
Abstract
Rapid increase in the prevalence of obesity-related metabolic inflammatory diseases has led to research focused on nutraceuticals for their treatment. This study investigated the effects of the ethanol extracts of mung bean testa (MBT) on the metabolic inflammation-induced lipogenesis in gastrocnemius muscle of KK-Ay diabese mice. Ethanol extracts of MBT were orally administered to diabese KK-Ay mice for 4 weeks after diet-induced obesity model was generated by feeding a 60% high-fat diet for 3 weeks. Although there were no changes in body weight gain, MBT treatments decreased total weight of white adipose tissue. MBT also decreased triacylglycerol and total cholesterol levels in the muscle by 30%, which was correlated with suppression of lipogenic genes such as ACC, C/EBP alpha, PGC-1 alpha, and PPAR gamma. In particular, decreased levels of p-ERK1/2, PPAR gamma, and C/EBP alpha in the MBT-treated groups suggest that MBT might inhibit adipogenesis and decrease differentiation via the MEK/ERK pathway. Furthermore, significantly lower amounts of plasma interleukin (IL)-6 and intramuscular tumor necrosis factor (TNF)-alpha and monocyte chemoattractant protein-1 (MCP-1) were detected in MBT groups, confirming the anti-inflammatory effect of mung bean. In addition, our in vitro pilot study with 3T3-L1 cells showed that vitexin, the functional chemical in MBT, inhibited inflammation-induced lipogenesis with significantly lower amounts of IL-6 and MCP-1 after 14 days of vitexin treatment. Thus, the functional compounds in the mung bean ethanol extracts such as vitexin and isovitexin may regulate intracellular lipogenesis and adipogenesis via anti-inflammatory mechanisms and MEK/ERK pathway in the KK-Ay mouse model.
Introduction
A
Mung bean (Vigna radiata L.) is native to Asian regions such as India and South East Asia. Mung bean has been shown to have antioxidant, antitumor, anti-inflammatory, hepatoprotective, antimicrobial, diuretic, and hypoglycaemic effects, in addition to the ability to improve cholesterol levels. 2 –4
Mung bean also contains abundant amounts of flavonoids such as vitexin, isovitexin, and coumestrol. Furthermore, the testa of mung bean (MBT) contains more vitexin, isovitexin, and
Materials and Methods
Sampling extract and high-performance liquid chromatography-mass spectrometry analysis
Mung beans were obtained from local farmers in Keumsung, Korea. MBT with a same quantity were ground using a vibration mill, and put into 80% ethanol solution (5.0 mL) at room temperature for 24 h for extraction. After centrifugation, the supernatant was collected and then used as samples of MBT for testa. For analysis of high-performance liquid chromatography (HPLC) and high-performance liquid chromatography-mass spectrometry (HPLC-MS), the 1 g ground sample of MBT was put into 80% methanol solution for 4 h to prepare the extract. After concentration, the extract was dissolved in 80% methanol (2.0 mL), followed by filtration using a 0.45 μm filter. The conditions of HPLC-MS analysis are described in Figure 1.
