC can also act as a sulfur provider, directly binding to APAP, forming nontoxic metabolites. Silymarin is a plant extract known to protect the liver. There is evidence supporting its antioxidant properties and abilities to inhibit lipid peroxidation and CYP activation; similarly, evidence suggests that it also has antifibrotic and immunomodulatory effects and can inhibit development of cell permeability and prevent mitochondrial damage. 9,10 Thus, silymarin protects from potential liver and kidney damages caused by APAP overdose. 11
Galangin is a natural flavonoid alcohol compound that can be extracted and isolated from galangal rhizome (Alpinia officinarum Hance), roots of Helichrysum aureonitens, and from propolis from various regions. It has been shown to exert significant antibacterial, 12,13 anticancer, 14 –16 antiobesity, 17 antioxidant, and anti-inflammatory 18 –20 effects. However, there is no report showing that galangin has any effect on acute liver and/or kidney injuries.
Establishments of APAP-induced liver injury (AILI) and kidney injury (AIKI) mouse models have many advantages, such as the ability to study APAP-mediated toxicological injury mechanisms, gene expressions affecting APAP toxicology, and drug screening for protection against APAP-induced toxic effects. 11,21,22 However, APAP is relatively insoluble in water and can only be taken orally in humans. It has slow analgesic onset and cannot be used immediately after surgery.
These disadvantages have been solved by using the U.S. FDA-approved APAP prodrug, propacetamol, in clinical practice. Propacetamol is water soluble and can be directly injected. After intravenous injection, propacetamol is completely hydrolyzed into APAP and diethylglycine by mammalian esterase within 7 min, 23 and 2 g of propacetamol is equivalent to 1 g of APAP. 24 However, there is currently no report for the use of propacetamol inducing AILI and AIKI in animals.
This study is the first, using injection of propacetamol, to establish AILI and AIKI model mice that mimic APAP overdose in patients. By comparing the effects of silymarin and NAC, we showed the protective effects of galangin in this mouse model.
Materials and Methods
Animals and reagents
Male 7-week-old BALB/c mice weighing 22–25 g were purchased from the National Laboratory Animal Center (Taipei, Taiwan). Mice were supplied with sterile water and rodent pellets ad libitum and maintained in a specific pathogen-free environment at the animal center at Chung Shan Medical University. All experimental procedures were conducted in accordance with the guidelines of the institutional animal committee (protocol number: 1239). Propacetamol, which is metabolized into acetaminophen and diethylglycine by a mammalian esterase (Fig. 1), was purchased from Standard Chem. & Pharm (Tainan, Taiwan); galangin (purity 99.5%) was purchased from Nanjing Zelang Medical Technology Co., Ltd., (Nanjing, China); and silymarin and NAC were purchased from Sigma-Aldrich (St. Louis, MO, USA).

The diagram shows the chemical metabolization of propacetamol into acetaminophen and diethylglycine by a mammalian esterase. The ratio of acetaminophen and diethylglycine formed is approximately 1:1.
Animal preparation and treatment
All animal experiments are summarized in Figure 2. Mice were randomly divided into different groups and fasted, but given tap water ad libitum for 12 h before an IP propacetamol injection. In time course experiments (n=5 for each group), blood was collected and mice were sacrificed for histopathological analysis at various time points after a single dose of 600 mg/kg propacetamol (equal to 300 mg/kg APAP) injection. In protection experiments (n=5 for each group), mice in the hepatotoxicity group were orally fed with the same dose (125 mg/kg) of galangin, silymarin, or NAC 1 h before a single dose of 600 mg/kg propacetamol injection, respectively. Mice were sacrificed 18 h after injection, and blood was collected just before sacrifice. Livers were isolated for analyses of GSH, TBARS, nitrotyrosine, and CYP2E1 levels.
Experimental design and time chart for the three animal experiments, including time course, protection, and therapy. Color images available online at
For therapy experiments (n=10 for each group), the same dose of galangin, silymarin, or NAC was orally administered 30 min after a single dose of 1200 mg/kg propacetamol (equivalent to 600 mg/kg APAP) injection. After injection, survival of mice was monitored every hour for 36 h. Galangin, silymarin, and NAC were suspended or dissolved in ultrapure water, and then administered a body weight-adjusted dose through oral gavage.
Measurement of serum ALT and AST activities and levels of BUN, CREA, and UA
Blood samples were collected from mouse tails, and then sera were collected after centrifugation. Sera were stored at −80°C for later use. Serum ALT and AST activities were determined using commercial assay kits (Randox, Inc., County Antrim, United Kingdom). Serum BUN, CREA, and UA levels were determined by the National Laboratory Animal Center.
Measurement of hepatic GSH and TBARS levels
The liver GSH level was determined using a glutathione assay kit (Cayman Chemical, Ann Arbor, MI, USA) according to the manufacturer's protocol. Lipid peroxidation in the liver was determined by measuring TBARS formation using the QuantiChrom™ TBARS Assay Kit (BioAssay Systems, Hayward, CA, USA) according to the manufacturer's protocol.
