
Abstract
Angiotensin II (Ang II) is a very important cardiovascular disease inducer and may cause cardiac pathological hypertrophy and remodeling. We evaluated a Chinese traditional medicine, alpinate oxyphyllae fructus (AOF), for therapeutic efficacy for treating Ang II-induced cardiac hypertrophy. AOF has been used to treat patients with various symptoms accompanying hypertension and cerebrovascular disorders in Korea. We investigated its protective effect against Ang II-induced cytoskeletal change and hypertrophy in H9c2 cells. The results showed that treating cells with Ang II resulted in pathological hypertrophy, such as increased expression of transcription factors NFAT-3/p-NFAT-3, hypertrophic response genes (atrial natriuretic peptide [ANP] and b-type natriuretic peptide [BNP]), and Gαq down-stream effectors (PLCβ3 and calcineurin). Pretreatment with AOF (60–100 μg/mL) led to significantly reduced hypertrophy. We also found that AOF pretreatment significantly suppressed the cardiac remodeling proteins, metalloproteinase (MMP9 and MMP2), and tissue plasminogen activator (tPA), induced by Ang II challenge. In conclusion, we provide evidence that AOF protects against Ang II-induced pathological hypertrophy by specifically inhibiting the insulin-like growth factor (IGF) II/IIR-related signaling pathway in H9c2 cells. AOF might be a candidate for cardiac hypertrophy and ventricular remodeling prevention in chronic cardiovascular diseases.
Introduction
A
Angiotensin II (Ang II), an octapeptide hormone, is an important player in the regulation of cardiovascular homeostasis. However, it is also considered to cause pathological cardiovascular tissue remodeling, such as atherosclerosis, hypertension, left ventricular hypertrophy (LVH), and heart failure. 4 –11
Most of the known physiological effects of Ang II in the cardiovascular system occur predominantly through the Ang II type-1 receptor (AT1R). 4,10,12 –14 In the stressed heart, Ang II and mechanical stress mediate AT1R activation to induce cardiac hypertrophy and stimulate intracellular effector systems. 15 AT2R has recently also been considered to be involved in some important Ang II actions in the heart. 16,17 Overexpression of angiotensin receptors has been demonstrated to increase in cardiac hypertrophy. 18,19 Although the functions of these two Ang II receptors are ambiguous, the harm of Ang II to cardiomyocytes is certain. 6,20,21
Previous studies indicate that insulin-like growth factor II (IGFII) contributes to cellular hypertrophy, remodeling, and apoptosis. 22 –25 Our recent studies found that IGFIIR gene expression is also up regulated in both pathological cardiac hypertrophy and with Ang II treatment. Ang II-induced apoptosis was found to be induced by extracellular signal-regulated kinase (ERK), c-Jun NH2-terminal kinase (JNK), and calcineurin pathways. 25,26 The evidence demonstrated that Ang II-induced IGFIIR gene expression occurred through histone acetylation. 26,27
It is known that Ang II binds to AT1-R to induce prohypertrophic effects and stimulate diverse intracellular signaling through Gaq, activating its downstream effector calcineurin, which induces immediate-early genes c-jun, jun B, Egr-1, c-fos, and c-myc. The protein products of fetal type genes such as the atrial natriuretic peptide (ANP), b-type natriuretic peptide (BNP), and skeletal α-actin will then be overexpressed, 11,28 –30 ultimately causing heart failure.
Alpinate oxyphyllae fructus (AOF; Alpinia oxyphylla MIQ) is an important traditional Chinese medicinal herb. According to the Chinese Pharmacopoeia, AOF has been widely used for dealing with diarrhea, polyuria, ulceration, dementia, antitumor, and gastralgia. 31 Several experiments indicated that the extract from AOF kernels possesses significant neuroprotective activity. 31 –35 It has been reported that aqueous or ethanolic extracts of AOF possess cardioprotective effectiveness. 36,37 AOF has been used for the treatment of cerebrovascular disorders and various symptoms accompanying hypertension in Korea. 33 Our previous reports show that AOF extracts effectively inhibit Ang II-induced cardiac apoptosis after calcineurin in H9c2 cardiomyoblast cells. 37 This study was specially designed to investigate the effectiveness of AOF for repressing these upstream IGF2R signaling factors.
Materials and Methods
AOF extraction
AOF (Alpinia oxyphylla MIQ) was obtained from Shin-Long Pharmaceutical Company (Taichung, Taiwan), 150 g was extracted with 0.6 L of boiling distilled water for 2 h and filtered by gravity-flow procedure at reduced pressure for convenience. This extract was spray dried at 17–20 mL/min and atomized through a 1-mm nozzle using compressed air. The product yield percentage was 7.2%.
Cell culture
H9c2 cardiomyoblasts were obtained from the American Type Culture Collection (Manassas, VA). H9c2 cells were cultured in Dulbecco's modified Eagle's medium in 100-mm culture dishes and supplemented with 10% fetal bovine serum, 100 U/mL penicillin, 2 mM glutamine, 100 μg/mL streptomycin, and 1 mM HEPS buffer. The cultures were incubated at 37°C in humidified air with 5% CO2. The medium was replaced every other day. Before treatment with indicated agents, the cells were incubated in serum-free essential medium for 4 h.