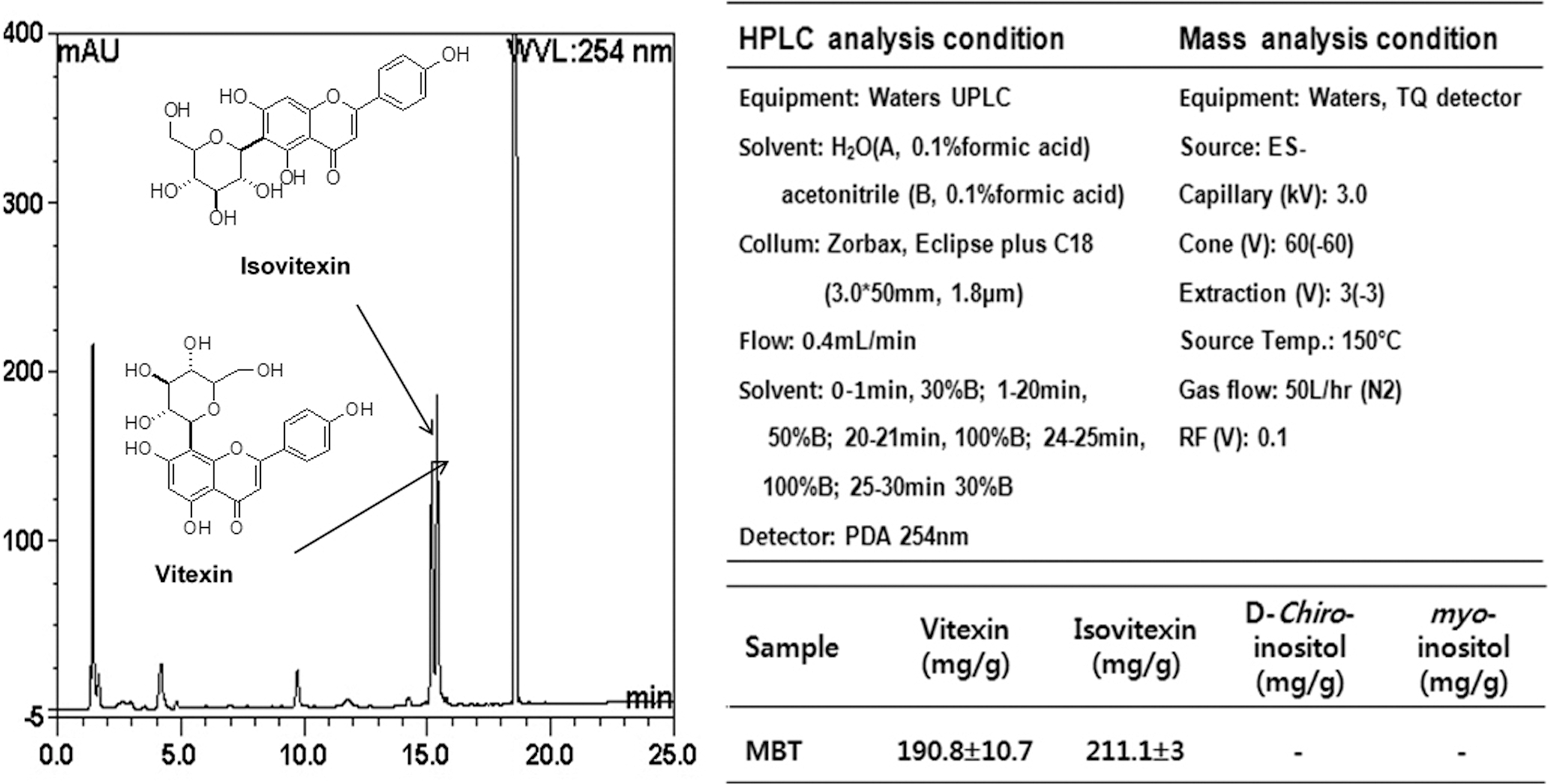
High-performance liquid chromatography (HPLC)-mass analysis and condition to detect effective materials in the ethanol extracts of MBT samples. MBT, mung bean testa; UPLC, ultra performance liquid chromatography.
Experimental design
As a pilot study, we examined whether or not the target chemicals such as vitexin in mung bean ethanol extracts inhibit inflammatory cytokines secretion and lipogenesis using 3T3-L1 preadipocytes. The 3T3-L1 preadipocytes were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). They were cultivated in the preadipocyte media (Dulbecco's modified Eagle's medium [DMEM] with glutamine, glucose, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid [HEPES], and sodium bicarbonarte) with 10% fetal bovine serum (FBS) and 1% antibiotics (penicillin–streptomycin). To induce differentiation, the media including methylisobutylxanthine (0.5 mM), insulin (10 μg/mL), and dexamethasone (1 μM) were replaced. In 2 days, the differentiation media were removed and maintenance media (10% FBS and insulin) were replaced for 8 days. The 3T3-L1 adipocytes, which are suitable for most assays 7–14 days postdifferentiation, were exposed to different levels of vitexin from 10 to 500 μM for every 2 days. Cell proliferation (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide [MTT] at 570 nm), fat accumulation (Oil Red O), and DNA, mRNA and protein expression (Western blotting & reverse transcription polymerase chain reaction) were determined.
For in vivo study, 5 week-old KK-Ay (yellow spontaneous mutation; Clea, Tokyo, Japan) mice, a noninsulin-dependent diabetes model, were used in this study. After 1 week of feeding chow diet, they were fed with high-fat diet (60%) for 3 weeks to induce obesity. Then, the mice were divided into the following three groups: CON, a group fed 60% high-fat diet; MBT, a group fed 60% high-fat diet with MBT(1 g/kg); and Met, a group fed 60% high-fat diet with metformin, which is an oral antidiabetic drug (1 g/kg). All the mice received daily oral injections once a day for 4 weeks. The mice were treated according to the ethical guidelines of the Experimental Animal Holding of Sungshin Women's University (approval number: SSWU AEC 2011-001). After 4 weeks of administration, the mice were sacrificed to collect the plasma and tissue samples for biochemistry and samples were stored at −80°C. We chose gastrocnemius muscle of diabese KK-Ay mice as a target tissue because this type I muscle uses fat from intramuscular fat, liver, or adipocytes as energy sources.
Blood biochemistry
Plasma total cholesterol (TC) and triacylglycerol (TG) levels were measured using a kit (Boehringer Mannheim Biochemical, Basel, Germany) according to the manufacturer's instructions. Plasma insulin and glucose concentrations were determined by mouse enzyme-linked immunosorbent assay (ELISA) kit (ALPCO, Windham, NH, USA). Plasma glycated hemoglobin (HbA1c) levels were measured using a DCA Vantage Analyzer (Siemens, Munich, Germany).