Microsome isolation
Mouse liver microsomes were prepared by homogenizing liver samples according to a previous report. 25
Western blotting
Protein concentration was determined using the Lowry method. 26 Forty micrograms of liver protein or ten micrograms of microsomal protein from each sample was analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and separated proteins were then transferred onto a polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA) for western blotting. Primary antibodies against nitrotyrosine (Abcam, Cambridge, MA, USA), CYP2E1 (Abcam), or β-actin (Cell Signaling Technology, Beverly, MA, USA) were used. The procedure of western blotting was followed according to a previous report. 27
Histopathology
Samples of the right lobe of the liver and the right kidney were fixed in 10% neutral buffered formalin and then embedded in paraffin. Five-micrometer-thick sections were prepared and stained with hematoxylin and eosin. Ten random fields of each section were analyzed by light microscopy. Liver histological scoring was performed by a pathologist according to a previous report. 28 In brief, liver injury was scored as follows: 0, no damage; 1, only a few fields affected; 2, zonal necrosis in most fields; 3, at least 3 foci of zonal necrosis in all fields; 4, centrilobular necrosis in most lobes in all fields; 5, panlobular confluent necrosis and hemorrhaging in all fields. Histopathological changes in kidneys were scored on a 5-point scale: 0=no damage, 1=0–25%, 2=25–50%, 3=50–75%, and 4=more than 75% of indicating degree of injury to the corticomedullary junctions as assessed by a pathologist according to a previously reported method. 29
Statistical analyses
The data are expressed as the mean±standard deviation (SD). The mean and SD of targeted analysis levels were calculated for clinical chemistry and western blot, and metabolite data were assessed using Microsoft Office Excel 2010 software. The statistical differences between groups were analyzed by t-test and considered to be significant at P<.05. A Kaplan–Meier analysis was used for estimation of survival curves, and a log-rank test was used to determine differences between two survival curves. Liver histopathological examination data were analyzed by the Kruskal–Wallis nonparametric test. Two-tailed P-values of less than .05 were considered statistically significant. SPSS software (version 10.1, SPSS, Inc., Chicago, IL, USA) was used for these statistical analyses.