Cell viability assay
Cell viability was determined using the MTT assay. H9c2 cells were cultured in 12-well culture plates at a density of 2 × 104 cells per well and incubated with different concentrations of AOF (0, 20, 40, 60, 80, and 100 μg/mL) for 24 h at 37°C, 5% CO2. The culture medium was subsequently replaced with 500 μL of MTT (0.5 mg/mL) solution. After culture at 37°C in darkness for 4 h, the MTT solution was carefully removed and 200 μL of dimethyl sulfoxide was added to solubilize the formazan product. The plate was measured using an automated microplate reader at 570 nm. The experiment was repeated three times.
Actin-immunofluorescence (rhodamine phalloidin)
H9c2 cells were fixed with 4% paraformaldehyde at room temperature (RT) for 30 min. Cells were washed five times with ice-cold phosphate-buffered saline (PBS) and permeabilized with 0.5% Triton X-100 for 10 min at 4°C. Nonspecific fixed cell binding was blocked with PBS containing 1% bovine serum albumin (BSA) at RT for 20 min. The cells were then stained with rhodamine phalloidin (1:100 dilution) for 1 h to observe F-actin. Cells were examined and photographed using a Zeiss Axioskop (Carl Zeiss Microscopy GmbH, Heidelberg, Germany) and fluorescence microscope. The cell sizes were magnified 400 times and analyzed using digital image analysis, using Zeiss Axio Vision software (Carl Zeiss Microscopy GmbH). The cell surface area was determined after culturing in six-well plates (5 × 104 cells/well) and imaging by fluorescence microscopy. The surface area was quantified by imaging to the complete boundary of individual cells in various conditions using Zeiss Axio Vision software (Carl Zeiss Microscopy GmbH). Thirty cells were counted in each condition and regarded as an independent experiment. Three independent experiments were performed in each condition.
Nucleus observation using DAPI staining
Cells grown in six-well plates were divided into several groups for various treatments. After treatment, the cells were fixed with 4% paraformaldehyde solution for 30 min at RT. After rinsing with PBS, the washed cells were then stained with 1 μg/mL 4′-6-diamindine-2-phenylindole dihydrochloride (DAPI; Roche Molecular Biochemicals, Basel, Switzerland) for 30 min in the dark. Stained cell images were recorded with a fluorescent microscope.
Immunofluorescence
Cells grown in six-well plates and subjected to various treatments were subsequently washed with ice-cooled PBS and fixed with 4% paraformaldehyde in PBS for 30 min. Cells were rinsed with PBS and permeabilized with 0.5% Triton X-100 at 4°C for 10 min and blocked with 1% BSA dissolved in PBS at RT for 20 min. After removing the blocking solution, the section was incubated with primary NFAT-3 antibodies overnight at 4°C. After washing, cells were incubated with antigoat bright green-fluorescent Alexa Fluor® 488 dye-conjugated antibodies (Molecular Probes, Inc., Eugene, OR) at 37°C for 1 h. Cells were then stained with bright green-fluorescent Alexa Fluor 488 dye-conjugated antibodies (Molecular Probes, Inc.). The fluorescence was observed using a fluorescent microscope coupled with an image analysis system.
Immunoblotting
Cultured myocardiac cells were lysed with lysis buffer (50 mM Tris, pH 7.5, 1.0 mM EDTA, pH 7.5, 1 mM BME, 0.5 M NaCl, 10% glycerol, 1% IGEPAL-630, and proteinase inhibitor cocktail tablets [Roche Molecular Biochemicals]) for 30 min on ice and centrifuged at 12,000 g for 30 min at 4°C to isolate total proteins. The supernatant was then collected for Western blot analysis. The cell lysate protein concentrations were determined using the Bradford method. Samples containing equal proteins (35 μg) were separated in 12% gradient SDS-PAGE and transferred to polyvinylidene fluoride (PVDF) membrane (Millipore, Belford, MA). Nonspecific protein binding was blocked with blocking buffer (5% nonfat dry milk, 20 mM Tris-HCl, pH 7.6, 150 mM NaCl, and 0.1% Tween 20) for at least 1 h at RT. Membranes were incubated with specific antibodies on an orbital shaker at 4°C overnight and incubated with horseradish peroxidase-linked secondary antibodies (antirabbit, antimouse, or antigoat IgG) at RT for 1 h.
Statistical analysis
Quantitative data are shown as the mean ± SEM, corresponding to three or more independent experiments. Statistical comparisons were made using the Student's t-test. Statistical significance was defined at the P < .05, .001, or .0001 levels.