Western blotting
Protein was extracted from gastrocnemius muscle and changes in the phosphorylation status or level of acetyl-CoA carboxylase (ACC), peroxisome proliferator-activated receptor gamma (PPARγ), CCAAT/enhancer binding protein alpha (C/EBPα), phosphorlyated 5′ adenosine monophosphate-activated protein kinase (p-AMPK), AMPK, proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α), GLUT4, p-p38, p38, phosphorlyated extracellular signal-regulated protein kinases 1/2 (p-ERK1/2), ERK1/2 were determined by western blotting.
Tissues were homogenized in lysis buffer supplemented with Halt™ Protease and phosphates Inhibitor Single-Use Cocktail (Pierce, Rockford, IL, USA). The supernatants were boiled at 95°C for 5 min in 2×Laemmli sample buffer (Bio-Rad, Hercules, CA, USA). The total protein concentrations of whole tissue extracts were determined by Bradford assay. Protein samples were separated by 8–10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto nitrocellulose membranes (Bio-Rad, Hercules, CA, USA). The membranes were blocked in 5% skim milk or bovine serum albumin (BSA) in Tris-buffered saline with 0.1% Tween 20 (TBS-T) and probed with appropriate primary antibodies (all from Cell Signaling, Danvers, MA, USA) for 3 h. Secondary antibodies were conjugated to horseradish peroxidase (Enzo Life Sciences, Farmingdale, NY, USA) and the signals were developed by chemiluminescence (BioFX, Owings Mills, MD, USA). The signals were visualized by exposing the membranes to X-ray films (Agfa, Mortsel, Belgium), and quantified using an Image J Analyzer (NIH, Bethesda, MD, USA).
Proinflammatory cytokine assay
The tissue concentrations of TNF-α, IL-6, and monocyte chemoattractant protein-1 (MCP-1) were determined with an ELISA kit (BioLegend, San Diego, CA, USA). The change in color was measured at a wavelength of 450 nm using a microplate reader (Multiskan Spectrum; Thermo Electron Co., Vantaa, Finland). Cytokine levels were expressed as pg/mg total protein. The protein content of the samples was measured by the method of Bradford; BSA was used as a standard. 8
Statistical analysis
Statistical analyses were performed using IBM SPSS software for Windows version 21. All data were presented as a mean±standard deviation. Data were analyzed by one-way analysis of variance with Duncan's test, at a significance level of P<.05.
Results
Functional materials in MBT
Vitexin and isovitexin concentrations were assayed to be 190.8 and 211.1 mg/g in MBT (Fig. 1). However, DCI and myo-inositol, which are mainly present in MBT, were barely detectable and coumestrol, which has been reported to increase during mung bean germination, was not detected at all in MBT.
Vitexin on inflammation
Administration of vitexin at doses of 10, 100, and 500 μM dose dependently decreased fat accumulation in 3T3-L1 adipocytes by 20%, 45%, and 80%, respectively, compared to the fat accumulation in control cells (data not shown). Likewise, the lipogenesis biomarkers (adipocyte protein 2 [aP2] mRNA) and lipolysis marker (phosphorylation of AMPK) were changed by 100 μM vitexin treatment in the same manner with decreases in fat accumulation (Fig. 2A, B). When cells were treated by vitexin for 14 days during adipocytes differentiation, the changes in inflammatory cytokines such as IL-6 and MCP-1 from day 14 to 0 were significantly lowered by vitexin levels despite a lack of changes in TNF-α (Fig. 2C). Therefore, we concluded vitexin, a major flavonoid of MBT ethanol extracts, might decrease levels of IL-6 or MCP-1-induced lipogenesis as doses increase.