Results
IP injection of propacetamol to induce AILI in mice
AILI was induced by injecting the mice with propacetamol for studying liver injury in a time course experiment (Fig. 2, Time Course). Mice were fasted for 12 h before an IP injection of 600 mg/kg propacetamol. After injection, peripheral blood was collected at 6, 12, 18, and 24 h for analyses of ALT and AST activities and their livers were isolated for histopathological analysis. ALT activities within mouse sera at 6, 12, 18, and 24 h after injection were significantly higher (100, 240, 370, and 320 times) than at 0 h (Fig. 3A). AST activities at 6, 12, 18, and 24 h after injection were also significantly increased (40, 90, 105, and 110 times) when compared with at 0 h postinjection (Fig. 3B). Furthermore, all liver sections showed significantly more cell necrosis (Fig. 3D–G).
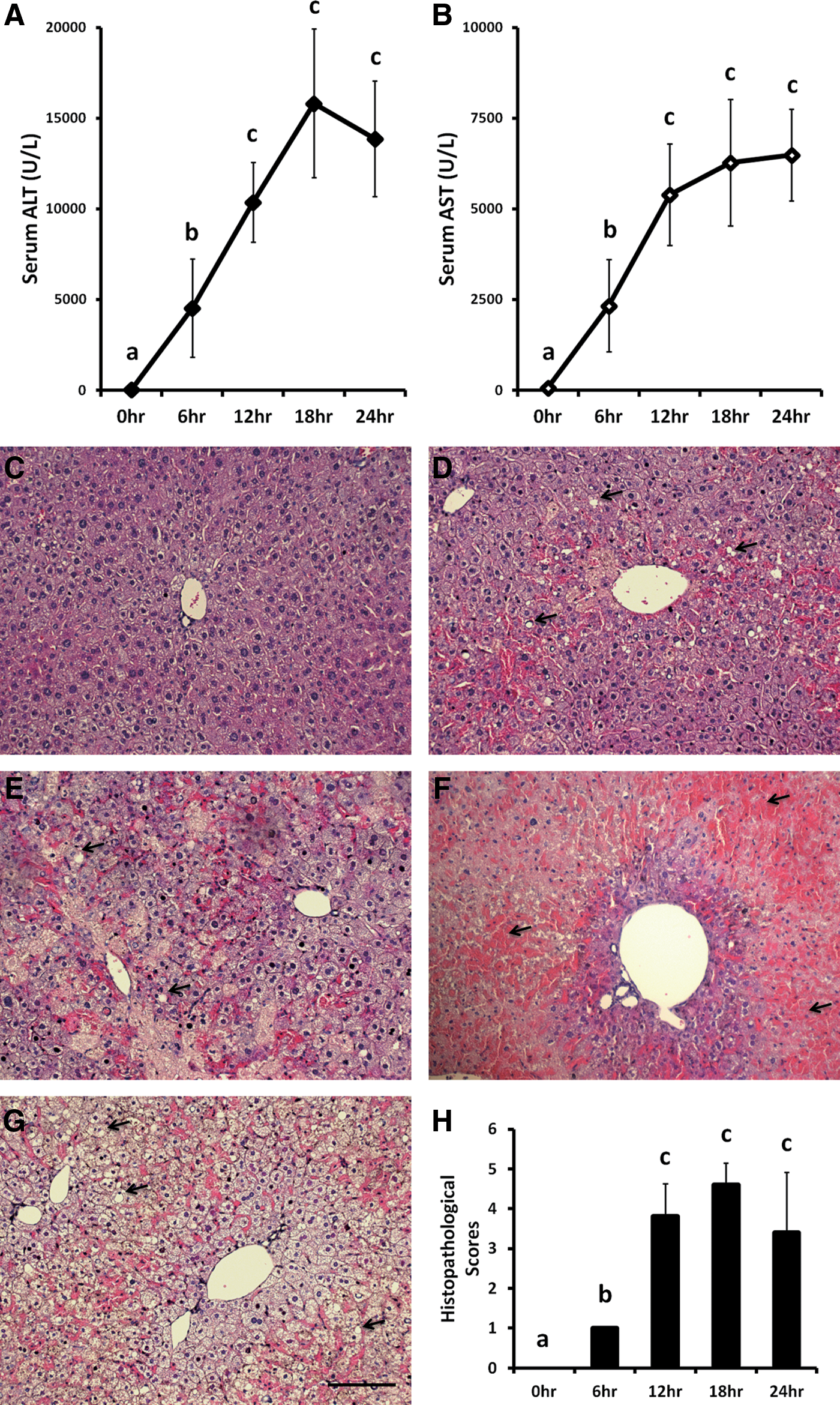
The ALT and AST levels in peripheral blood and liver biopsy pathology after injection of propacetamol in mice. ALT
Liver sections at 6 h postinjection showed some centrilobular necrosis, but at 12 h postinjection showed centrilobular necrosis in most lobes in all fields. Confluent panlobular necrosis and hemorrhaging were observed in all fields in liver sections at both 18 and 24 h postinjection. Results of histopathological scores show significant liver damages after 6 h postinjection (Fig. 3H).
From results in Figure 3, we concluded that IP injection of 600 mg/kg propacetamol indeed induced AILI in mice. Because both ALT and AST activities (Fig. 3A, B) and results of histopathological scores (Fig. 3H) showed no significant differences between mice at 18 and 24 h postinjection and mice were nearly dead and blood was difficult to collect through the tail vein in 24 h postinjection, we used the time point of 18 h postinjection for further experiments.