Results
AOF effects on cell viability
We evaluated the proliferative effect of AOF on H9c2 cells. During these experiments we first observed the effect of various AOF concentrations (0, 20, 40, 60, 80, and 100 μg/mL) on cell viability for 24 or 48 h. We found that cellular viability was significantly elevated at the 60, 80, and 100 μg/mL concentrations for 24 h (Fig. 1).
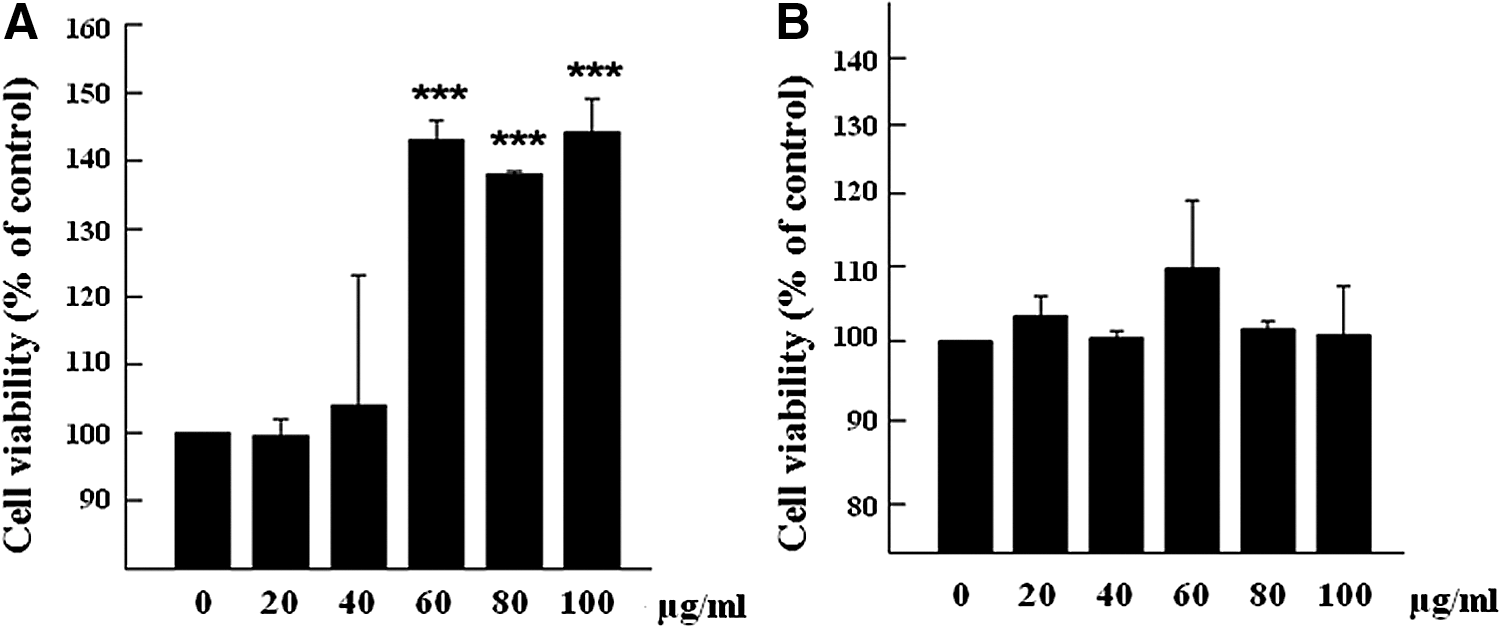
AOF effect on H9c2 cell viability. H9c2 cells were treated with 0–200 μg/mL AOF for
To clarify whether AOF would prevent Ang II-induced cytoskeletal changes and hypertrophy in H9c2 cells, cell size was measured using actin-staining (red image), and DAPI was used to stain cell nuclei (blue spots) (Fig. 2A). We observed the increase in cell size induced by Ang II was significantly decreased with AOF pretreatment at 60–100 μg/mL concentrations (Fig. 2B). Moreover, the increased levels of hypertrophic protein ANP and BNP from Ang II were decreased by AOF treatments (Fig. 2C).

AOF effect on Ang II-induced hypertrophy in H9c2 cells.
The results summarized in Figure 2 indicate that AOF treatments markedly inhibited the Ang II-induced cytoskeletal change and hypertrophy in H9c2 cells. Therefore, we further examined transcription factor NFAT-3 activation, which is implicated in the development of cardiac hypertrophy. As shown in Figure 3, the nuclear localization of NFAT-3 was markedly induced when cells were treated with Ang II (5 × 10−7 M) for 24 h. In contrast, pretreatment with AOF (60–100 μg/mL) for 1 h before Ang II challenge significantly decreased Ang II-induced nuclear translocation of NFAT-3 (Fig. 3A). AOF also significantly decreased the NFAT3/p-NFAT3 protein ratio levels at 60–100 μg/mL AOF administrations (Fig. 3B).