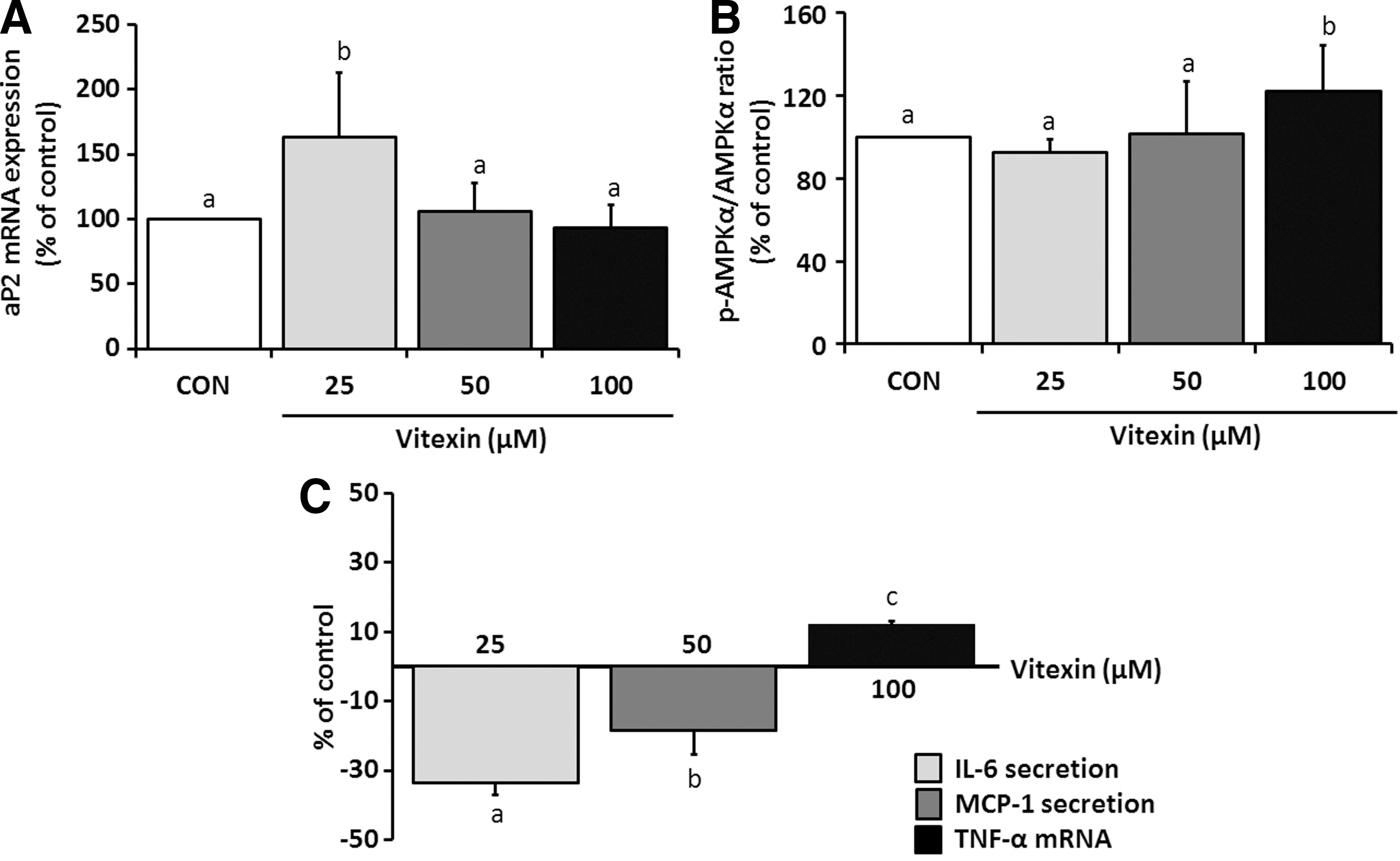
Inhibitory effects of vitexin on inflammation-induced lipogenesis in 3T3-L1 cells. Cell proliferation and fat accumulation were significantly lowered as the dose of vitexin increased.
Changes in body composition
Hyperglycemia and urine glucose significantly increase in KK-Ay mice if their body weight exceeds 35 g. In this study, all mice groups had a mean body weight of >50 g, 2 weeks after starting a high-fat diet. The body weight gain was found to be higher in the MBT and Met groups than in the CON group, but it was not statistically significant (Table 1). No difference in the food intake or food efficiency (body weight gain/feed intake amount) was found among the groups, explaining that MBT did not promote weight loss or changes in the amount of food consumed (data not shown). In addition, there was no difference in blood glucose level between CON and MBT groups, and that from Met group was significantly lower as metformin is an antidiabetic drug that suppresses glucose production and reduces fasting glucose levels. 9 However, the total white adipose tissue including epididymal and perirenal adipose tissues significantly decreased in the MBT groups compared with the CON or Met groups, and the decrease in epididymal adipose tissue was more significant than the decrease in perirenal adipose tissue (Fig. 3).