Galangin pretreatment reduces the severities of AILI induced by propacetamol injection
In protection experiments, twofold elevated doses of galangin were administered to mice 1 h before the induction of AILI by propacetamol injection. Peripheral blood was collected for analyses of ALT and AST activities, and livers were isolated for histopathological analysis. In comparison with mice injected with propacetamol only, mice fed 62.5 mg/kg of galangin showed approximately 0.75 times the ALT activity (11,825±2497) and 0.9 times AST activities (5589±776), but these differences were not significant (Fig. 4A, B). Mice fed 125 mg/kg of galangin showed the most significantly lower activities of both ALT and AST at 0.15 times (2404±2165) and 0.13 times (776±868), respectively. Mice fed 250 mg/kg of galangin also exhibited significantly lower ALT and AST activities at 0.46 times (7299±4133) and 0.37 times (2332±1527), respectively. Furthermore, the liver biopsy results (Fig. 4C–G) were consistent with ALT and AST activities. Statistical analyses of histopathological scores showed significant decreases in both 125 and 250 mg/kg galangin pretreatments when compared with the propacetamol only (Fig. 4H). Pretreatment with 125 mg/kg galangin also provided the greatest protection against liver damage with a mean histopathological score of approximately 1/3 of the propacetamol only. Results of protection experiments demonstrated that prefeeding galangin decreased AILI induced by propacetamol injection, and the dose of 125 mg/kg showed the most protection.
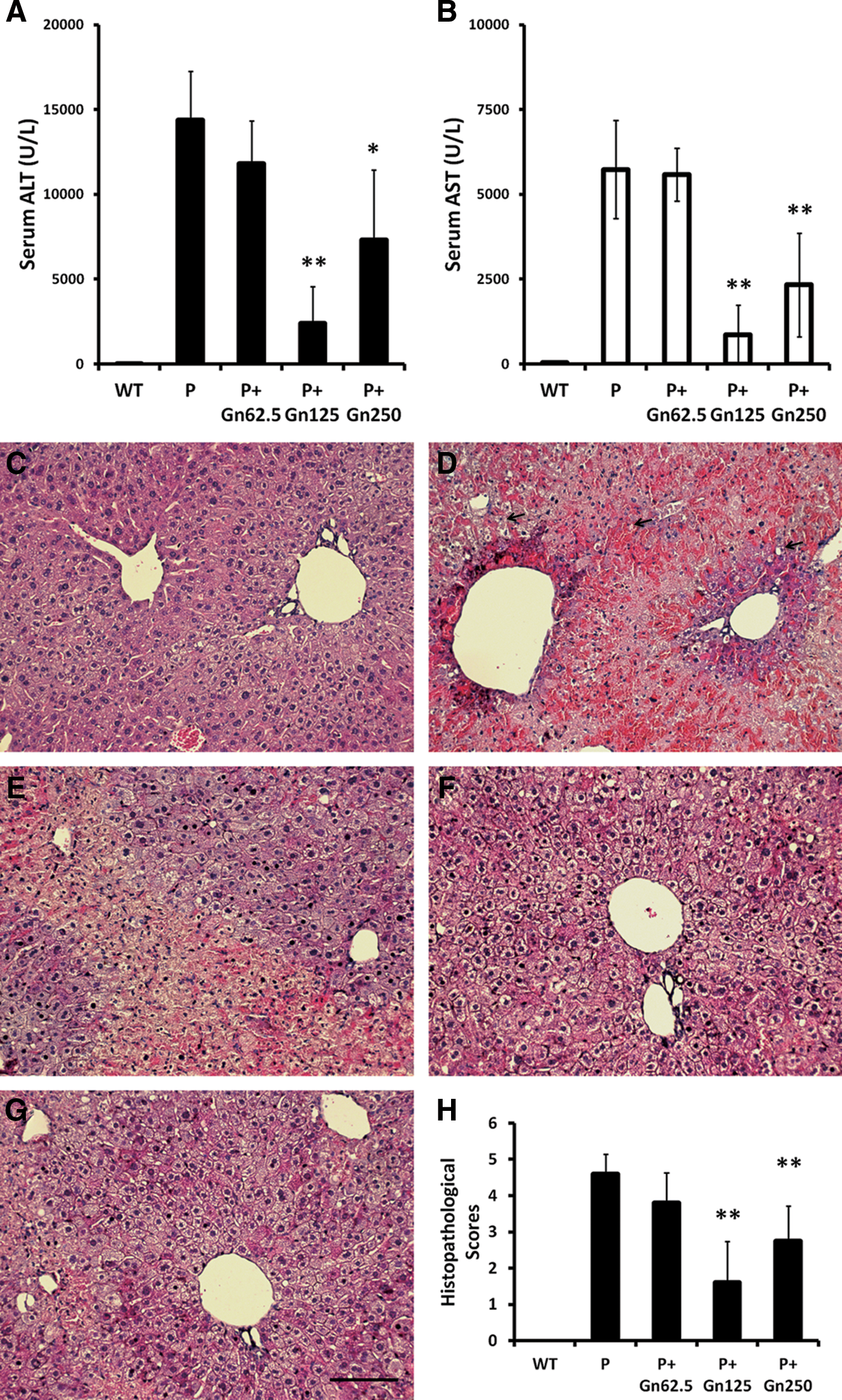
Galangin pretreatments showed significant liver protective effects in AILI mice induced by propacetamol injection. The mice were pretreated with different doses of galangin. ALT
Comparisons of protective effects with the same dose of NAC, silymarin, and galangin in AILI mice