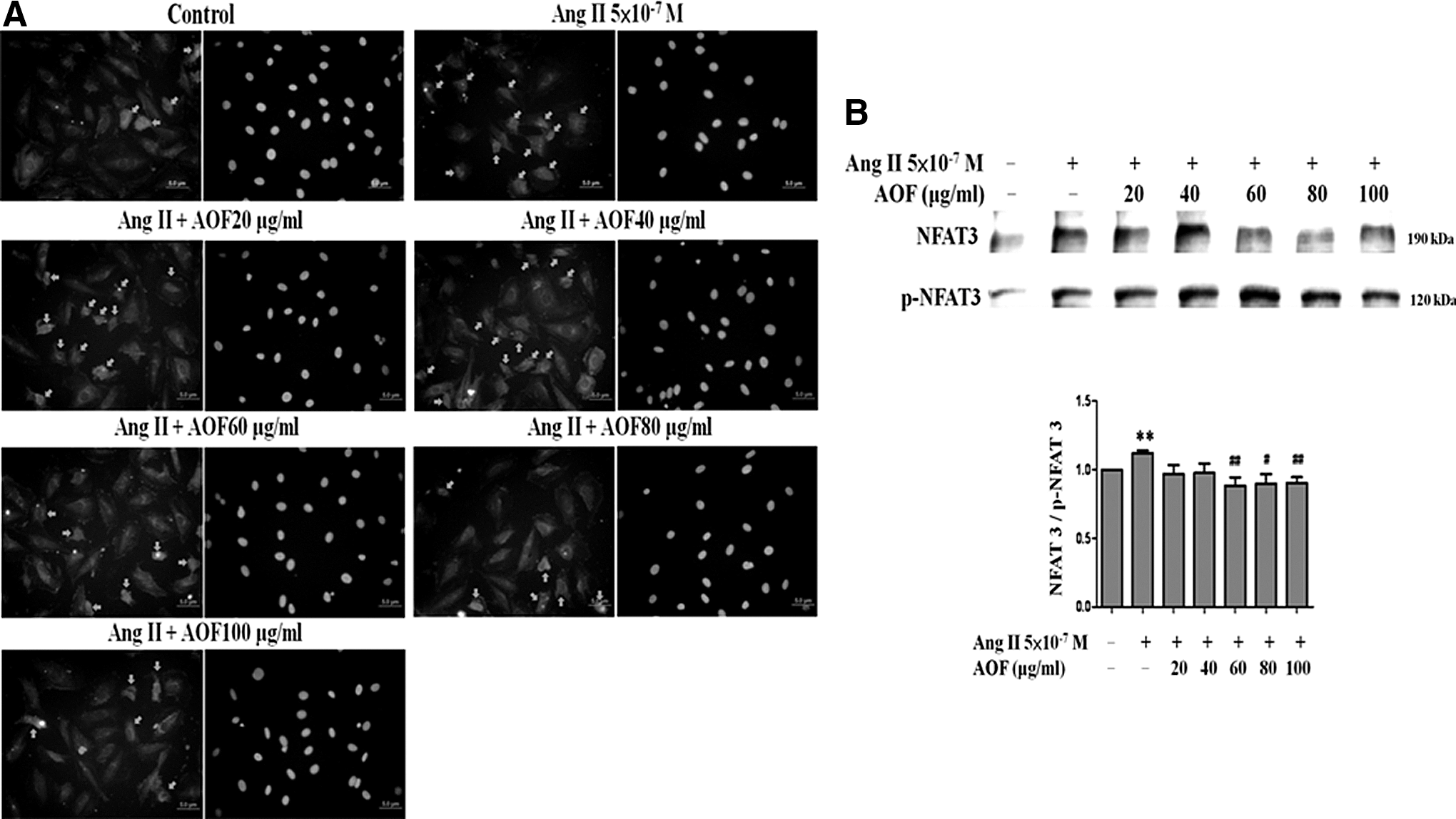
AOF effect on Ang II-induced nuclear localization of NFAT-3 in H9c2 cells.
Gαq, PLCβ3, and calcineurin Ang II-induced IGFII/IIR signaling proteins were measured after AOF pretreatments 1 h before Ang II 24 h challenge to determine whether AOF would inhibit Ang II signaling. As shown in Figure 4, the Gαq, PLCβ3, and calcineurin proteins all increased under Ang II challenge and decreased dose dependently after AOF administration.