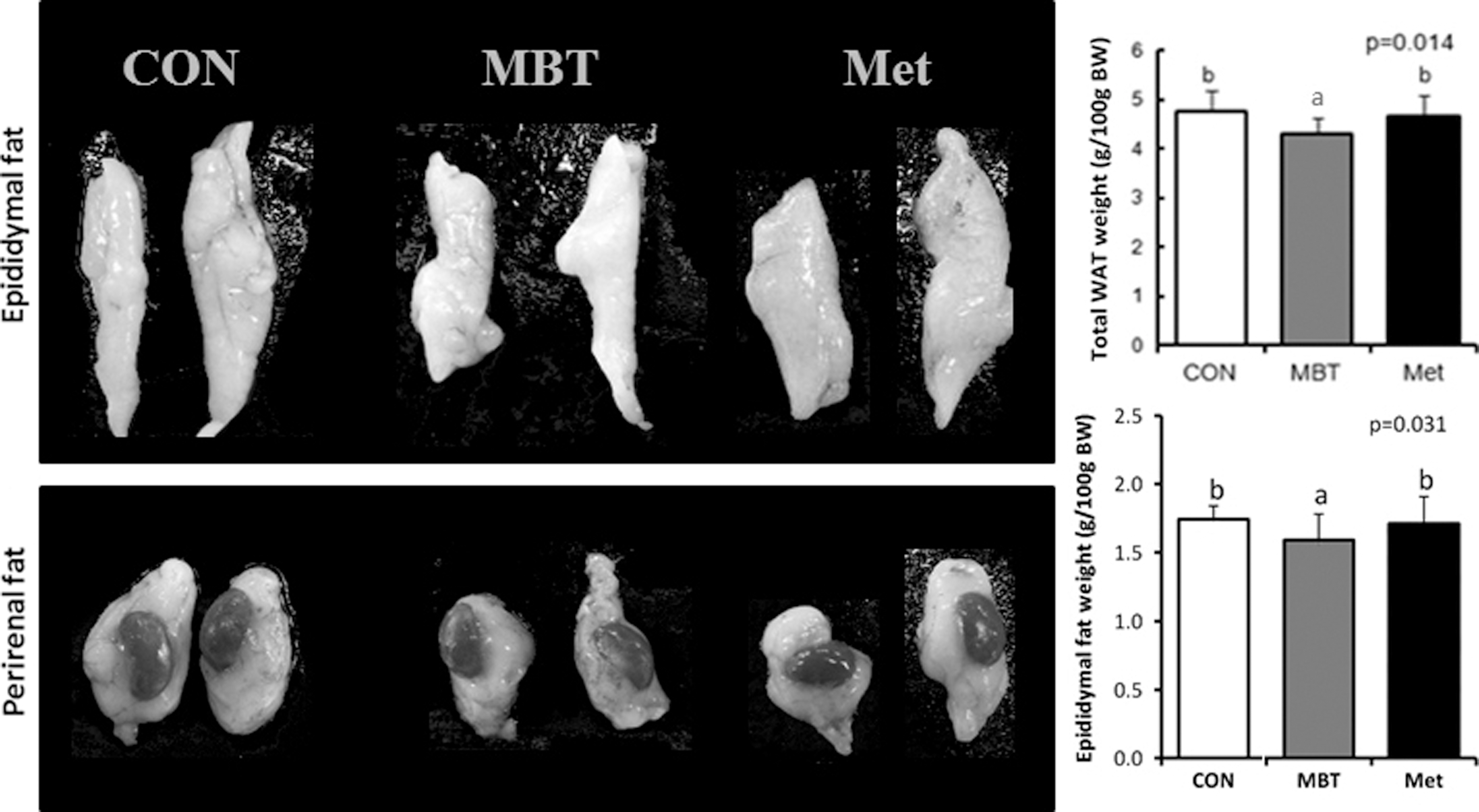
Effects of MBT on white adipose tissue (WAT) weights in KK-Ay mice fed with 60% HFD. CON: 60% HFD; MBT: 60% HFD+MBT; Met: 60% HFD+Met. The weights of total WAT and epididymal and perirenal fat were significantly lower in MBT group when compared with CON group. Data on the graph represent the mean±SD of mice for each group. Different letters indicate significant differences among groups. Data were analyzed by one-way analysis of variance (ANOVA) with Duncan's test. CON, control; HFD; Met, metformin.
Data were presented as the mean±SD (n=5) of mice for each group. Different superscript letters indicate different groups with significant statistical differences. Data were analyzed by one-way analysis of variance with Duncan's test.
BWG, body weight gain; CON, control; MBT, mung bean testa; Met, metformin; N.S., not significant; TC, total cholesterol; TG, triacylglycerol.
Blood lipid profiles and insulin resistance
Even though the plasma TG and TC levels increased in the MBT groups compared to CON group, the levels were lower than those in the Met group (P<.01). No significant difference in HbA1c levels were found among the groups (data not shown). In this study, it was the reason why the fasting blood glucose level exceeded 300 mg/dL at 3 weeks after being fed a high-fat diet, which also demonstrates the characteristic feature of KK-Ay mice. We found that MBT treatments could not decrease the fasting blood glucose levels in mice fed a normal chow diet compared with the high fat diet (data not shown). The pancreatic tissue section was stained with an insulin antibody. The immunohistochemical analysis of pancreatic tissue indicated that the insulin level significantly decreased in the Met group but not in the MBT group (data not shown).
Inflammatory adipocytokines
Plasma TNF-α levels tended to increase by 21% in the MBT group and 43% in the Met group, compared with the CON group, but these levels were not statistically significant. Plasma IL-6 levels significantly decreased by 96% in the MBT group and 19% in the Met group, compared with the CON group (P<.01). No significant difference in the MCP-1 level was observed among the groups (Fig. 4A). In the gastrocnemius muscle, the TNF-α level significantly decreased by 34% in the MBT group, and 13% in the Met group, compared with the CON group. The MCP-1 level significantly decreased by 41% in the MBT group, and 14% in the Met group, compared with the CON group. No significant difference in the IL-6 level was found among the groups (Fig. 4B). In the liver, no difference in the levels of the three cytokines was found among the groups (data not shown). In summary, mice in the MBT groups showed significantly decreased plasma IL-6 levels, and intramuscular TNF-α & MCP-1 levels.