The same dose of NAC, silymarin, or galangin was fed to mice before AILI was induced by propacetamol injection, and their liver protective effects were compared. Results showed that all pretreatments with NAC, silymarin, and galangin significantly lowered blood ALT and AST activities when compared with mice without pretreatment (Fig. 5A, B). ALT activities were decreased to approximately 0.5, 0.14, and 0.15 times, while AST activities were decreased to approximately 0.6, 0.13, and 0.13 times for NAC, silymarin, and galangin pretreatments, respectively. Silymarin and galangin showed greater protection than NAC, and there was no significant difference between them. Furthermore, both liver section staining (Fig. 5C–G) and histopathological scores (Fig. 5H) also showed similar trends. Areas of liver injury were decreased for NAC (0.7 times), silymarin (0.5 times), and galangin (0.4 times) pretreatments compared with no pretreatment. In brief, results of Figure 5 indicated that all pretreatments with NAC, silymarin, and galangin significantly protected AILI induced by propacetamol injection, and both galangin and silymarin were more protective than NAC.
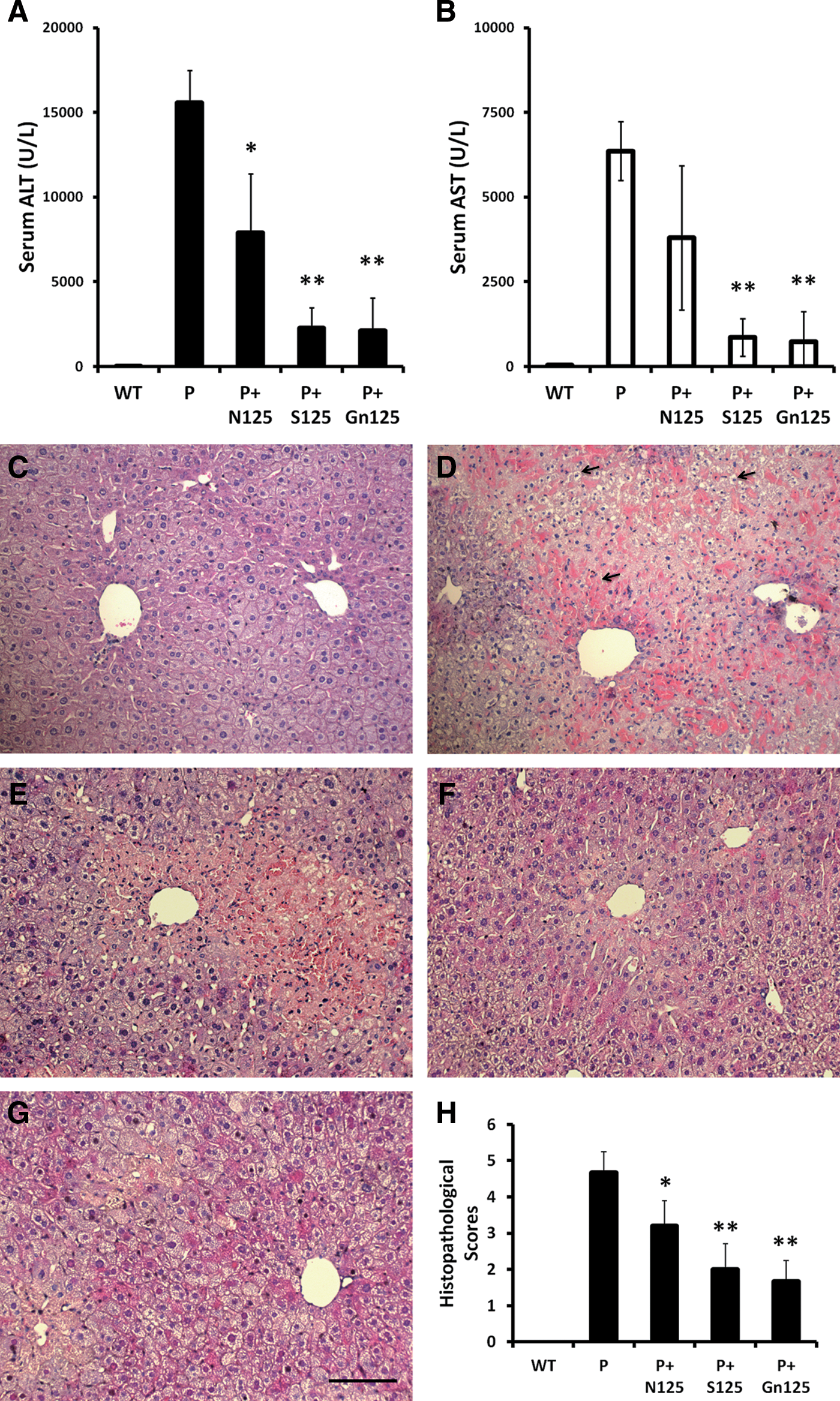
ALT and AST values in peripheral blood and changes in liver biopsies of propacetamol-induced AILI mice after treatment with the same doses of NAC, silymarin, and galangin. ALT
Double doses of propacetamol induce lethal AILI; comparisons of rescue effects of NAC, silymarin, and galangin
To decipher whether a higher dose of propacetamol could be lethal in mice as observed in clinical APAP overdose, mice were injected with a double dose of propacetamol to induce fatal AILI. Half an hour after injection, mice were fed the same dose of NAC, silymarin, or galangin as a therapy for fatal AILI. Mice without therapy began to die at 6 h postinjection and all had died within 13 h postinjection (Fig. 6). Therefore, injection of a higher dose of propacetamol indeed induces fatal AILI in mice.
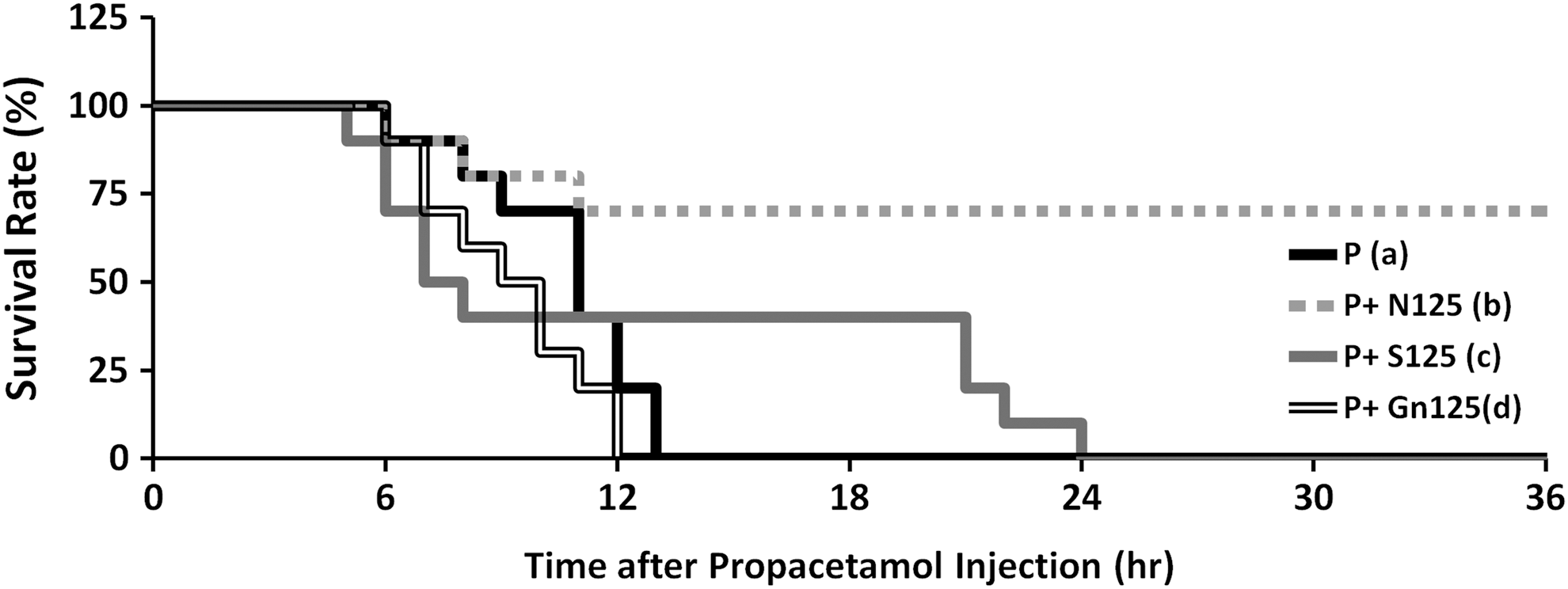
Survival of mice treated with NAC, silymarin, or galangin used as an emergency treatment after the injection of a lethal dose of propacetamol. Survival of mice treated with a 125 mg/kg dose of NAC (P+N125, b), silymarin (P+S125, c), or galangin (P+Gn125, d) as an emergency treatment half an hour after the injection of propacetamol. Survival of mice injected with only propacetamol (P, a). There is a significant difference only between (a) and (b) groups with P<.01.