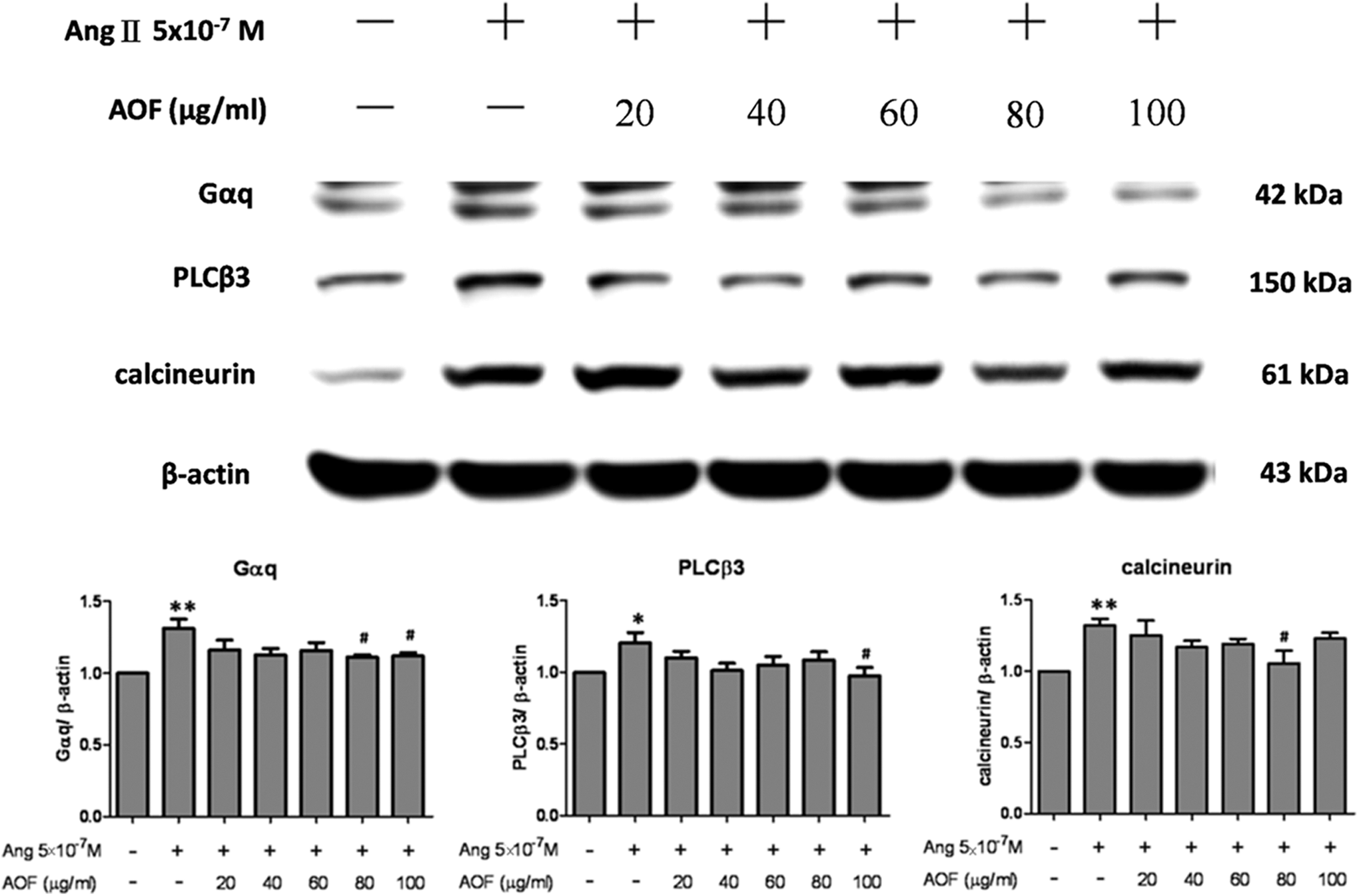
AOF down regulated Gαq, PLCβ3, and calcineurin in Ang II-treated H9c2 cells. H9c2 cells were pretreated with AOF (20, 40, 60, 80, and 100 μg/mL) for 1 h, then treated with Ang II (5 × 10−7 M) for 24 h, and finally harvested and lysed. Entire protein extracts were separated from cells using 12% SDS-PAGE, transferred to PVDF membranes, and immunoblotted with antibodies against Gαq, PLCβ3, and calcineurin. Equal loading was verified with an anti-β-actin antibody. *P < .05 and **P < .01 as compared with the control group, # P < .05 as compared with the Ang II group.
The remodeling proteins were measured to clarify whether AOF would prevent Ang II-induced cardiac remodeling in H9c2 cells. Ang II-treated H9c2 cells exhibited dose-dependent increases in MMP9, MMP2, and tissue plasminogen activator (tPA) levels, which were dose dependently decreased after AOF administration (Fig. 5).
We also examined whether the antihypertrophic effects of AOF are mediated by IGFII. Cells were treated with Leu 27 -IGF-II, an agonist of the IGFII/IIR pathway in mammalian cells. We found that the protein levels of IGFII, calcineurin and ANP were significantly enhanced by Leu 27 -IGF-II (10−8 M); however, the ANP levels, the IGFII and calcineurin were significantly lower after AOF administration (80 μg/mL). Based on those findings, we suggest that AOF elicits a protective effect against Ang II-induced hypertrophy through specifically targeting IGF-II/II R related signaling pathway (Fig. 6).