Effects of MBT on proinflammatory cytokines from KK-Ay mice fed with 60% HFD.
Inflammation-induced adipogenesis and oxidative stress
Intramuscular TG and TC levels significantly decreased by ∼30% in the MBT compared with the CON group, similar to the Met group (Fig. 5A). No differences in the hepatic lipid levels were observed among the groups. In addition, no differences in the levels of blood aspartate and alanine aminotransferases (AST and ALT), which are markers of liver injury, were found after MBT administration (data not shown). With regard to the expression of proteins related to inflammation-induced lipogenesis such as the ACC, PPARγ, and C/EBPα protein, expression levels were significantly lower in the MBT groups (Fig. 5B, C). The expression of ACC protein was significantly lower by 51% and 39% in the MBT and Met groups, respectively, compared to the CON group (P=.037). The expressions of PPARγ and C/EBPα protein were also significantly decreased in treatment groups compared with the CON group (P<.001). Despite increases in the expression of AMPK protein in the MBT groups, the p-AMPK/AMPK ratio decreased. The expression of PGC-1α protein significantly decreased by 27% and 13% in the MBT and Met groups, respectively, compared with the CON group (P=.009). The expression of GLUT4 protein increased in the MBT and Met groups, but not significantly (Fig. 5B). The levels of p-ERK and p-p38, which increase under conditions of oxidative stress, decreased in the MBT groups (Fig. 5C).
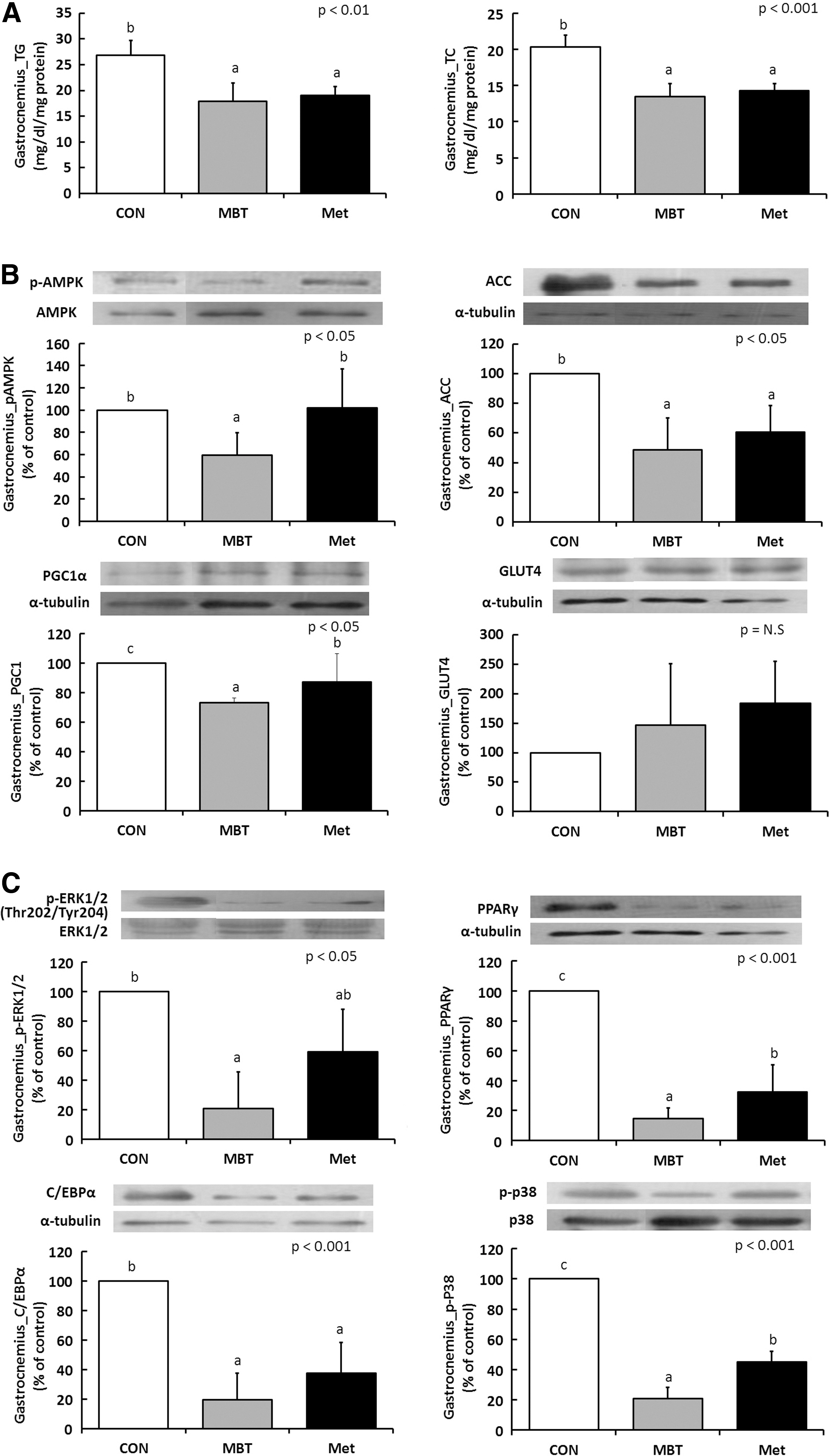
Effects of MBT on lipid accumulation, lipogenesis, and adipogenesis in KK-Ay mice fed with 60% HFD.