Results of therapy experiments showed that only NAC was significantly effective for rescuing fatal AILI mice (Fig. 6). Although the first mouse in the NAC group died at 6 h postinjection, no more mice died until after 11 h postinjection and the survival rate remained at 70% until the end of the experiment. Although galangin was very protective against liver injury in AILI mice (Figs. 4 and 5), there was no significant therapeutic effect of galangin in fatal AILI mice (Fig. 6). Mice began to die at 6 h postinjection and all had died within 12 h postinjection. For therapy with silymarin, although the first mouse died at 5 h postinjection and initially showed the most rapid death rate, no more mice died from 8 to 21 h postinjection and 40% of mice survived for 21 h postinjection; however, all mice had died within 24 h postinjection. Therefore, the therapeutic effectiveness of silymarin was not as effective as the same dose of NAC.
We found that both silymarin and galangin exerted more protective effects in AILI mice, but were less effective for preventing fatality in high-dose AILI mice. However, NAC, which showed a less obvious protective effect in AILI mice, was the most therapeutic for fatal AILI.
Propacetamol injection induces kidney damages; comparisons of protective effects of NAC, silymarin, and galangin
Peripheral blood and kidneys were collected at 0, 6, 12, 18, and 24 h after propacetamol injection; BUN, CREA, and UA were measured in the sera (Fig. 7A–C). BUN values began to rise at 6 h postinjection and were significantly increased at 18 and 24 h to approximately 2.5 and 4.8 times of those observed at 0 h, respectively. CREA values increased slowly and began to rise prominently at 24 h postinjection; however, there were no significance differences because of large variations between mice at each time point. UA values were significantly decreased to approximately 0.22 times at 6 h postinjection and continued to the last time point.
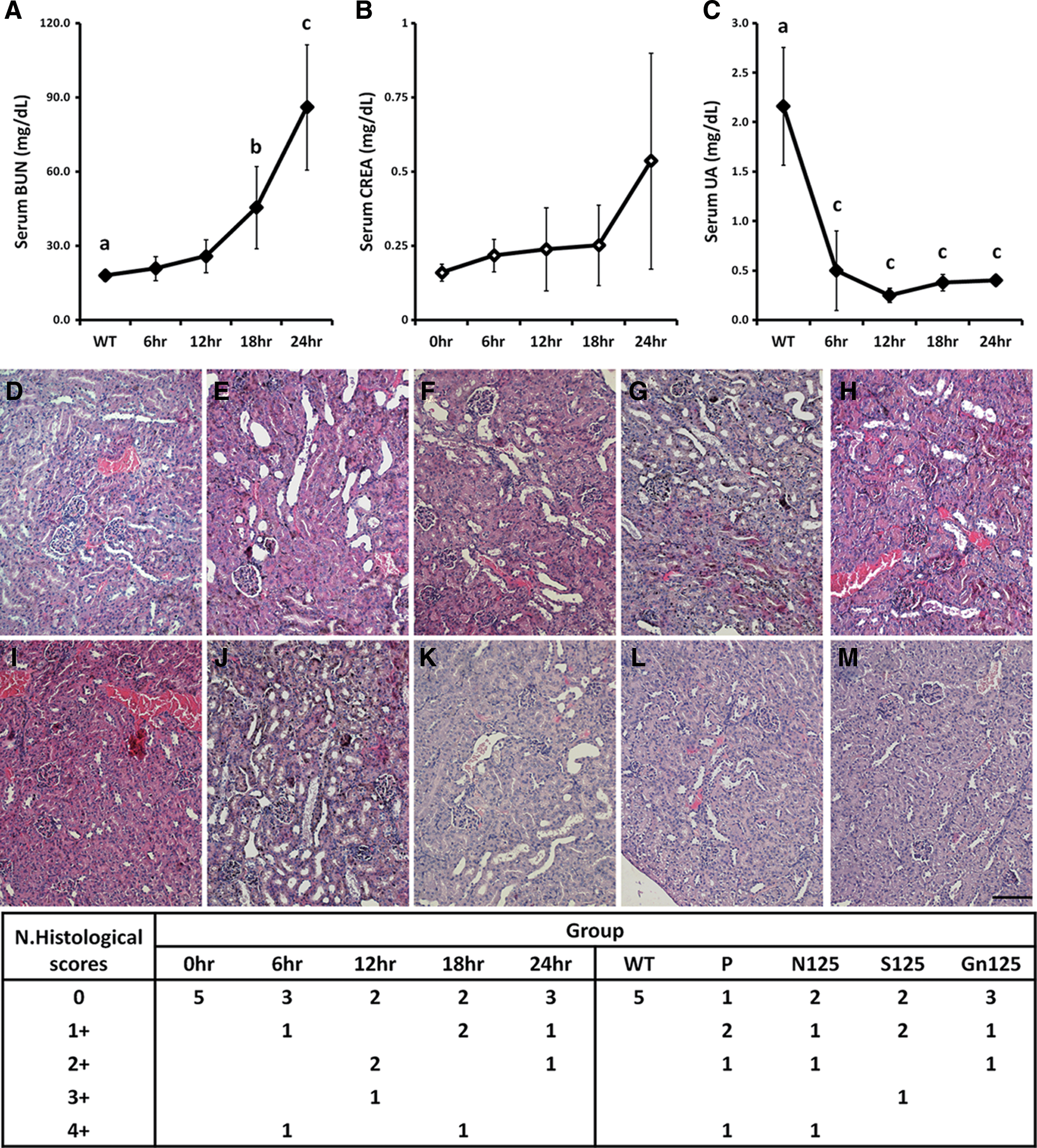
Biochemical indices of renal injury in the peripheral blood and renal biopsy changes following injection of propacetamol. BUN
Results of the kidney section staining indicated that tubular epithelial damage, tubular dilatation, and intratubular cast formation had developed at 6 h postinjection, and these lesions also appeared at 12, 18, and 24 h postinjection (Fig. 7D–H). However, results of histopathological scores did not show more severe kidney damage developing over time (Fig. 7N, left). These renal pathological analyses showed that propacetamol injection also induced AIKI in a subset of mice similar to those observed in APAP overdose patients.
To test and compare protective effects of NAC, silymarin, and galangin, these compounds were fed 1 h before propacetamol injection and their protective effects on kidneys were evaluated. Histopathological results showed that galangin and silymarin, but not NAC, reduced AIKI caused by propacetamol (Fig. 7I–M). Furthermore, results of histopathological scores showed that galangin had the greatest protective effect (Fig. 7N, right).
Galangin pretreatment decreases propacetamol-induced oxidative stress and levels of CYP2E1 in mouse livers; comparisons of protective effects of NAC and galangin