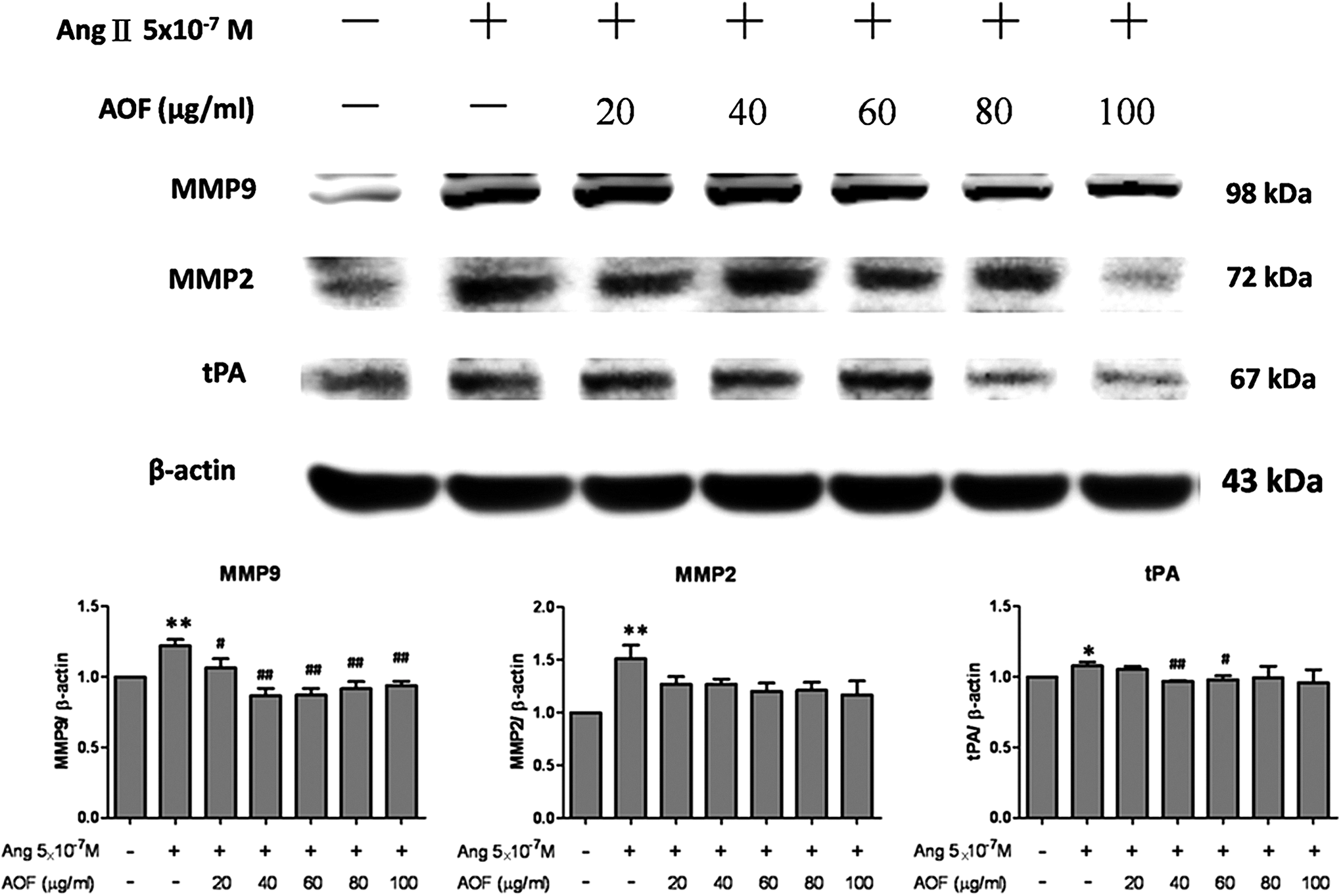
AOF prevented Ang II-induced MMP9, MMP2, and tPA in Ang II-treated H9c2 cells. H9c2 cells were pretreated with AOF (20, 40, 60, 80, and 100 μg/mL) for 1 h, then treated with Ang II (5 × 10−7 M) for 24 h, and finally harvested and lysed. Entire protein extracts were separated from cells using 12% SDS-PAGE, transferred to PVDF membranes, and immunoblotted with antibodies against MMP9, MMP2, and tPA. Equal loads were verified with an anti-β-actin antibody.*P < .05 and **P < .01 as compared with the control group, # P < .05 and ## P < .01 as compared with the Ang II group. tPA, tissue plasminogen activator.

AOF reversed the IGFII, calcineurin, and ANP expressions induced by Leu 27 -IGFII and/or Ang II in H9c2 cardiomyoblast cells. H9c2 cells were pretreated with AOF (80 μg/mL) for 1h, then treated with 10−8 M Leu 27 -IGFII and/or 5×10−7 M Ang II for 24h. Cells were harvested and immunoblotted with antibodies against IGFII, calcineurin and ANP. Equal loads were verified with an anti-β-actin antibody. *P < .05, **P < .01 and ***P < .001 as compared with the control group.