Discussion
It is known that mung bean has protective effects such as antioxidative and anticancer activities, but only a few studies have been conducted on its effects on obesity. 10 Our results in KK-Ay mice showed no differences in the final body weight after MBT administration and these results corresponded with those of Yao et al. 3 They reported that no changes were observed in the body weight of mice after administration of either 2 or 3 g/kg of mung bean sprouts or 3 g/kg seed coat of mung bean, a dose which was three-fold higher than the MBT used in this study. In this study, the epididymal fat weight significantly decreased after MBT administration, suggesting that mung bean inhibited the increase in body fat caused by a high-fat diet. We found that vitexin in MBT extracts reduced fat accumulation in a pilot study with 3T3-L1 cells; the effect of mung bean extracts on the epididymal fat reduction may come from vitexin. In particular, no lipid accumulation was found in the mouse livers from the MBT group, and the intramuscular TG and TC levels significantly decreased by 30%. It is likely that intramuscular fat was used as an energy source and lipid precursors such as acetyl CoA, which were excessively produced during this process, were produced in the liver and released into the blood without accumulating in the liver.
We concluded that MBT treatments significantly decreased the expression of lipogenic genes such as ACC, PPARγ, and C/EBPα. The ACC gene, which is a rate-limiting enzyme of fatty acid synthesis; C/EBPα, which is a liver-enriched transcriptional regulator for fat metabolism; and PPARγ, which promotes adipocyte differentiation and fatty acid storage, were decreased by the MBT treatment. 11 This is likely to be associated with the decreased TG and TC levels in the muscle and the decreased amounts of epididymal fat. This result appears to be associated with the possible mechanisms of intracellular homeostasis such as AMPK-induced lipolysis, inflammation-induced lipogenesis, MAPK-induced insulin resistance, and so on.
The first possibility is that MBT may activate AMPK-induced lipolysis. AMPK is a critical regulator of leptin-induced fatty acid metabolism and glucose uptake in skeletal muscle. 12 –14 Even though vitexin in mung bean extracts increased AMPK protein expression and phosphorylation of AMPK in 3T3-L1 cells, p-AMPK protein decreased in the MBT groups despite the increase in AMPK in vivo. Therefore, p-AMPK level was not high enough to inhibit energy-consuming pathways while metformin activated p-AMPK-induced ACC activation. Among the novel mechanisms of p-AMPK signaling, ACC suppression and PGC-1α and GLUT4 expression were not corresponded to AMPK activation in the gastrocnemius muscle tissue with MBT treatment. Therefore, MBT-induced lipogenesis inhibition was not triggered by AMPK activation.
The second possibility is that MBT may decrease inflammation-induced lipogenesis. Macrophages associated with adipose tissue have been implicated in chronic, low-grade inflammation of adipose tissue in obesity, which is causally linked to insulin resistance. 13 In particular, TNF-α secreted by cells plays an important role in inflammation, and IL-6 has proinflammatory and anti-inflammatory effects, which is secreted from T cells and macrophages. 15 MCP-1 is an inflammatory chemokine and it affects obesity, and insulin resistance. We found inflammatory cytokines such as IL-6 and MCP-1, but not TNF-α, from day 14 to 0 were significantly reduced after vitexin treatment for 14 days in adipocytes. In an in vivo study, plasma IL-6 level and intramuscular TNF-α and MCP-1 levels significantly decreased in the MBT group compared with high-fat diet or metformin groups. Thus, it is likely that the MBT used in this study had a positive effect on the improvement of insulin resistance by reducing the expression of TNF-α and MCP-1, which are inflammatory markers in diabetes and obesity.