To prove that IP injection of propacetamol increases oxidative stress and drug metabolism in mouse liver as IP injection of APAP, we first assayed the levels of lipid peroxidation (TBARS) and GSH in mouse livers. Lipid peroxidation was significantly increased (7.3 times), but the GSH level was significantly decreased (0.43 times) in mouse liver by propacetamol injection when compared with normal mice (Table 1). Furthermore, both of the nitrotyrosine adducts (1.90 times) and microsomal CYP2E1 levels (3.00 times) were significantly increased in mouse liver by propacetamol injection when compared with normal mice (Fig. 8). These results indicated that injection of propacetamol increased oxidative stress and drug metabolism in mouse liver as APAP injection.

Hepatic expression levels of nitrotyrosine adducts and microsomal CYP2E1 in mice of different experimental groups. Normal mice (WT), mice injected with propacetamol (P), and mice pretreated with 125 mg/kg of galangin (P+G) or NAC (P+N) 1 h before a propacetamol injection that induces AILI 18 h later. Expression levels of nitrotyrosine adducts and microsomal CYP2E1 in mouse livers were analyzed by western blotting
Values are average±SD (n=3). Hepatic TBARS and GSH levels of normal mice (WT), mice injected with propacetamol (P), and mice pretreated with 125 mg/kg of galangin (P+Gn125) or NAC (P+N125) 1 h before propacetamol injection. Different superscript letters indicate statistically significant differences between groups and are indicated with * (P<.05) and ** (P<.01). For TBARS values, the P group showed a significant difference with the WT group (P<.01); the P+Gn125 group showed a significant difference with the P group (P<.01), but no difference with the WT group; and the P+NAC125 group showed significant differences with all of WT, P, and P+Gn125 groups (all P<.01). For GSH values, the P group showed a significant difference with the WT group (P<.05); the P+Gn125 group showed a significant difference with the P group (P<.01), but no difference with the WT group; and the P+NAC125 group showed significant differences with all of WT, P, and P+Gn125 groups (all P<.01).
TBARS, thiobarbituric acid-reactive substances; GSH, glutathione; NAC, N-acetylcysteine; AILI, APAP-induced liver injury.
To decipher the liver protective mechanism(s) of galangin and compare the protective effects of galangin and NAC, we pretreated the mice with the same dose of galangin or NAC 1 h before propacetamol injection. Pretreatments with the same dose of both galangin and NAC resulted in significantly lower hepatic levels of lipid peroxidation, approximately 0.13 and 0.27 times, and significantly higher hepatic levels of GSH to approximately 4.14 and 7.00 times (Table 1). Furthermore, galangin pretreatment caused significantly lower hepatic expression levels of nitrotyrosine adducts (0.17 times) and microsomal CYP2E1 (0.20 times) induced by propacetamol injection, whereas NAC pretreatment resulted only in significantly lower hepatic nitrotyrosine levels (0.36 times), but not significantly lower microsomal CYP2E1 levels (0.44 times) (Fig. 8). These results indicated that both galangin and NAC pretreatments significantly decreased the hepatic oxidative stress (both TBARS and nitrotyrosine) induced by propacetamol injection and increased the major reducing component (GSH). Only galangin pretreatment significantly decreased hepatic microsomal CYP2E1 levels induced by propacetamol injection. In brief, galangin is more effective for decreasing hepatic oxidative stress caused by propacetamol injection, while NAC is more effective for increasing the hepatic GSH level.