Discussion
We evaluated the protective effects of AOF pretreatment on Ang II-induced H9c2 cardiomyoblast cell pathological hypertrophy in this study. The AOF effect also inhibited the IGFII/IIR-related signaling pathway, which specifically up regulated IGFII as determined by Leu 27 . Leu 27 determined that IGFII is a 7.42 kDa, human IGFII analog that results from a tyrosine 27 -to-leucine 27 mutation. 38 Leu 27 found that IGFII specifically binds to IGFIIR with a high affinity and is able to induce IGFIIR-induced H9c2 cell hypertrophy through Gαq, which involves increased PLC-β3, calcineurin, ANP, and BNP activities. 22,23 AOF pretreatment suppressed the expressions of cardiac fibrosis proteins, metalloproteinase (MMP9 and MMP2), and tPA, induced by the IGFIIR-dependent signaling pathway. 24 Hence, the AOF inhibition might specifically target IGFII/IIR and then down regulate the IGFII/IIR-related signaling pathway that induces cardiomyoblast cell pathological hypertrophy and remodeling.
Numerous studies reported that the renin–angiotensin system (RAS) will increase the angiotensin converting enzyme (ACE) activity when chronic cardiovascular diseases occur, such as hypertension, and further stimulate the formations of LVH and heart failure. 39 –41 RAS system inhibition by ACE inhibitors or AT1 receptor blockers can prevent ventricular remodeling and improve survival rate, 40 suggesting that Ang II is indeed involved in heart failure. 42 Our previous study demonstrated that Ang II seems to evoke IGFII/IIR and further activates cardiac cell apoptosis through calcineurin-dependent pathways. 25 Ang II can also induce cardiac hypertrophy through the calcineurin-dependent transcriptional pathway. 43,44 Our previous study revealed that the IGFII/IIR signaling pathway is involved in cardiac hypertrophy and it occurs through small G-protein-derived signaling pathway regulation. 23
AOF extract has been widely applied as a traditional medicine in China and Korea for the treatment of salivation, polyuria, and neurodegenerative conditions. 31 –35 AOF is also used as a treatment for hypertension and cerebrovascular disorders. 33 Protocatechuic acid (PCA), a phenolic acid that is one of the active components in AOF, is also commonly found in many plant species. 45,46 PCA also showed positive effects against cardio pathogenesis and correlates with the effects of AOF by preventing cardiac apoptotsis and enhancing the cardiac survival in a hypertensive rat model. 47
We noted that AOF concentrations of 60 to 100 μg/mL significantly increased cell viability in this study. The protective effect of AOF against Ang II-induced cytoskeletal changes was associated with modulation in the levels of hypertrophic proteins such as ANP and BNP in H9c2 cells.
Data from our previous studies showed that AOF inhibits Ang II-induced cardiac apoptosis through Ca2+/calcineurin signaling cascades. 37,48 It is well known that calcineurin/NFATc3 is one of the major signaling pathways for downstream calcium. NFATc3 has been reported to play a critical role in Ca2+-mediated cardiac hypertrophy. 49 The dephosphorylation of NFATc3 in the cytoplasm permits translocation into the nucleus and facilitates hypertrophic gene expression. 50 –52 In this study, Ang II caused changes associated with cellular pathological hypertrophy, such as increases in transcription factors NFAT-3/p-NFAT-3, hypertrophic response genes (ANP and BNP), and Gαq down-stream effectors (PLCβ3 and calcineurin) expression. Pretreatment with AOF (60–100 μg/mL) led to a significant reversal of hypertrophic bodies and decrease in hypertrophic proteins ANP and BNP expression levels in H9c2 cells. This means that the antihypertrophic potential of AOF administration inhibits IGFII/IIR downstream signaling induced by Ang II.
Cardiac hypertrophy has been associated with multiple pathophysiological responses such as interstitial fibrosis. 51,53 A previous study indicated that Ang II-induced transforming growth factor-β1 synthesis resulted in the generation of myofibroblast collagen fibrosis. Studies have also suggested that collagen accumulation is coregulated by metalloproteinases and fibroblast growth factor. 14,54,55 Evidence has demonstrated that AOF pretreatment significantly suppresses the cardiac remodeling proteins, metalloproteinases (MMP9 and MMP2), and tPA, which were induced by Ang II challenge.
A similar study showed that the IGFII/IIR signaling pathway plays a critical regulatory role in cell apoptosis in cardiomyoblast cells. IGFIIR is the upstream clacineurin factor, which might contribute to hypertrophy, apoptosis, and heart failure. 22,23,56
We found that hypertrophy inhibition by AOF occurred by targeting the IGFII/IIR-related signaling pathway induced by Leu 27 —IGFII and/or Ang II stimulation in H9c2 cells. That means that AOF can also suppress these upstream IGFIIR signaling factors in addition to the downstream calcineurin signaling. AOF treatment has recently demonstrated good protective effects against oxidative stress, neurotoxicity, and apoptosis in vivo. 31,33,35,37 This study revealed the beneficial effects of AOF in H9c2 cells against Ang II-induced hypertrophy. However, the relative effects of AOF in vivo need to be further clarified.