The third hypothesis is that MBT may inhibit adipogenesis and decrease differentiation via MEK/ERK pathway. Our result showed a decreased amount of p-ERK1/2, PPARγ, and C/EBPα after MBT administration. Studies demonstrated that activation of the MEK/ERK signaling pathway during the first 12 h of adipogenesis improved the activity of factors that regulate both C/EBPα and PPARγ expression. 16,17 On the other hand, when MEK/ERK expression increased during the late stage of adipogenesis, adipogenic gene expression decreased due to MAPK-dependent phosphorylation of PPARγ. As PPARγ serves as the adipocyte-specific transcription factor, it is also used as a substrate of ERK. Its transcriptional activity becomes diminished and adipocyte differentiation is inhibited by this phosphorylation. Therefore, the function of ERK in adipogenesis has to be regulated opportunely: at the beginning, ERK is turned on for a proliferative step and has to be shut-off later to avoid PPARγ phosphorylation. 18 As seen in our result, the expressions of ERK, PPARγ, and C/EBPα were all decreased, suggesting that ERK expression might be blocked during the early stage of adipogenesis after MBT administration. MBT appears to work through the MEK/ERK pathway during adipogenesis so that it inhibits adipogenic gene expression and decreases lipid accumulation in the gastrocnemius muscle.
As the last hypothesis, MBT may decrease MAPK-induced insulin resistance and as a consequence inhibit lipid accumulation. Type 2 diabetes mellitus may also result from insulin resistance via interactions with insulin-target organs such as the liver, fat, and muscle. As branched chain amino acids (leucine and valine), which are abundant in mung bean and are used as energy sources during muscle contraction, they not only inhibit muscle damage but also relieve insulin resistance. 19 The KK-Ay mouse model used in this research progressively develops glucose intolerance, hyperglycemia, hyperinsulinemia, and obesity by the age of eight weeks. 20 In this study, the PPARγ and PGC-1α levels of the muscle tissue significantly decreased in the MBT groups, with no significant difference in GLUT4. PPARγ has been known to regulate not only adipocyte differentiation but also glucose metabolism; therefore, it has been regarded as a target for antidiabetic drugs. It was reported that systemic insulin resistance occurred in PPARγ knockout muscle, but that the glucose utilization rate was found to be normal in the muscle. 15 However, another study reported that gradual and serious insulin resistance occurred due to insulin-induced glucose utilization in the muscle. Thus, the physiological importance of PPARγ in the muscle needs further investigation. 21 PGC-1α is a transcription factor that plays a critical role in fatty acid oxidation by activating GLUT4, which serves as a mechanism for glucose absorption and regeneration of mitochondria in the muscle. 22 During physical exercise, the increased PPARγ and PGC-1α expression in the muscle may mediate the beneficial effects of insulin sensitivity. 11 Thus, it is clear that PGC-1α and PPARγ expression affect energy utilization in the muscle.
Other studies on the effect of mung bean on blood glycemic control have shown that glucose levels decreased by 21% and 129% when DCI (contained in mung bean) was administered to streptozotocin-induced diabetic rats at a single dose of 15 and 100 mg/kg, respectively. 23 However, no DCI was detected in the MBT used in this study, which might have resulted from using a different variety of mung bean from different regions such as China and Korea. In addition, among the MAPKs activated by various oxidative stresses, the expression of p-ERK and p-38 decreased in the MBT groups, compared with the CON group. As these MAPKs inhibit intramuscular metabolic stress that may occur during intensive exercise, they are likely to be correlated with insulin resistance and reduction in the expression of adipogenic proteins. Taken together, the hypothesis seems to be supported; however, the GLUT4 result of a nonsignificant increase makes it inconclusive.
As described earlier, mung bean is rich in branched chain amino acids such as leucine (Leu), isoleucine, and valine, which are known to improve insulin resistance and increase insulin sensitivity. 24,26 However, it was reported that 100 or 200 μM of Leu induced the downregulation of AMPK by 50% and 70% and caused insulin resistance in vivo. 26 In this study, the level of Leu in MBT extracts may be higher than those 26 considering the solubility of Leu in ethanol solutions. 27 Decreased AMPK activity has been detected in tissues of many obese insulin-resistant rodents and in liver and adipose tissue of rats. 28 –31 This may be the reason why phosphorylation of AMPK decreased despite increases in AMPK in vivo, with opposite results from our in vitro study. As mung bean is not composed of a single nutrient, it appears that both the beneficial effect from vitexin and adverse effect from Leu on AMPK activity and insulin resistance occurred at the same time. Therefore, in terms of developing functional foods in industries, we need to consider these inconsistent effects from Leu to utilize mung bean as antiobesity nutraceuticals.
Vitexin and isovitexin that are abundant in mung bean were shown to be involved in the regulation of inflammation-induced lipogenesis, oxidative stress, insulin resistance, and lipid synthesis. We found vitexin and MBT ethanol extracts significantly decreased the inflammation-induced lipogenesis and adipogenesis with in vitro and in vivo study. Therefore, these compounds could be useful for preventing metabolic inflammatory diseases. Further study is required to investigate the exact mechanisms by which mung bean compounds exert these beneficial effects, including experiments in transgenic adipocytes.
Footnotes
Acknowledgments
This work was supported by two grants, the Cooperative Research Program for Agriculture, Science, and Technology Development (PJ907089) and the Technology Commercialization Support Program, Ministry for Food, Agriculture, Forestry, and Fisheries (811003031SU000), Republic of Korea.
Author Disclosure Statment
No competing financial interests exist.