Discussion
The two major findings of this study are 1. establishing novel AILI and AIKI model mice by propacetamol injection and 2. identifying the hepatorenal protective effect and possible protective mechanism of galangin.
APAP overdose in rodents is one of the most common and convenient models used to identify compounds with hepatoprotective effects. 30,31 The current experimental method to induce AILI in animals was to dissolve the relatively insoluble APAP in water or PBS by heating especially in high concentration, and then an adequate cooling is needed before IP injection into animals. Heating the APAP solution for too long might cause APAP degradation, and cooling the APAP solution to room temperature might precipitate APAP. Thus, APAP dissolved in warm PBS or water (usually higher than 50°C) was usually directly injected into animals. 32 The manipulation is not only difficult for researchers but also causes pain to animals. A transient moderate hepatotoxicity may be obtained. 25 For researchers without experience, more laboratory animals may be needed for inducing AILI by using APAP.
However, the APAP prodrug, propacetamol, is very water soluble and already used in clinics. In this study, mice injected with propacetamol showed classic AILI and AIKI as APAP injection. These AILI mice induced by propacetamol injection showed significant increases in peripheral ALT and AST activities and liver injury (Fig. 3) and significant increases in hepatic oxidative stress (Table 1 and Fig. 8B) and drug metabolism (Fig. 8D). Furthermore, these AILI mice can be protected by pretreatment with a known liver protective extract, silymarin, 11 and lethal AILI mice can be rescued by the only clinical agent for treating APAP toxicity, NAC. 8
Furthermore, excessive use of APAP causes acute liver injury and probably causes development of acute and chronic kidney disease. 7 As shown in Figure 7, there were significant differences of UA values in peripheral sera starting at 6 h postinjection, while BUN values began to show significant differences at 18 h. Kidney tubular injury became evident histologically at 6 h postinjection (Fig. 7E) with a likelihood of 40% to 60%; however, the likelihood of kidney damage did not increase with time (Fig. 7N, left). Thus, propacetamol injection also reproduced the probability of AIKI appearing similar to the clinical APAP overdose.
Galangin, a member of the flavonol class of flavonoids, is the active component of the rhizome of Alpinia galangal. Galangin has antioxidative and free radical scavenging activity. It has been found to inhibit inducible nitric oxide synthase and cyclooxygenase-2 transcriptions, which may contribute to an anti-inflammatory response. 18 It also prevents oxidative damage and downregulates the inflammatory pathway in the liver of fructose-fed rats. 19 In this study, pretreatment with galangin significantly reduced AILI induced by propacetamol injection (Fig. 4) by increasing GSH levels, decreasing lipid peroxidation (TBARS) (Table 1) and protein oxidation (nitrotyrosine), and decreasing one of the major enzymes for drug metabolism (CYP2E1) (Fig. 8). The mechanism of galangin pretreatment against AILI is majorly through reducing hepatic oxidative stress caused by propacetamol injection.
The influences of galangin pretreatment were more effective than NAC pretreatment, except for GSH levels, so galangin showed greater protection than NAC in this AILI mouse model (Fig. 5). Furthermore, only galangin significantly decreased the probability of AIKI induced by propacetamol injection (Fig. 7N, right). However, galangin showed no therapeutic effect against fatality in AILI mice (Fig. 6). Since APAP toxicity is related to GSH depletion, galangin treatment could not elevate hepatic GSH levels compared with NAC (Table 1) and may account for lack of rescue by galangin.
For using propacetamol injection to induce AILI model mice, some points need to be addressed. First, after dissolving the propacetamol powder with PBS, the propacetamol solution can be warmed to 37°C and should be immediately injected. Second, propacetamol has been clinically used by IV injection. For replacing the current method of using IP injection of APAP, IP injection of propacetamol was used in this study. The results showed similar hepatic toxicology of IP injection of propacetamol as IP injection of APAP does. Thus, the injection of propacetamol can be IP or IV in mice. Finally, AILI induced by IP injection of APAP is usually analyzed at 12 h postinjection. We suggest analyzing AILI induced by IP injection of propacetamol at 18 h postinjection because propacetamol should be completely hydrolyzed into APAP and diethylglycine by mammalian esterase. It takes time to cause APAP overdose.
Because of the convenient usage of propacetamol, we look forward to replacing the current method of APAP injection with propacetamol injection to induce AILI in laboratory animals for screening hepatoprotective drugs or components in the future.
Footnotes
Acknowledgments
This study was financially supported by grant aid from the National Science Council of Taiwan (NSC99-2314-B-212-001-MY3, NSC100-2320-B-040-012). Upright fluorescence microscopy was performed in the Instrument Center of Chung Shan Medical University, which is supported by the National Science Council, the Ministry of Education, and Chung Shan Medical University.
Author Disclosure Statement
No competing financial interests exist.