In conclusion, we provide evidence that AOF protects H9c 2 cells against Ang II-induced pathological hypertrophy by specifically inhibiting the IGFII/IIR-related signaling pathway in H9c2 cell (Fig. 7). The optimum concentrations of AOF are between 60 and 100 μg/mL. Although the PCA effect in IGFII-related signaling pathway needs further clarification, AOF might be a candidate therapy for treating cardiac hypertrophy and for preventing ventricular remodeling prevention in chronic cardiovascular diseases.
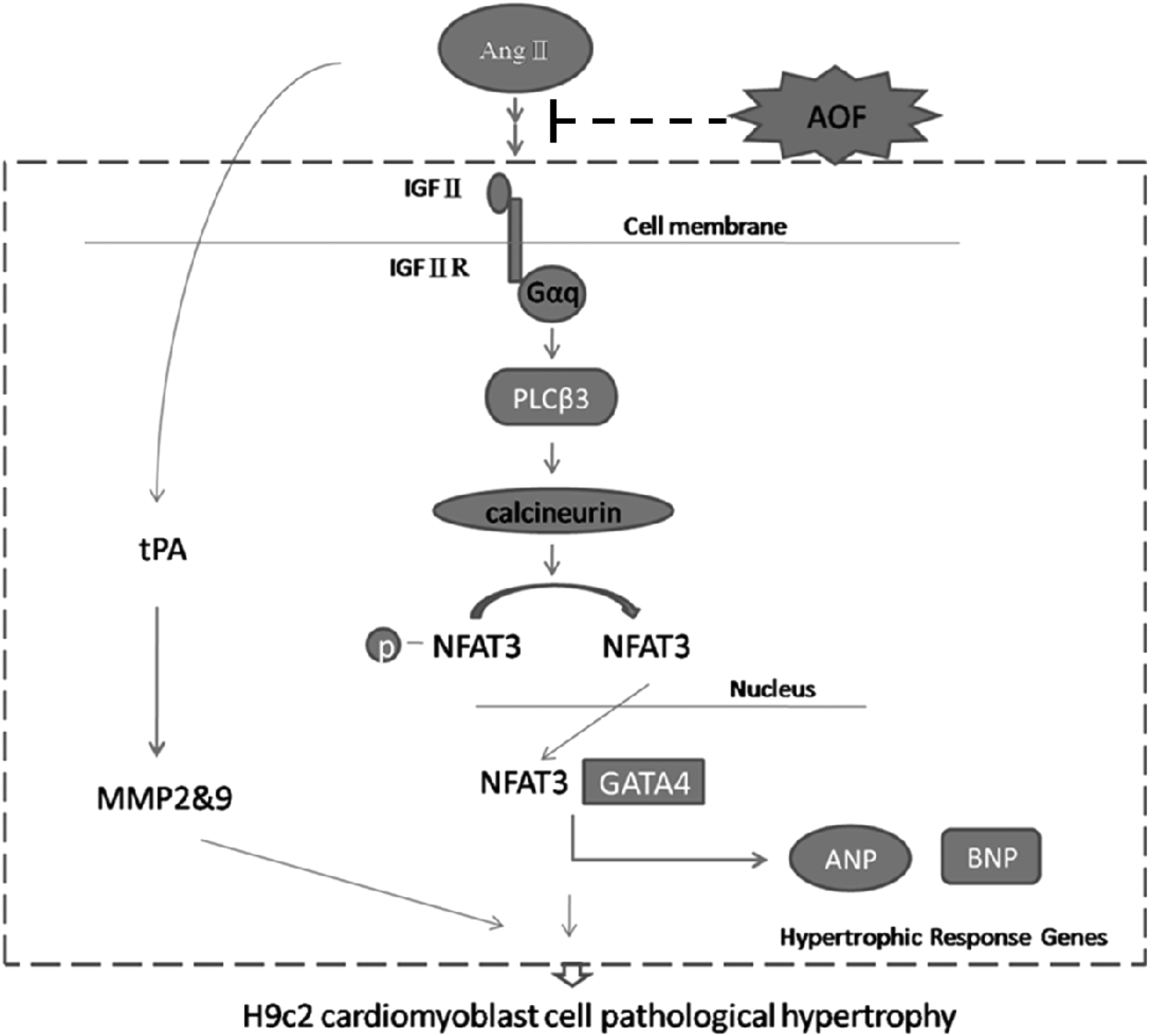
Possible mechanism of AOF protects H9c2 cells against Ang II-induced pathological hypertrophy by specifically inhibiting the IGFII/IIR related signaling pathway.
Footnotes
Acknowledgments
This work was supported by Taiwan Ministry of Health and Welfare Clinical Trial and Research Center of Excellence (MOHW105-TDU-B-212-133019) and, in part, by China Medical University and Asia University (CMU99-COL-08-1, CMU99-COL-08-2). It was also supported by China Medical University under the Aim for Top University Plan of the Ministry of Education, Taiwan.
Author Disclosure Statement
No competing financial interests exist.